贡嘎山海螺沟冰川退缩区原生演替不同阶段优势植物光合生理特征
2020-01-08杜流姗陆琦梁紫嫣类延宝
杜流姗 ,陆琦 ,梁紫嫣 ,类延宝*
1.中国科学院、水利部成都山地灾害与环境研究所山地表生过程与生态调控重点实验室,四川 成都 610041;2.中国科学院大学,北京 100049
植物光合生理特征能体现植物对不同生境的适应,反映植物对不同环境变化的响应(Jiang et al.,1999)。植物叶片积累了大部分的氮,约一半叶片氮用于光合作用,分配到光合系统中的氮素越多,植物生长越快(Onoda et al.,2004;Feng et al.,2009)。光合氮利用效率(photosynthetic nitrogen use efficiency,PNUE)指单位面积叶片光饱和速率与单位面积叶片氮含量的比值,是植物与生态特性相关的一种固有属性(Takashima et al.,2004),是描述植物叶片养分利用、生理特性和生存策略的重要特征,影响植物生长速率、竞争能力和叶片寿命等多方面的特性(Hikosaka,2004)。叶片的各项功能性状是影响PNUE的重要因素,同时也反应植物资源获取和利用的能力。陈世伟等(2015)对甘南亚高山草甸围封地群落演替不同阶段优势植物的研究发现植物净光合速率、水分利用效率(water use efficiency,WUE)随演替的进行呈降低趋势。入侵种大叶醉鱼草(Buddleja davidii)的PNUE显著高于乡土木本植物,有助于醉鱼草成功入侵(Feng et al.,2007);在入侵地与原产地生长的紫茎泽兰研究中也发现类似规律(Feng,2008)。在中国亚热带森林演替早、中和后期阶段的优势植物光合特性的研究中表明随着演替的进行植物的光饱和净光合速率(light saturated photosynthesis,Pmax)、比叶面积(specific leaf area,SLA)和PNUE呈现下降趋势,在森林演替过程物种的光合特性的变化体现森林演替过程中植物的生理特性(Xiao et al.,2018)。原生演替受人为因素和历史因素的干扰较少,对原生演替的研究能够更好的反应自然状况下植被的发展规律(Wiegleb et al.,2001)。目前,对冰川退缩区原生演替过程中植物光合生理特性的研究较少,而演替过程中植物的Pmax、PNUE等光合特性的变化及其影响因素可能是冰川退缩区原生演替过程中物种代替的重要机制,对原生演替过程中植物光合特性的研究具有重要意义。
全球气候变暖加快冰川退缩速度,贡嘎山海螺沟冰川经过130年的退缩演化出完整的从裸地到先锋群落再到顶级群落的连续植被原生演替序列(杨丹丹等,2015)。植被原生演替经历了裸地-草本地被、沙棘-高山柳、川滇柳-冬瓜杨、冬瓜杨-糙皮桦和云冷杉-杜鹃等几个阶段,并形成完整的可定龄的土壤发育序列和植被演替带谱(李逊等,1995;Li et al.,2010),为研究自然状态下的植被原生演替各种过程和机理提供了最理想场地和实验研究途径(吴艳宏等,2014)。本文以贡嘎山海螺沟冰川退缩区原生演替不同阶段的优势植物为研究对象,通过对植物各项光合生理特征等叶片功能性状的研究,探索原生演替不同阶段植物的光合生理特性对环境变化的响应及其生态策略,为认识高山植物种间相互作用、物种共存和群落构建机制等提供一定的科学依据。
1 材料和方法
1.1 研究地概况
研究区域贡嘎山(29°30′—30°20′N,101°30′—102°15′E)处于青藏高原寒带和温暖潮湿亚热带季风区域的过渡地带。海螺沟冰川处于贡嘎山东坡,为典型的季风海洋性山谷冰川。冰川退缩区在长约2000 m,宽50—200 m,海拔2850—3000 m(垂直高差仅150 m)的区域内演化出完整的植被演替带谱。该地年均气温为3.8 ℃,最冷月1月气温约为-4.4 ℃,最热月7月为11.9 ℃(海拔3000 m气象观测站数据),年降水量约为2000 mm,6—10月的降水量占全年降水量的80%以上,年平均相对湿度90%。
1.2 研究方法
1.2.1 样地选择与样品采集
贡嘎山海螺沟冰川退缩区原生演替经历了裸地-草本-灌木-乔木-针叶林等几个阶段(杨丹丹等,2015;Lei et al.,2015),选取冰川退缩后 3—12、40—80、120 a分别代表原生演替的早期、中期和后期。演替早期优势植物为黄芪(Astragalus membranaceus)、沙棘(Hippophae rhamnoides)和长叶柳(Salix phanera),演替中期优势植物为冬瓜杨(Populus purdomii)、川滇柳(Salix rehderiana)和大叶柳(Salix magnifica),演替后期优势植物为峨眉冷杉(Abies fabri)、麦吊云杉(Picea brachytyla)和大白杜鹃(Rhododendron decorum)。每个阶段的代表种选取 3株植物,测定其光合参数后,采集长势相近的叶片30片,用于比叶面积的测定和叶片碳氮元素含量的分析。每个演替阶段设置3个5 m×5 m的样方,每个样方采集5份土壤后混合成一份土壤样品,用于测定不同演替阶段土壤的基本理化性质。
1.2.2 光合参数的测定
在植物的生长季(2018年8月13—20日)天气晴朗的上午 9:00—11:30,使用 Li-6400型光合仪(Li-6400XT,Biosciences,Lincoln,NE,USA)对不同演替阶段的优势植株进行光合参数的测定。演替中期和后期植株较高,每棵树木用高枝剪从顶端向阳位置剪下3—5根1—2 m长的健康树枝,选取最好的一枝即刻放入装满清水的水桶中,随后剪掉树枝的低端以防止气体栓塞(唐敬超,2017)。
气体交换参数设定,(1)光合响应曲线:利用CO2小钢瓶控制参比室 CO2浓度,设定值为 400µmol·mol-1,流速设定为 500 μmol·s-1,温度为(25±1) ℃,Li-6400型光合仪红蓝光源,选择阳光充足、长势良好、无病虫害、完全成熟的当年生叶片进行测定,净光合速率数据保持平衡大约3 min进行记数。(2)二氧化碳响应曲线:设置饱和光合光强为1200 μmol·m-2·s-1PPFD,利用 CO2小钢瓶改变参比室CO2浓度,其他各项参数与测量光合曲线相同。通过测量可直接获取基于面积的净光合速率(Pn)、蒸腾速率(Tr)和气孔导度(Gs)等数据。光合氮利用效率(PNUE)=光饱和净光合速率(Pmax)/单位面积叶片氮含量(Narea),水分利用效率(WUE)=光饱和净光合速率(Pmax)/蒸腾速率(Tr)。
1.2.3 比叶面积测定
将带回实验室的植物叶片样品,用扫描仪扫描叶片面积后,利用 ImageJ软件处理并计算叶片面积,将叶片放入信封中,50 ℃下烘至干质量不变,比叶面积(SLA)=叶片面积(cm2)/叶片干质量(g)。
1.2.4 叶氮分配参数计算
叶氮在光合系统各组分的分配系数使用Niinemets et al.(1997)提出的方法计算:
叶氮在羧化组分(主要是Rubisco)中的分配比例PC:

叶氮在生物力能学组分(主要为光合磷酸化载体和光合电子传递)中的分配比例PB:

其中:Narea是单位面积叶片氮质量(g·m-2);Vcr是单位 Rubisco 蛋白的固定活性,取 20.78 μmol·g-1;Jmc为单位细胞色素f(Cytf)的最大电子传递速率,取 155.65 μmol。Vcmax(maximum carboxylation rate)为最大羧化速率,Jmax(maximum electron transport rate)为最大电子传递速率,参考 Farquhar et al.(1980)和Loustau et al.(1999)提出的方法计算。
1.3 数据分析
分别采用SPSS 18.0(SPSS,Chicago,IL,USA)和Origin 8.0(Origin,Northampton,USA)进行统计分析和作图。所有的数据变量在分析之前均进行正态分布和方差齐性检验。利用单因素方差分析检验不同演替阶段优势植物各项参数的差异(One-way ANOVA),平均数之间的多重比较采用邓肯检验。使用线性回归分析叶片功能性状之间的关系,Pearson’s相关系数评估叶片功能性状之间的相关性。
2 结果与分析
2.1 不同演替阶段土壤的基本理化性质
由表1可知,原生演替早期和演替中期土壤的pH无显著差异(P=0.055),演替后期土壤的pH显著降低(P<0.01),早期、中期和后期土壤的pH的平均值分别为6.50、5.97和4.73。不同演替阶段土壤的体积质量(容重)无显著差异(P=0.328),早期、中期和后期土壤容重(g·cm-3)的平均值分别为1.64、1.32和1.17。原生演替过程中土壤的有机质含量显著增加(P<0.01),早期、中期和后期土壤的有机质含量(g·kg-1)的平均值分别为1.50、2.30和3.50。演替早期、中期土壤的无机氮含量无显著差异(P=0.624),演替后期土壤的无机氮含量显著增加(P<0.01),早期、中期和后期土壤的无机氮含量的平均值分别为 115.20、107.91、254.97 mg·kg-1。不同演替阶段土壤的有效磷含量具有显著性差异(P<0.01),早期、中期和后期土壤的有效磷含量的平均值分别为 6.74、48.47、40.46 mg·kg-1,演替中期土壤的有效磷含量为演替早期的7.19倍。
2.2 不同演替阶段优势植物叶片基本理化性质
由表2可知,演替早期优势植物的叶片干物质含量(leaf dry matter content,LDMC)显著低于演替中期和后期优势植物的LDMC(P<0.01),演替中期和后期优势植物的 LDMC无显著性差异(P=0.972),早期、中期和后期优势植物的 LDMC(%)的平均值分别为25.90、38.50和38.37。原生演替早期到后期优势植物的 SLA显著降低(P<0.01),早期、中期和后期优势植物的 SLA(cm2·g-1)的平均值分别为 176.84、141.45 和 103.21。演替早期和中期优势植物的Cmass无显著性差异(P=0.773),演替后期优势植物叶片碳含量(Cmass)显著增加(P<0.01),早期、中期、后期优势植物的Cmass(mg·g-1)的平均值分别为 455.61、453.57 和499.46。不同演替阶段优势植物的Narea无显著差异(P=0.065),早期、中期和后期优势植物的 Narea(mg·g-1)的平均值分别为1.69、1.64和1.33。原生演替早期到后期优势植物的 Nmass呈显著下降趋势(P<0.01),早期、中期和后期优势植物的 Nmass(mg·g-1)的平均值分别为30.69、22.27和13.39。

表1 不同演替阶段土壤pH、土壤体积质量(容重)和有机质、无机氮、有效磷质量分数Table 1 Soil pH, bulk density, and mass fractions of soil organic material, inorganic nitrogen, and available phosphorus at different successional stages
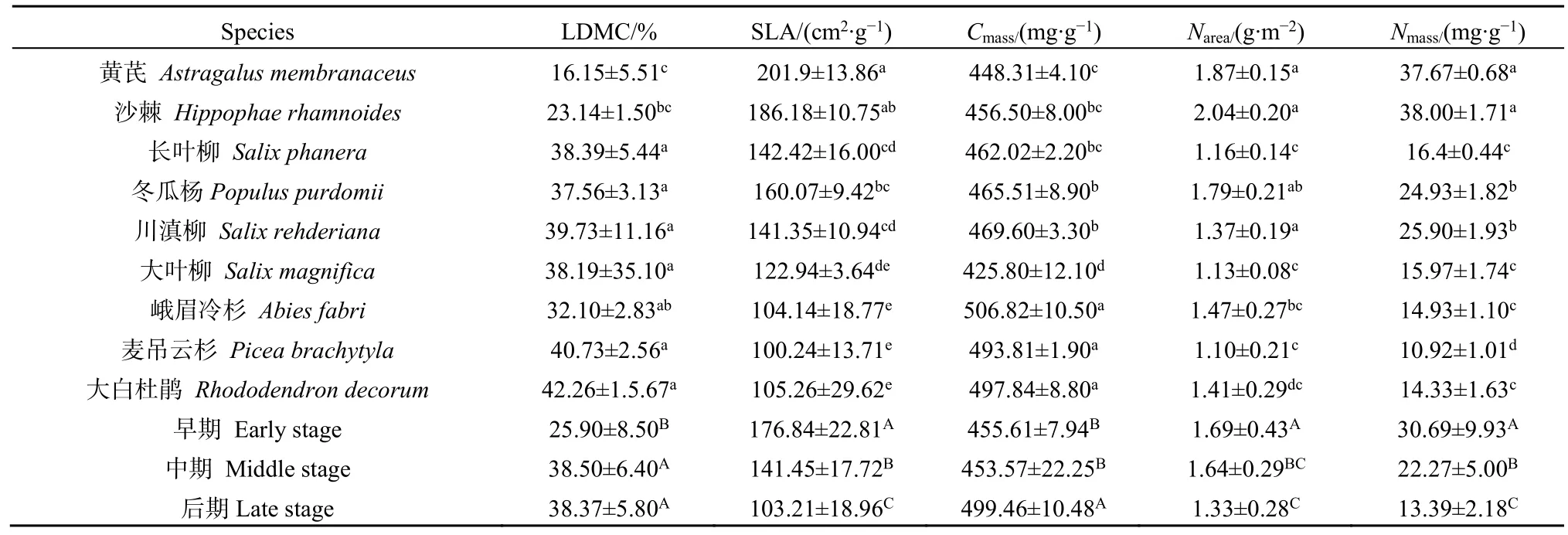
表2 不同演替阶段优势植物叶片干物质量分数、比叶面积、叶片碳质量分数、单位面积氮质量和叶片氮质量分数Table 2 Leaf dry mass fraction (LDMC), specific leaf area (SLA), carbon mass fraction (Cmass), and nitrogen mass fractions of unit area and leaf (Narea and Nmass) of dominant species at different successional stages
2.3 不同演替阶段优势植物光合特性
如图1所示,演替早期到后期优势植物的Pmax和PNUE呈显著降低趋势(P<0.01,图1A和B),早期、中期和后期优势植物的 Pmax(μmol·m-2·s-1)的平均值分别为 27.30、22.37和 9.53,PNUE(μmol·g-1·s-1)的平均值分别为 16.66、13.87 和 7.48。不同演替阶段优势植物的 WUE率无显著差异(P=0.274,图1C),早期、中期和后期优势植物的WUE(mmol·mol-1)的平均值分别为 8.21、8.14 和6.56。演替早期到后期优势植物叶片气孔导度(stomatal conductance,Gs)显著降低(P<0.05,图1D),早期、中期和后期优势植物的Gs(mol·m-2·s-1)的平均值分别为0.25、0.19和0.14。演替早期和中期优势植物 Vcmax和 Jmax无显著差异(P=0.141和P=0.185),演替后期显著降低(P<0.01,图 2A、B)。早期、中期和后期优势植物的 Vcmax(μmol·m-2·s-1)的平均值分别为 72.16、62.63和 43.98,Jmax(μmol·m-2·s-1)的平均值分别为 108.40、95.00 和68.51。演替早期到后期优势植物叶片的羧化组分氮分配比例(PC)和生物力能学组分氮分配比例(PB)呈显著降低趋势(P<0.05,图2C、D),早期、中期和后期优势植物的PC(g·g-1)的平均值分别为0.33、0.29和 0.26,PB(g·g-1)的平均值分别为 0.051、0.046和0.041。
2.4 不同演替阶段植物光合生理特征间的相关性
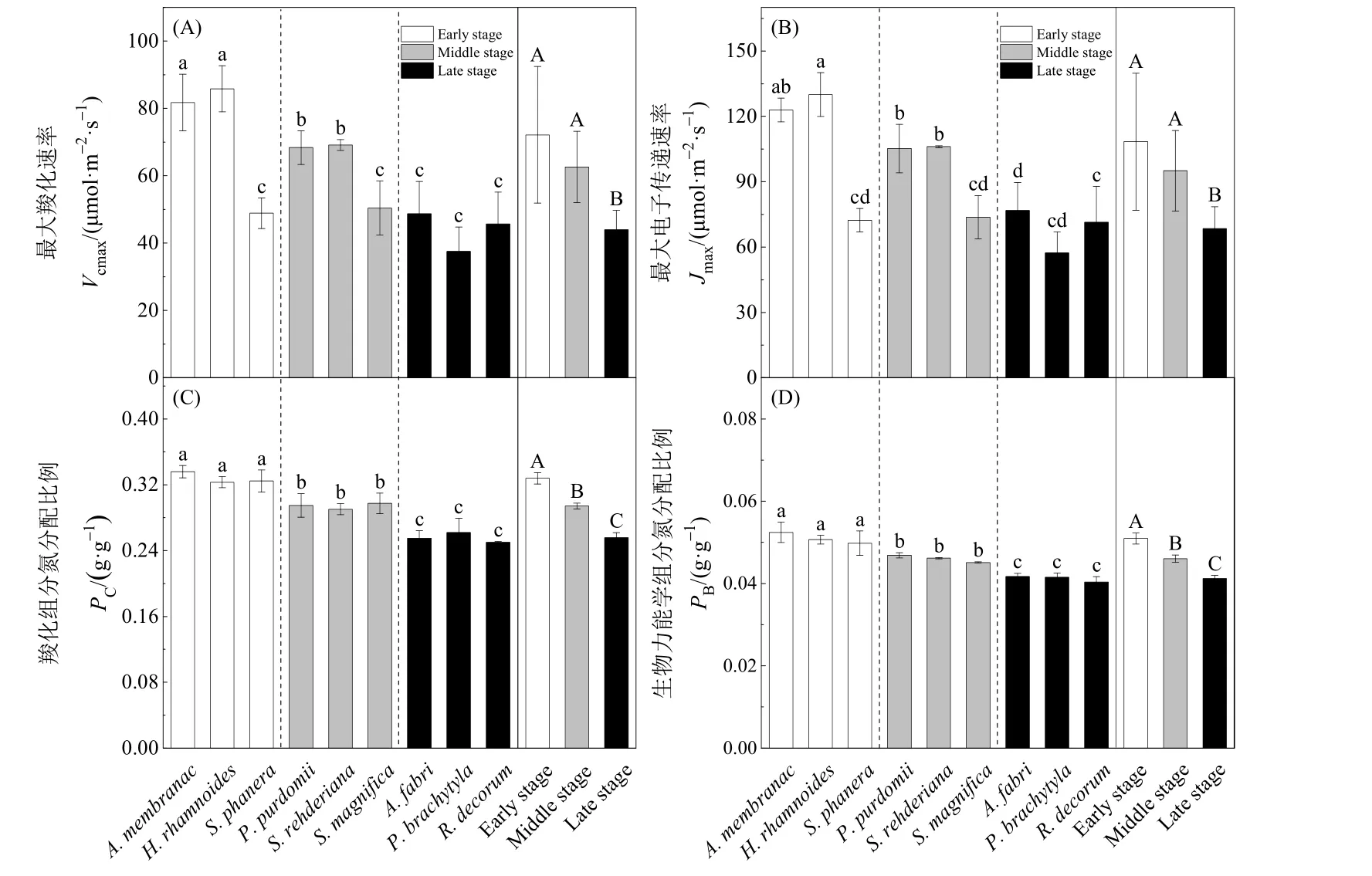
图2 不同演替阶段优势植物最大羧化速率(A)、最大电子传递速率(B)、羧化组分氮分配比例(C)、生物力能学组分氮分配比例(D)Fig. 2 Mean values (±SD) of maximum carboxylation rate (Vcmax, A), maximum electron transport rate (Jmax, B), the fraction of leaf nitrogen allocated to Rubisco (PC, C) and bioenergetics (PB, D) for dominant species at different successional stages

表3 叶片性状之间的相互关系Table 3 The coefficient matrix among leaf functional traits
由表3可知,原生演替过程中优势植物的叶片功能性状之间具有一定的相关性:优势植物的Pmax与SLA和Nmass呈显著正相关(P<0.01,相关系数(r)分别为 0.88和 0.86),与 WUE呈正相关(P=0.050,r=0.41),与 LDMC呈显著负相关(P<0.01,r=-0.70)。SLA 与 Nmass呈显著正相关(P<0.01,r=0.89),与WUE呈正相关(P=0.023,r=0.44),与LDMC 呈负相关(P<0.05,r=-0.72),Nmass与WUE呈正相关(P=0.019,r=0.45),与LDMC呈显著负相关(P<0.01,r=-0.72),LDMC与WUE之间的相关性不显著(P>0.05)。如图3所示,原生演替过程中优势植物的 PNUE随着优势植物的SLA、Pmax、PC和PB的增加而增加,呈显著的正相关关系(P<0.01),r分别为 0.67、0.78、0.85和 0.84。
3 讨论
随着原生演替的进行,影响植物生长的环境因子发生一定的变化,植物采取不同的策略适应环境变化,表现为不同阶段植物具有不同的光合生理特性等叶片功能性状。Cmass和LDMC能够反映植物的固碳潜力和植物对资源的获取能力,是植物适应环境变化的重要指标(Boerner,1984;李玉霖等,2005)。演替后期优势植物叶片的Cmass和LDMC显著大于演替早期优势植物的Cmass和 LDMC,可能与演替早期优势植物快速的营养循环以供给植物合成利用,演替后期优势植物利用增加自身营养的储存效率来提高自身的竞争力有关(柴永福,2013)。植物的SLA是植物在生长过程中资源获取策略的重要功能性状(胡耀升等,2015),与植物的生长速率密切相关,尤其是在低光照的环境中(Poorter et al.,1999)。演替早期优势植物(黄芪和沙棘)具有较高的SLA,植物具有更快的生长速率。演替后期优势植物(云杉和冷杉)的SLA较小,部分原因是具有相对高的细胞壁物质来抵御有害的非生物和生物胁迫(Lambers et al.,1992)。光合作用能力是植物基本的属性,反映植物的生长快慢和资源利用的能力。演替早期黄芪的Pmax高于沙棘、长叶柳等灌木和冬瓜杨、川滇柳等乔木植物的Pmax,与Wright et al.(2010)的研究结果一致。Hikosaka et al.(1998)也发现在相同的 Narea下,草本植物Chenopodium album的光合速率是常绿树种Quercus myrsinaefolia的二倍。贡嘎山海螺沟冰川退缩区植被原生演替过程中,从演替早期草本植物到演替后期针叶林群落,优势植物光合速率显著下降,与Xiao et al.(2018)对中国亚热带森林演替优势植物光合速率的研究结果相似。
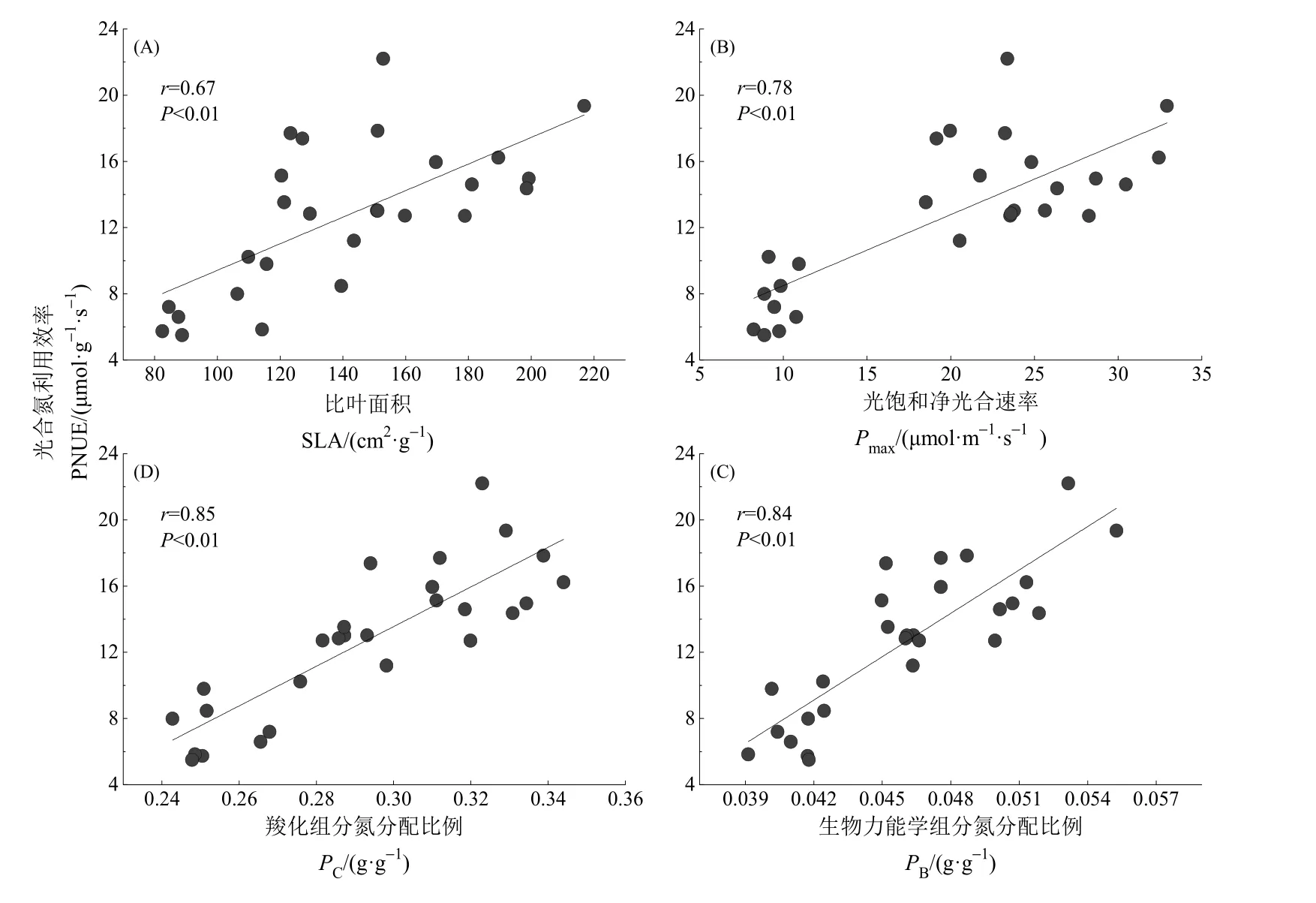
图3 不同演替阶段优势植物比叶面积(A)、光饱和净光合速率(B)、羧化组分氮分配比例(C)、生物力能学组分氮分配比例(D)与光合氮利用效率(PNUE)的相关性Fig. 3 Relationship between specif i c leaf area (SLA, A), light saturated photosynthetic rate (Pmax, B), the fraction of leaf nitrogen allocated to carboxylation(PC, C), bioenergetics (PB, D) with photosynthetic nitrogen use eff i ciency (PNUE) for dominant species at different successional stages
原生演替过程优势植物的SLA与Pmax呈正相关关系,与Benomar et al.(2011)的研究结果相似,高的 SLA可能通过影响叶片氮分配进一步影响Pmax(Feng,2008)。已有较多研究证明叶片氮含量与植物的光合特性具有较强的相关性(Harrison et al.,2009)。Ellsworth et al.(1996)的研究显示植物叶片氮含量随演替进行逐渐降低,与本研究结果一致。原生演替过程中优势植物的Pmax与Nmass呈正相关关系,植物叶片氮含量越高,更多的氮被分配到光合器官中,表现为从演替早期到后期优势植物的Nmass、PC和PB显著降低,演替早期优势植物具有更高的Pmax。贡嘎山海螺沟冰川退缩区具有较高的年降雨量,原生演替区域的水分条件能够满足植物对水分的需求,在Gs较低的情况下优势植物提高自身的WUE更好的进行光合作用,表现为优势植物的WUE与Pmax呈正相关关系,而在干旱条件上植物则通过增加Gs以增加植物光合作用能力,导致WUE 下降(Huang,2008)。
PUNE是能综合反映生理和适应策略的重要的叶片经济学性状(Feng,2008;Onoda et al.,2017)。原生演替过程中优势植物的PNUE显著降低,类似结果也在中国亚热带森林和温带森林的演替过程中得到验证(Chai et al.,2015;Xiao et al.,2018)。植物的比叶面积和叶片氮素在光合器官各组分间(羧化组分、生物力能学组分)的分配等是影响PNUE 的主要因素(Feng et al.,2009;Moon et al.,2015)。原生演替过程中优势植物的SLA、PC和PB呈下降趋势且与PNUE呈显著的正相关关系,验证了落叶树种和具有高 SLA的植物通常分配更多的氮到光合器官中,比常绿植物和低SLA的植物拥有更高的PNUE(Takashima et al.,2004)。演替早期土壤营养匮乏,能够固定大气中的氮素的豆科植物定殖在营养匮乏的土壤上,改变土壤性质(Titus,2009),为演替中期的植物提供较适宜的环境;演替中期的植物一般具有较高的 SLA、光合能力和PNUE,表现为“以快制胜”的生存策略;演替后期的物种虽然光合能力和PNUE降低,但大多具有更长的叶片寿命,因而具有更高的成本-收益(costbenefit),从而更具竞争优势(数据待发表)。同时,植物对氮素的吸收和利用具有一定的偏好性,根系形态、菌根共生以及偏好利用等都影响植物养分策略,与土壤环境、菌根类型相适应,能协同进化的植物逐渐成为优势物种(Chen et al.,2016)。
4 结论与展望
贡嘎山海螺沟冰川退缩区植被原生演替过程中优势植物叶片的Nmass、SLA、Pmax、PNUE、PC和PB显著下降(P<0.01),叶片功能性状之间具有显著的相关性:Pmax与SLA、Nmass和Gs呈显著正相关(P<0.01),PNUE 与 SLA、Pmax、PC和 PB呈显著正相关(P<0.01)。原生演替过程中具有与环境相适应的光合特性和叶片功能性状的植物逐渐成为不同演替阶段的优势种植物,光合生理特征的变化和叶片功能性状之间的联系可能代表了植被原生演替进行的基本生理机制。本研究中只分析了冰川退缩区原生演替过程中植物光合特性和氮素利用等地上功能性状,在未来的研究中,将进一步从植物磷利用等方面加深研究;并聚焦地下微生物,尤其是共生的根真菌类型,以期揭示植被演替、土壤发育与微生物群落的耦合关系和协同效应,从而为认知高山植物种间相互作用、群落的构建机制及环境变化下植被演替动态提供科学依据。
