祁连山中部不同树龄及不同生长率青海云杉径向生长对气候的响应*
2020-01-09詹思敏冉依林张凌楠刘晓宏
李 卉 詹思敏 冉依林 张凌楠 刘晓宏
(陕西师范大学地理科学与旅游学院,陕西 西安 710119)
0 引 言
树木作为重要的自然资源,它的生长受自身遗传因素和气候因素的影响[1].树木自身的遗传因素可能会影响到树木寿命的长短和生长的速度,进而影响树木对气候要素的响应[2].随着年龄的增加,树木茎干粗度和高度也随之增加,从而导致水分运输距离增加[3];另外,树木根系也会不断生长,从土壤中的吸收养分和水分的能力增强.二者都可能导致树木生长对气候的响应随年龄的变化而改变[4-5].树木初始生长率的快慢差异也会影响树木对于气候因子的响应.同时有研究显示初始生长率较低的树木比初始生长率高的树木拥有更长的寿命,而初始生长率快的树木可生存的时间长短则不尽相同[6-7].不同树龄与不同初始生长率的树木对气候因子的响应略有差异[8].
关于不同树龄树木对气候因子响应的研究主要分为两种观点:一种是老龄树比幼龄树对气候响应更敏感[9-10].例如,对于意大利阿尔卑斯山高欧洲落叶松(Larixdecidua)和瑞士石松(Pinuscembra)的研究表明,2个树种对气候要素响应一致,且响应程度随年龄增加而加强[11];同样,奥地利阿尔卑斯山的挪威云杉(Piceaabies)对5—6月的降水的响应随着年龄的增加而增强[10].另一种观点则为幼龄树比老龄树对气候响应更敏感[12-13].例如,对于青藏高原东南部色季拉山长苞冷杉(Abiesgeorgeivarsmithii)的研究显示幼龄树比老龄树对气候的响应更显著[14];在西天山的研究显示树龄小于210岁的雪岭云杉(Piceaschrenkiana)对于气候更加敏感[15];对于葡萄牙西北部的海岸松(Pinuspinaster)的研究也显示幼龄树的早材宽度比老龄树对气候响应更敏感,而老龄树的晚材宽度则对气候更敏感[16].老龄树对气候因子的敏感性低,可能是由于生长率的下降使得树木生长所需的资源减少;而幼龄树的径向生长对气候更敏感,可能是由于幼龄树根系浅,吸收不到更多的水分与养分,因此降水少或高温导致的蒸发加剧使幼树获取的水分变化更剧烈,生长更易受到气候的影响[17].研究不同树龄树木对于气候因子响应差异,对于分析祁连山森林树龄结构、预测森林发展动态有着重要意义[18].现有关于不同初始生长率树木对气候因子的响应的研究,大多是通过分析初始生长率对于树龄的影响,从而探究不同初始生长率树木与气候因子的相关关系,而直接比较不同初始生长率树木对气候因子响应差异的研究还比较少.例如,在崆峒山的研究显示生长缓慢的树木对生长季前期的降水更为敏感[19].同样,在树木年轮学中,通常利用初始生长率较低、生长缓慢的老龄树来进行气候重建,这也从侧面反映了初始生长缓慢的树木对于气候有着更强的响应[20].
祁连山是中国西部重要的水源涵养区,是维护青藏高原生态平衡、维持河西走廊绿洲稳定、保障西部地区生态安全的天然屏障,是我国生物多样性保护的优先区域,也是西北地区重要的生物种质资源库.虽然在祁连山已经进行了很多树轮气候学的研究,但关注不同树龄及初始生长率青海云杉对气候响应的研究还很有限.本研究以祁连山隆畅河林场的青海云杉(Piceacrassifolia)作为研究对象,探究不同树龄及不同初始生长率树木径向生长对于气候因子的响应,主要探究:树木年龄与初始生长率间的关系;初始生长率相近、不同树龄的树木对气候因子响应的差异;树龄一致时不同初始生长率的树木对气候因子响应的差异.
1 研究区概况
祁连山作为我国西北部重要山脉之一,东起乌鞘岭,西至阿尔金山东段,地势西北高、东南低;其东部受东亚季风影响,西部受西风带影响,两风带在祁连山中部交汇,降水由东南向西北逐渐减少,温度也随之逐渐降低.祁连山海拔高度为2 500~4 500 m,植被表现出明显的垂直地带性,祁连山的优势针叶树种青海云杉和祁连圆柏主要分布在海拔2 650~3 200 m[13].青海云杉具有较高的气候敏感性,适合用于研究祁连山树木径向生长对气候因子的响应.
2 样品采集处理和气象资料收集
2.1 样品采集和处理
2016年8月于祁连山中部的隆畅河林场采样,选取无人类活动干扰或其他自然灾害影响、树木竞争较小,经纬度一致(38°46′48″N,99°39′36″E),海拔不同的3个采样地进行取样,采样点分别为LCH1、LCH2、LCH3,相关详细信息见表1. 选定采样点树木后,在树木距离地面1.3 m处使用的生长锥(直径5.15 mm)沿平行坡面和垂直坡面方向各钻取1个树芯并按照标准的树木年轮处理方法对样芯进行处理[21]:样芯固定、阴干、打磨,并根据年轮的宽窄进行初步定年[22].利用Lintab 6树木年轮宽度仪测量树轮宽度[23].
2.2 气象资料收集
研究过程中采用的气象数据来源于距离采样点较近的野牛沟气象站和肃南气象站,其具体信息见表2.通过计算2个气象站1960—2013年的月平均温度、月总降水量发现降水集中在7和8月,温度较高的月份为6—8月(图1).由于气象站和采样点海拔不一致,因此采用反距离权重方法计算采样点的月总降水量[24],并根据气温直减率计算样点处的月平均温度[25].选用树轮形成前一年6月至当年10月的月总降水和月平均温度作为气候因子,分析树木径向生长与气候因子的相关关系.

表1 采样点信息

表2 祁连山采样点周围气象站信息
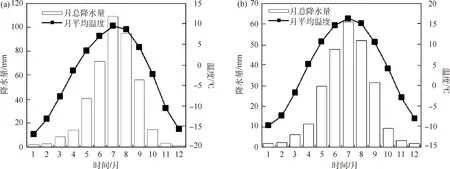
图1 2个气象站逐月平均气温和月总降水量(1960—2013年)
3 分析方法
3.1 年表建立
使用Cofecha软件对初步定年的每个采样点定年的结果和测量得到的轮宽序列进行检验,再用Arstan软件中步长50年的样条函数对原始树轮宽度序列进行去趋势[26],最终得到各采样点的总年表及每根样芯的标准年表[21].
3.2 数据分组和相关分析
虽然3个采样点海拔略有差异,但通过对3个海拔采样点分别建立年表,并分析3个总年表与气候因子的相关关系(1960—2013年),发现其无显著差异(图2),故将3个采样点数据进行合并.在确定树木年轮时只保留树心一侧最后1个年轮距离树心距离<2.0 cm的样芯,并使用直尺结合目测的方法确定髓心,用年轮的数量及估计的缺失树轮数量计算树木的年龄[27-28].本研究地青海云杉生长良好,年轮宽度较宽,且树木在幼年时生长较快,因此距离髓心2.0 cm处缺失的年轮数量较为有限,不会过于影响对树龄的估计.由于胸径处的树木年龄代表每棵树生命的早期阶段,因此通过平均每棵树前40年的原始年轮宽度(包括缺失年轮的数量)来计算初始生长率(单位: mm/y).为分析树龄与初始生长率的关系,使用聚类分析将样本划分为3组,Ⅰ组为树龄大但初始生长率很低的样本,Ⅱ组为树龄小而初始生长率高的样本,Ⅲ组则是树龄与初始生长率均很低的样本.根据对所有单根样芯年表聚类结果可看出,Ⅰ和Ⅲ组的初始生长率相近(初始生长率<1.8 mm/y),可合并用来反映树龄对于径向生长与气候因子相关关系的影响,合并为年龄组;Ⅱ组和Ⅲ组的树龄相近(树龄<125岁),可合并用来反映初始生长率对径向生长与气候因子相关关系的影响,合并为生长率组(图3).本研究共采集133根树芯作为样本,分组后,年龄组有样芯65根,生长率组有样芯95根.计算单根年表与气候因子在1960—2013年的Pearson相关系数.用线性拟合分析树龄及初始生长率与径向生长和气候要素相关系数绝对值的关系.
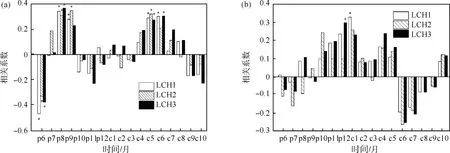
图2 3个采样地总年表与气候因子的相关关系(1960—2013年)
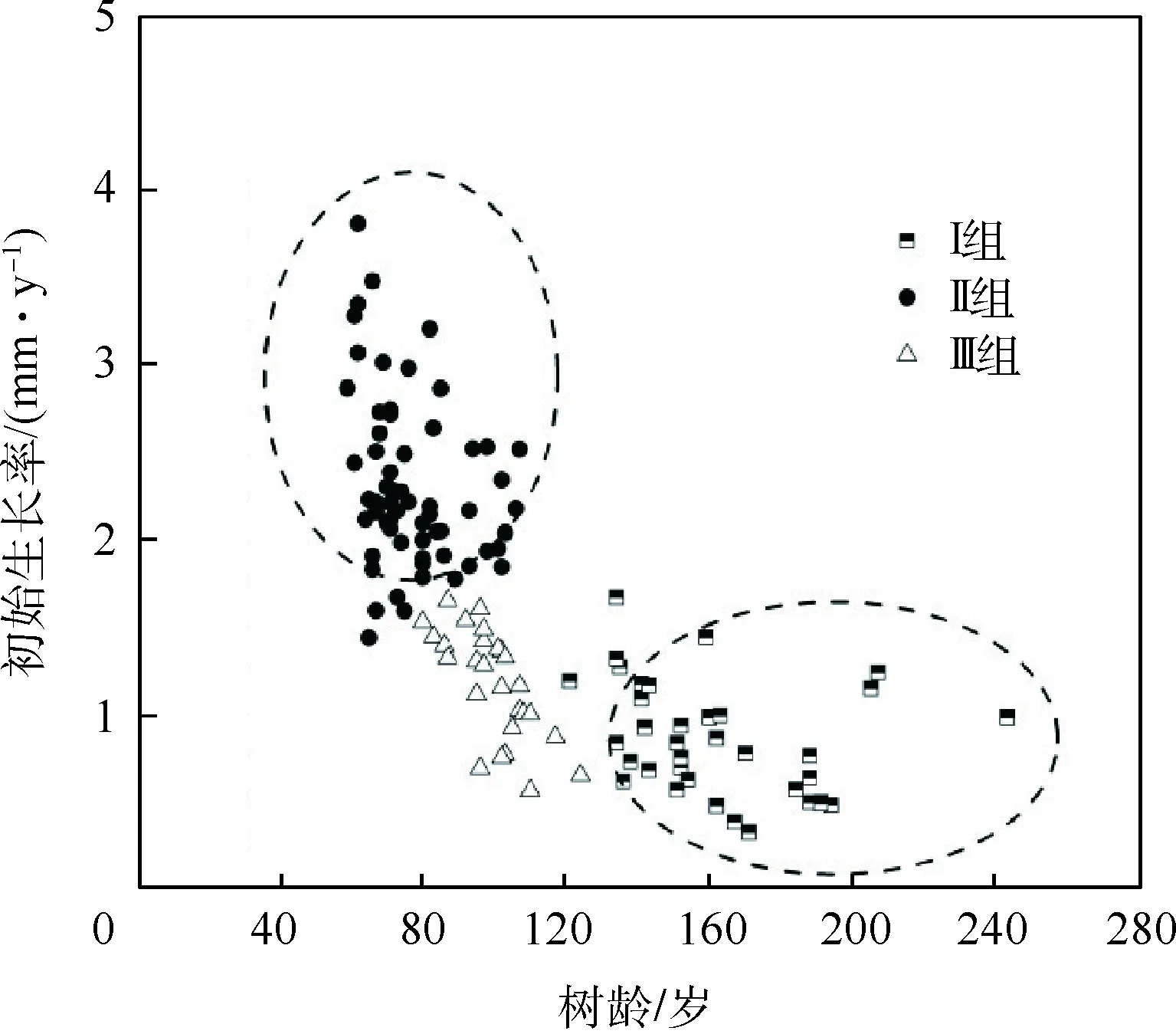
图3 树龄与树木初始生长率关系
4 结 果
4.1 样地年表及单根年表与气候因子的相关关系分析
通过对树木径向生长与不同月份气候因子的相关分析可以看出,年龄组的树木径向生长与前一年8月降水(Pp8)、当年6月降水(Pc6)以及前一年8月至当年6月的累积降水(Pp8c6)存在着显著的正相关关系;其中约50%的样芯年表与Pp8显著正相关关系,70%的样芯年表与Pc6、Pp8c6显著正相关关系[图4(a)].同时年龄组的树木与前一年6月的月平均温度(Tp6)、前一年7月的月平均温度(Tp7)、当年6月的月平均温度(Tc6)和当年7月的月平均温度(Tc7)呈现出显著负相关关系:表现为约45%的样芯年表与Tp6显著负相关关系,约65%的样芯年表与Tp7显著负相关关系,75%的样芯年表与Tc6、Tc7显著负相关关系[图4(b)].生长率组的树木也表现为与Pp8、Pc6以及Pp8c6存在正相关关系,其中约50%的样芯年表与Pp8显著正相关关系,约70%的样芯年表与Pc6和Pp8c6显著正相关关系[图4(c)].此外,生长率组70%的样芯年表与Tp7、Tc6和Tc7表现出显著负相关关系[图4(d)].
4.2 径向生长对气候响应与树龄及初始生长率的关系
在分析树木径向生长与气候因子相关关系的基础上,挑选出与超过50%的单根年表显著相关的气候因子进行进一步线性拟合分析,即年表与Pc6、Pp8c6以及Tc6相关系数绝对值与树木年龄、初始生长率进行线性拟合.对于年龄组的树木而言,年表与Pc6和Pp8c6相关系数绝对值与树龄间存在显著的线性关系,其中树龄与Pc6相关系数绝对值的线性关系要强于树龄与Pp8c6相关系数绝对值的关系,老龄树对Pc6和Pp8c6的响应要强于幼龄树.此外树木径向生长对Tc6的响应与树龄间不存在显著线性关系(图5).对于生长率组的树木而言,年表和Pp6、Pp8c6相关系数绝对值与初始生长率都存在着明显的线性关系,且P<0.01,结果表明初始生长快的树木对Pp6和Pp8c6的响应要强于初始生长慢的树木.年表与Tc6相关系数绝对值与初始生长率间也不存在显著线性关系(图6).
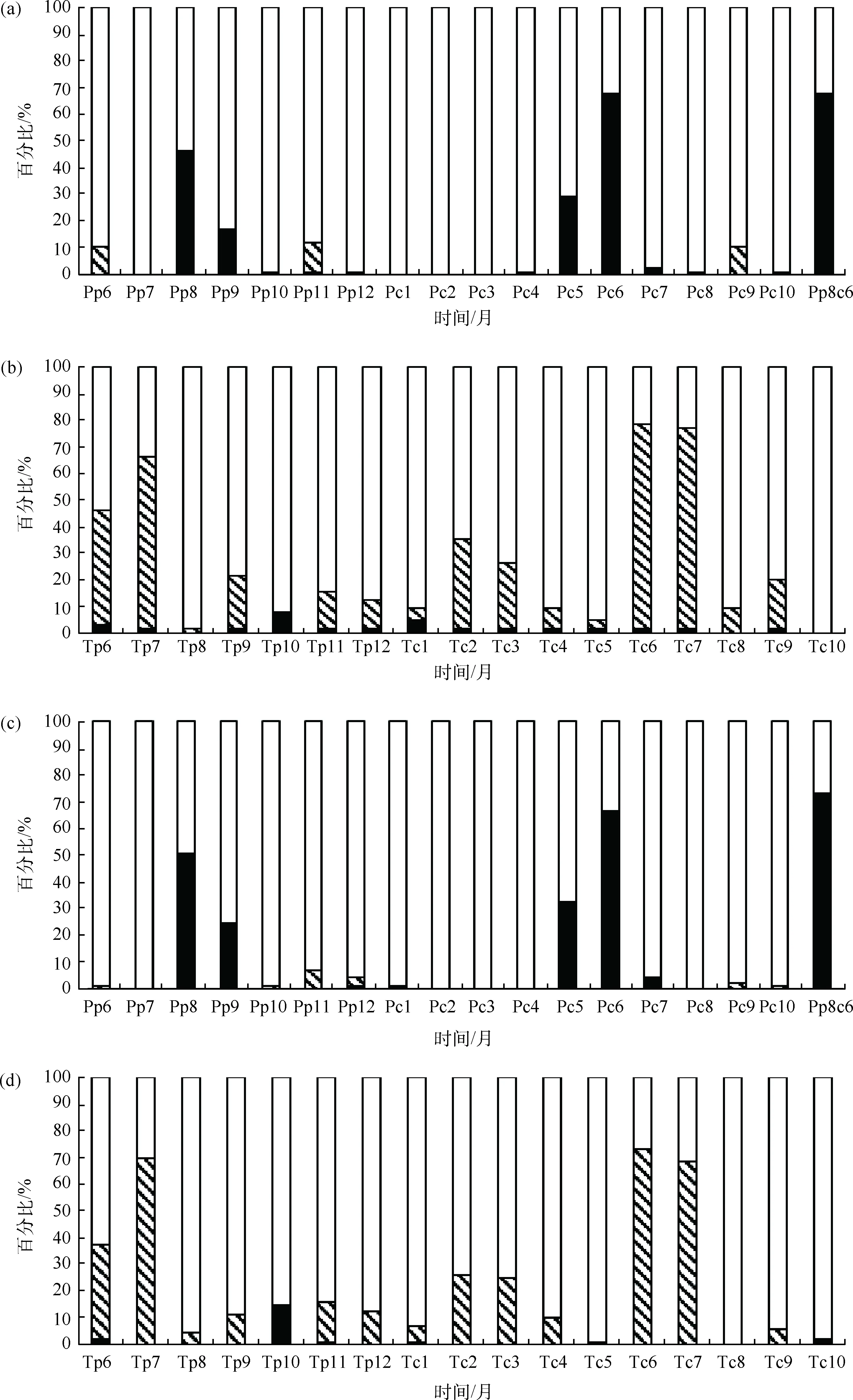
图4 不同组单根样芯与气候因子相关系数比例
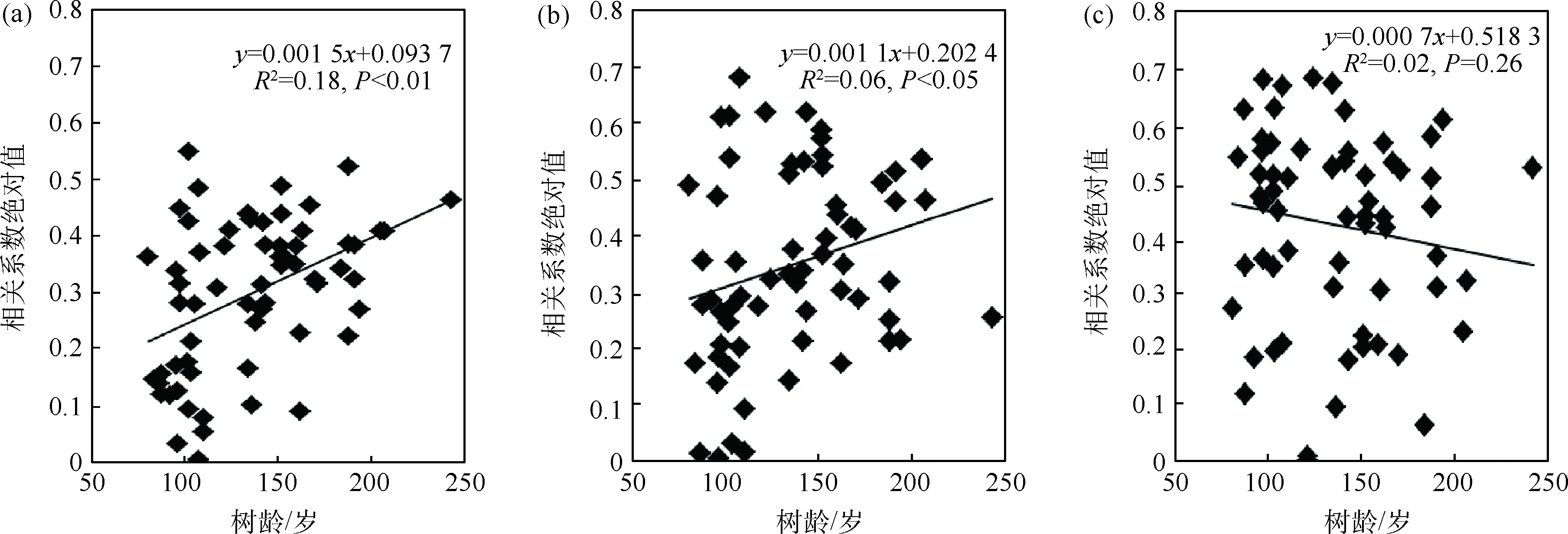
图5 年龄组单根年表与气候因子相关系数绝对值与树龄的关系

图6 生长率组单根年表与气候因子相关系数绝对值与初始生长率的关系
5 讨 论
5.1 树木径向生长对降水和温度因子的响应
结果表明年龄组和生长率组的树木均表现出与前一年8月、当年5、6月及前一年8月至当年6月累积降水有显著正相关关系,这说明前一年夏季降水、当年生长季初期降水以及从上一生长季中期到本生长季中期的累积降水均对本研究区树木有普遍的影响作用.前一年8月的丰沛降水可以缓解同时期高温引起的土壤干旱,降低树木遭受干旱胁迫的概率,使树木能储存更多的营养物质,为下一年生长提供有利条件[29].当年6月充沛的降水可以促进生长季初期形成层细胞的产生和分化,从而利于当年树木的径向生长[11].有研究发现祁连山树木径向生长与前一年夏季的降雨量呈显著正相关关系.祁连山区域的青海云杉均普遍表现出来与前一年8月、当年6—8月的降水显著的相关性[20,30-31].Song等[32]研究青藏高原东北部祁连圆柏(Sabinaprzewalskii)和祁连山中部寺大隆林场的青海云杉与降水因子关系时,发现树木与当年夏季初期降水有着显著的相关关系.前一年生长季中期到当年生长季中期充沛的累积降水对树木径向生长有明显的促进作用.同样,河西走廊地区和祁连山西部的青海云杉以及贺兰山油松(Pinustabulaeformis)也对前一年7月至当年6月的累积降水有着强烈响应,说明上一生长季中期到本生长季中期的累积降水对半干旱区和干旱区树木的径向生长有普遍的显著影响[33-35].
同时结果表明两组树木均表现出与前一年6、7月和当年6、7月的月平均温度有显著负相关关系,这说明前一年和当年夏季的高温会对本研究地树木径向生长造成明显的抑制作用.树木径向生长与温度之间的显著负相关关系可能是因为高温加强了土壤表面的蒸发作用从而加剧干旱,同时高温导致叶片气孔关闭,降低光合作用速率,减少碳水化合物的产生,从而导致当年形成的年轮较窄[36].类似的情况也出现在祁连山中部,即青海云杉径向生长与当年夏季前期的温度表现出显著负相关关系[37].在天山中部和西部的低海拔地区,树木生长也表现出与夏季温度显著负相关关系[38].
5.2 树木径向生长对气候因子响应强度与树龄的关系
对于年龄组的树木而言,老龄树比幼龄树对于前一年6月降水和前一年8月至当年6月的累积降水响应更加强烈.但树木径向生长与当年6月的月平均温度相关系数绝对值与树龄不存在显著线性关系.这可能是因为老龄树的光合作用要明显小于幼龄树,而老龄树自身复杂的生理结构会消耗更多营养物质,更容易受到外界降水因子的影响[39-40].Rossi等[41]的研究发现老龄树春季的活动会受到较粗的树干和较厚的树皮影响,因此老龄树运输水分时可能比幼龄树更困难,老龄树比幼龄树更容易受到降水的影响.类似的研究结果有,在奥地利阿尔卑斯山较为干旱的针叶混交林中,挪威云杉(Piceaabies)对5—6月的降水响应会随着树龄的增加而增强[11];对于罗马尼亚喀尔巴阡山的挪威云杉和加拿大西北部麦卡锡山和富兰克林山的黑云杉(Piceamariana)的研究都表现出老龄树对气候更敏感的特点[42-43].
径向生长与当年6月平均温度响应与树龄间不存在显著线性关系,说明高温对各年龄树木生长的抑制作用较为一致.Sun和Liu[44]将黄土高原东部油松划分为3个等年龄组,研究结果也表明各年龄组树木径向生长均表现出与前一年6月、当年6与7月的降水显著正相关关系,与当年6月平均温度负相关关系的特征.对阿尔卑斯山中部瑞士石松的研究结果显示,不同树龄年表与气候因子的相关关系无明显差别,由此认为树龄不影响树木生长对气候的响应,即树木径向生长对气候的敏感性与树龄无明显关系[45].
5.3 树木径向生长对气候因子响应强度与初始生长率的关系
研究显示生长率组树木的径向生长对前一年6和8月至当年6月累积降水的响应与初始生长率都存在着明显的线性关系,即初始生长快的树木对前一年6和8月至当年6月累积降水的响应比初始生长慢的树木更强.这可能是由于初始生长较快的树木自身具有较为发达的生理系统,对于降水有更加强烈的响应.但也有研究提出树木生长过快会导致地上生物量与地下生物量的比值太高,对碳水化合物的投入减少,从而使得抗病菌能力减弱,降低呼吸效率,影响树木对气候因子的响应[46].树木径向生长对当年6月的月平均温度的响应与初始生长率间不存在显著线性关系,表明不同生长率树木对温度的响应是较为一致的.这可能是由于在温度较高的夏季,水分缺失会抑制树木的生长,树木会选择在气候较为适宜的春季和早夏开始形成层活动并产生管胞或导管,使生长更加迅速[16].
5.4 树龄与初始生长率间的关系
分析树木树龄与初始生长率的关系显示,在本研究区树龄大的树木都具有较低的初始生长率,即初始生长慢的树通常比初始生长快的树能存活至更大的寿命.初始生长过慢的树木由于在生长前期将过多资源分配给木质部,用以防止病原菌的袭击,从而导致新陈代谢速率降低,抵抗力增强使树木寿命延长[47].初始生长快的树增加了短期内生存的可能性,但是对防御的投入较少,使得树木根部发育不良,易降低树木存活率[5].有研究也证明初始生长较快的树木因为养分好,竞争力强,有一定竞争优势.但这些生长较快的树木木材密度低,抗性差,因此更容易过早死亡,导致此部分抵抗力差的树木年龄普遍不高[4].
6 结 论
在本研究区内,年龄组和生长率组树木均表现出与前一年8月、当年5和6月及前一年8月至当年6月的累积降水有显著正相关关系,同时也表现出与前一年6、7月和当年6、7月的月平均温度有显著负相关关系.对于年龄组树木而言,老龄树比幼龄树对于降水的响应更加强烈,不同年龄树木对夏季中期温度的响应较为一致.对于生长率组的树木而言,初始生长快的树木对降水的响应比初始生长慢的树木更强烈,但不同初始生长率树木对夏季中期温度的响应亦无明显差别.研究区树木也表现出树木寿命越长其初始生长率越低的特点,而初始生长率低且年龄小的树木对降水的响应最不敏感.在森林经营管理中应对初始生长过快和树龄过大的树木加强管理和保护.
