秦岭松栎林土壤生态化学计量特征及其对海拔梯度的响应
2020-01-08吴昊邹梦茹王思芊万洪秀
吴昊 ,邹梦茹,王思芊,万洪秀
1. 信阳师范学院生命科学学院,河南 信阳 464000;2. 西北农林科技大学林学院,陕西 杨凌 712100;3. 中国科学院南京地理与湖泊研究所,江苏 南京 210008
土壤养分是土壤肥力的物质基础,土壤中的碳(C)、氮(N)、磷(P)等重要生命元素是植物体内元素的主要来源,其含量及分布状况决定了地上植物生长、植被结构、生产力水平和生态系统稳定性(Vitousek et al.,2010;李红林等,2015;吴昊,2015a)。生态化学计量学(Ecological stoichiometry)通过分析多重化学元素(主要是C、N、P)质量平衡对生态交互作用的影响来探究生态系统结构与功能(Elser et al.,2000),被广泛应用于植物养分利用效率、植物-土壤营养元素耦合关系、土壤元素化学计量空间分异性等诸多方面的研究中(Han et al.,2005;卢同平等,2017;Sardans et al.,2017)。其中,土壤生态化学计量作为表征土壤内部养分循环和对植物养分供应状况的指标,对于揭示元素的可获得性及其循环与平衡机制、生态过程对全球变化的响应等科学问题具有重要意义(王绍强等,2008;李红林等,2015)。
诸多学者已围绕土壤生态化学计量特征的空间分布格局、土壤化学计量特征对植物生产力的影响、土壤-植物-枯落物的化学计量耦合关系以及土壤生态化学计量学的驱动因素等方向展开大量研究(Yuan et al.,2012;杨佳佳等,2014;卢同平等,2017)。近年来,环境梯度上的土壤生态化学计量特征变化引起研究者的广泛兴趣并逐渐成为研究热点。如:研究发现,我国东北地区阔叶红松林土壤中的 C꞉N、C꞉P均随纬度升高显著增加,而黄土高原刺槐林土壤的 C꞉N、N꞉P随纬度升高均呈指数递减趋势(张向茹等,2013;李喜霞等,2018);太白山阔叶林带土壤的C꞉N随海拔增高呈现降低趋势,N꞉P先上升后下降(李丹维等,2017);随着海拔增加,天山北坡土壤的C꞉P和N꞉P均先升高后降低,且其土壤化学计量特征在不同植被类型之间差异显著(谢锦等,2016)。相较于其他生态系统,海拔梯度对森林土壤生态化学计量特征的决定性作用尤为显著,这是由于海拔变化通常会造成森林土壤理化性质、植被分布和凋落物量的显著改变,进而影响森林碳库大小及其土壤养分供应状况(向慧敏等,2015;李丹维等,2017)。深入探讨森林土壤计量特征对海拔变化的响应有助于理解土壤元素的生物地球化学循环过程及森林生态系统的功能维持机制。
松栎混交林是秦岭最具典型地带性的植物群落之一,其分布面积占整个林区40%以上。其中,广泛分布于 1000—1800 m 之间的油松(Pinus tabuliformis)—锐齿槲栎(Quercus aliena var.accuteserrata)林是最主要的松栎混交类型,对维护秦岭群落结构稳定性及生态系统功能的发挥具有重要作用(吴昊,2015b)。针叶林混交阔叶树是改善林地土壤肥力、增强生态系统养分循环的重要经营措施(徐芷君等,2019)。以往关于该林地土壤养分的研究主要集中于养分含量空间变异、土壤理化性质随海拔变化趋势、土壤营养对植物群落物种多样性的影响等方面(张巧明等,2011;吴昊等,2012;吴昊,2015a),而关于林地土壤生态化学计量的研究较少。本研究通过对秦岭油松-锐齿槲栎混交林土壤的野外调查与取样分析,探讨不同土层中C꞉N、C꞉P和N꞉P化学计量特征沿海拔梯度的变化趋势,以期为秦岭森林土壤养分优化与管理及森林可持续经营提供参考依据。
1 研究区概况
研究区位于陕西省境内秦岭南坡中段的火地塘和旬阳坝林区。火地塘海拔1450—2470 m,年均温8—10 ℃,年降水量1000—1200 mm。土壤以黄棕壤、棕壤和暗棕壤为主,土层厚度 45—105 cm,土壤偏酸性。旬阳坝平均海拔1300 m,年均温10 ℃,年均降水量1133 mm。土壤主要为砾质黏土,pH值呈微酸性。研究区内松栎混交林在20世纪 60—70年代受过人工皆伐的干扰,其后经过长时间封山育林及天然林保护,现存植被多为恢复起来的天然次生林,部分地段存在一定面积的人工林;林地面积较为集中,主要分布于低中山地带,林龄平均为58 a,林地群落处于“油松林—松林混交林—锐齿槲栎林”这一次生演替序列的中间阶段(张洪武等,2014;张明霞等,2015)。群落植物物种组成丰富,乔木层除了建群种油松、锐齿槲栎之外,还共存有华山松(Pinus armandii)、漆树(Toxicodendron vernicifluum)、梾木(Cornus macrophylla)等树种。灌、草层中广泛分布鞘柄菝葜(Smilax stans)、五味子(Schisandra chinensis)、忍冬(Lonicera japonica)、崖棕(Carex siderosticta)、细叶苔草(Carex rigescens)、冰草(Agropyron cristatum)和铁线蕨(Adiantum capillus-veneris)等多种植物(吴昊,2015;2017)。此外,研究区松栎林群落的垂直结构明显,大体上可划分为8类群丛,其中“油松+锐齿栎-五味子-细叶苔草+铁线蕨”是秦岭南坡中段油松—锐齿槲栎林群落的主要混交类型(吴昊等,2012)。
2 研究方法
2.1 土壤采集
通过采用典型取样方法进行野外综合调查,于植物生长旺盛的 7—8月,采集研究区 1164—1810 m海拔范围内油松-锐齿槲栎混交林群落的土样。按照海拔每上升200 m设置样地,每个样地面积为20 m×20 m。共设置油松-锐齿槲栎混交林样地23个,具体海拔梯度分布为:1100—1300 m,4个;1300—1500 m,5个;1500—1700 m,9个;1700—1900 m,5个(表1)。在对混交林的植物进行常规群落学调查之后,沿着每个样地任意一条对角线随机选择 3个样点挖掘土壤剖面,分层采集 0—20、20—40、40—60 cm土层的土样。每个土样取250 g,将各样地土样按层混合装入土壤袋,及时带回实验室风干备用。使用手持GPS定位仪和罗盘仪记录样地的海拔、坡度、坡位、坡向等地形因子。
2.2 土壤测定
土壤样品自然风干后,经研磨过100目筛,按常规方法测定土壤 C、N、P元素含量:其中,采用重铬酸钾氧化—外加热法测定土壤总有机C;凯氏定氮法测定土壤总N;硫酸—高氯酸溶解法测定总P(吴昊,2015a;李丹维等,2017)。
2.3 数据处理
利用SPSS 16.0软件计算不同海拔梯度土壤的总有机C、总 N、总P以及C꞉N、C꞉P、N꞉P等指标的平均值(Mean)、标准差(SD)和变异系数(CV)。异系数(CV)=标准差÷平均值,CV<0.1为弱变异性,0.1≤CV≤1.0为中等变异性,CV>1.0为强变异性。利用单因素方差分析(One-way ANOVA)和最小显著差数法(Least Significant Difference,LSD)多重比较探讨23个样地土壤整体的C、N、P含量及其化学计量比在不同海拔梯度(低海拔:1100—1300 m;中海拔:1300—1500 m;中高海拔:1500—1700 m;高海拔:1700—1900 m)之间的差异、不同土层深度(0—20、20—40、40—60 cm)之间各自C꞉N、C꞉P、N꞉P的差异、以及不同土层中C、N、P化学计量比在4个海拔梯度之间的差异。利用线性回归拟合法(Linear Regression)探讨土壤总C、总N和总P之间的显著性耦合关系。
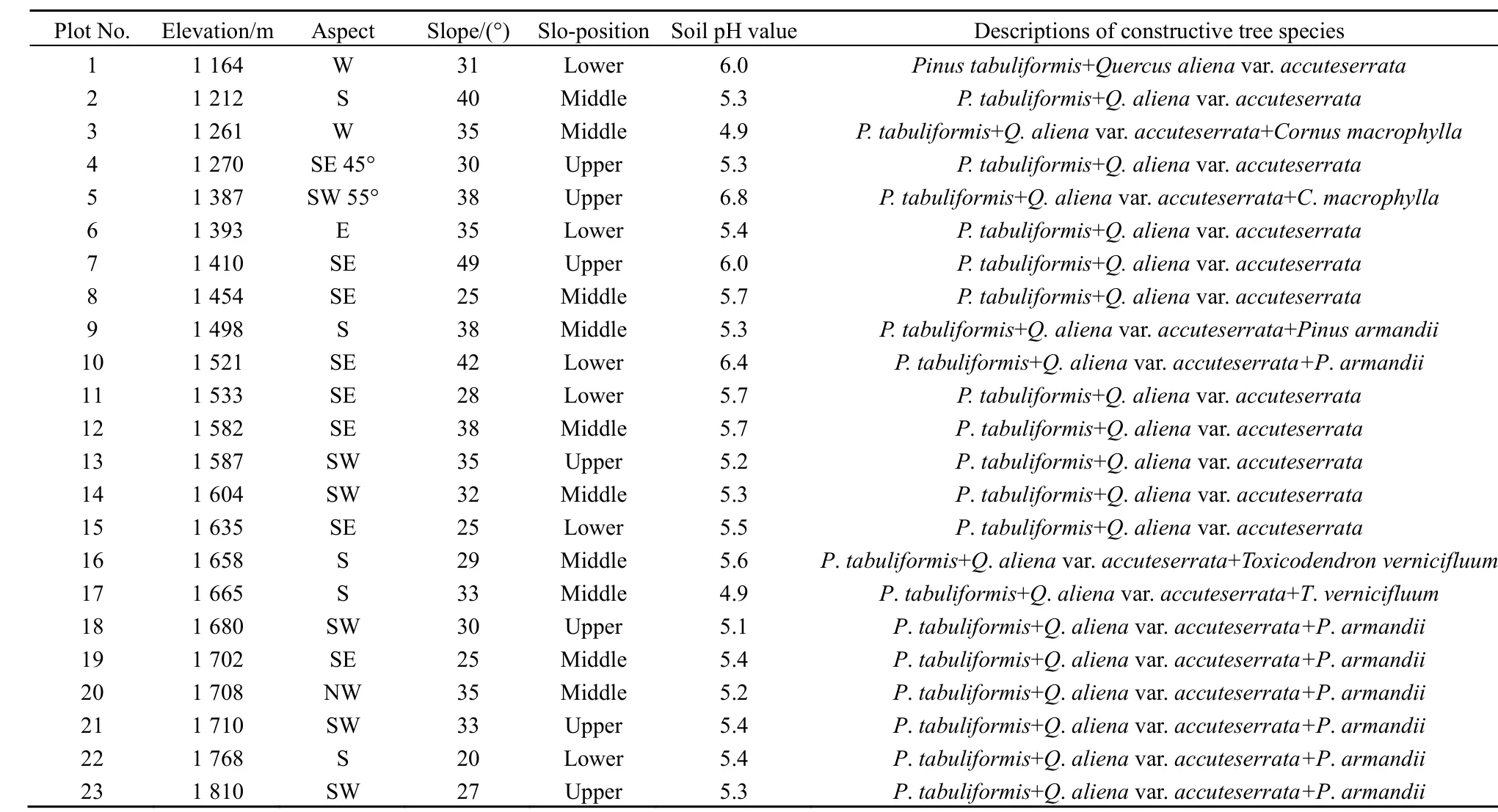
表1 研究区样地群落的环境因子及建群种概况Table 1 The environmental factors andconstructive species of sampling plot community in study area
3 结果与分析
3.1 不同海拔梯度土壤的C、N、P含量及其生态化学计量特征
由表2可知,土壤总有机C含量介于10.473—49.439 g·kg-1,均值为 23.723 g·kg-1,最小值和最大值分别位于海拔1393 m和1521 m处;总有机C在海拔 1500—1700 m 范围内的变异系数最高(CV=0.277),表现为中等变异性,在海拔 1100—1300 m内的变异系数最低,为(CV=0.087),表现为弱变异性。土壤总 N含量介于 0.537—2.477 g·kg-1,均值为 1.641 g·kg-1,其最小值和最大值分别位于海拔1393 m和1710 m处;总N在海拔1300—1500 m范围内变异系数最高(CV=0.295),在海拔1100—1300 m内变异系数最低(CV=0.173),均表现为中等变异。土壤总 P含量介于 0.563—1.917 g·kg-1,均值为 1.039 g·kg-1,其最小值和最大值分别位于海拔1261 m和1582 m处;总P在海拔 1500—1700 m范围内变异系数最高(CV=0.323),在海拔1300—1500 m内变异系数最低(CV=0.215),均为中等变异。
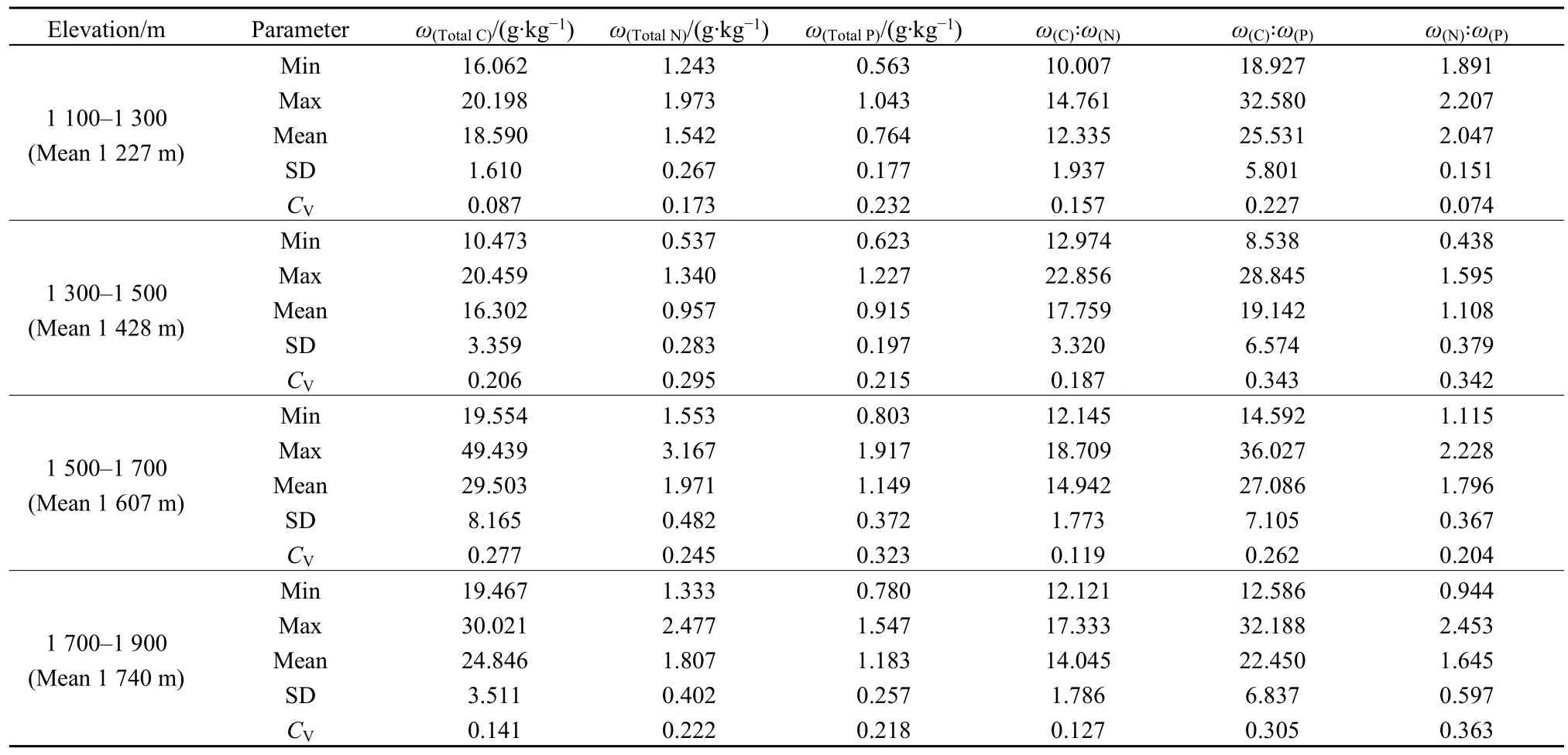
表2 不同海拔梯度土壤C、N、P生态化学计量特征的描述统计Table 2 Statistics of soil C, N, P eco-stoichiometry characteristics in different elevation gradients
化学计量比统计结果表明(表 2):土壤 C꞉N介于10.007—22.856,均值为14.906,最小值和最大值分别位于海拔1164 m和1410 m处;C꞉N在海拔1300—1500 m内变异系数最高(CV=0.187),在海拔1500—1700 m内变异系数最低(CV=0.119),均为中等变异。土壤C꞉P介于8.538—36.027,均值为24.081,最小值和最大值分别位于海拔1393 m和1604 m处;C꞉P在海拔1300—1500 m内变异系数最高(CV=0.343),在海拔1100—1300 m内变异系数最低(CV=0.227),均呈中等变异。土壤 N꞉P介于0.438—2.453,均值为1.657,最小值和最大值分别位于海拔1393 m和1708 m处;N꞉P在海拔1700—1900 m内变异系数最高(CV=0.363),为中等变异,在海拔1100—1300 m内变异系数最低(CV=0.074),为弱变异性。
单因素方差分析表明(图 1):土壤总有机 C(F3,22=5.980,P=0.005)、总 N(F3,22=6.178,P=0.004)在不同海拔梯度之间均存在显著性差异,整体表现为中、高海拔土壤总C、总N含量大于低海拔;土壤 C꞉N(F3,22=3.924,P=0.025)、N꞉P(F3,22=3.811,P=0.027)在不同海拔梯度间也存在显著性差异,表现为海拔 1300—1500 m 的 C꞉N 显著高于海拔1100—1300 m和1700—1900 m,而海拔1300—1500 m 的 N꞉P显著低于海拔 1100—1300 m 和1500—1700 m;土壤总P(F3,22=1.980,P=0.151)及C꞉P(F3,22=1.372,P=0.282)在不同海拔梯度之间均无显著性差异。
3.2 不同土层土壤生态化学计量特征及 C、N、P相关性
对不同土层的土壤C꞉N、C꞉P和N꞉P分别进行单因素方差分析(图 2),结果表明:不同土层的C꞉N之间无显著性差异(F2,68=0.150,P=0.861);不同土层C꞉P之间存在显著性差异(F2,68=13.681,P<0.001),0—20 cm表层土壤的C꞉P显著高于中层和下层土壤,20—40 cm和40—60 cm土壤C꞉P无显著性差异;不同土层N꞉P之间存在显著性差异(F2,68=14.337,P<0.001),0—20 cm 表层土壤的N꞉P显著高于20—40 cm中层和40—60 cm下层土壤,而中、下层土壤之间的N꞉P无显著性差异。
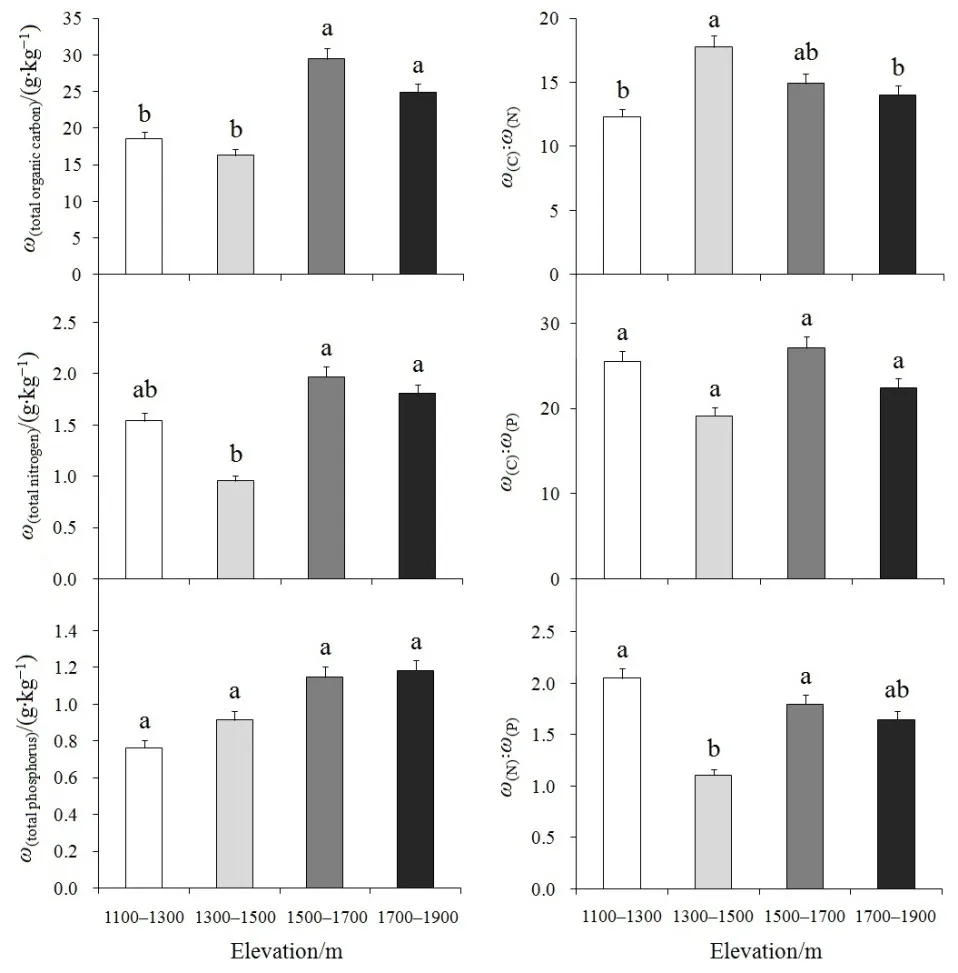
图1 不同海拔梯度土壤C、N、P及其生态化学计量特征的多重比较Fig. 1 Multiple comparisons of C, N, P and their eco-stoichiometry at different elevation gradients
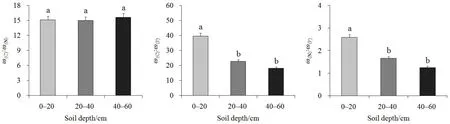
图2 不同土层土壤C、N、P生态化学计量特征的多重比较Fig. 2 Multiple comparisons of C, N, P eco-stoichiometry characteristics at different soil layers
对不同海拔梯度内0—60 cm整体土壤的总C、总N和总P分别进行线性回归分析,结果显示(图3):总 C和总 N之间呈极显著的线性拟合关系(R2=0.791,P<0.001),总N随总C的增加而显著上升;总C和总P、总N和总P之间均存在显著的正向相关关系(R2=0.185,P=0.040;R2=0.181,P=0.043)。这表明秦岭松栎林土壤的总C、总N、总P的空间分布在一定程度上相互耦合,特别是土壤总C和总N之间存在紧密的正向相关性。
3.3 不同土层生态化学计量特征沿海拔梯度的变化
对0—20、20—40、40—60 cm不同土层的C、N、P化学计量沿海拔梯度变化进行单因素方差分析,结果如图4所示。表层土壤C꞉N(F3,22=1.525,P=0.240)、下层土壤 C꞉N(F3,22=1.242,P=0.322)在 4个海拔梯度之间均无显著性差异;中层土壤C꞉N(F3,22=3.147,P=0.049)在不同海拔间存在显著差异,其中,1300—1500 m处的C꞉N显著高于1100—1300 m处,而 1500—1700 m和 1700—1900 m较高海拔处的C꞉N与1100—1300 m、1300—1500 m低海拔处均无显著性差异。0—20 cm土壤 C꞉P(F3,22=1.378,P=0.280)、20—40 cm 土壤C꞉P(F3,22=0.612,P=0.616)、40—60 cm 土壤 C꞉P(F3,22=2.937,P=0.060)在不同海拔梯度间均无显著性差异。表层土壤N꞉P(F3,22=0.761,P=0.530)在不同海拔梯度间无显著性差异。中层土壤 N꞉P(F3,22=4.370,P=0.017)在不同海拔间存在显著性差异,其中,1100—1300 m低海拔处的N꞉P显著高于中、高海拔处,而1300—1500、1500—1700、1700—1900 m等3个海拔段间的N꞉P均无显著差异。下层土壤N꞉P(F3,22=3.416,P=0.038)在不同海拔间也存在显著性差异,其1100—1300 m低海拔以及 1700—1900 m 高海拔处的 N꞉P均显著高于1300—1500 m中海拔处,而1500—1700 m中高海拔与其他3个海拔梯度处的N꞉P之间无显著差异。
4 讨论
4.1 海拔梯度对土壤C、N、P化学计量特征的影响
海拔变化导致的水热条件及植被改变显著影响土壤化学计量特征(Zhao et al.,2016;卢同平等,2017)。C꞉N是土壤质量的敏感性指标,低海拔处温度较高,土壤微生物活性强,其对动植物残体的分解速率增大(刘兴锋等,2019),使得土壤 C꞉N在一定范围内随海拔上升而增加。但伴随海拔升高,混交林中针叶树种的比例逐渐增加,而针叶树凋落物中单宁等酯溶性物质输入土壤的数量也相应增多,抑制微生物活性,导致土壤C累积速率慢于N的累积(李丹维等,2017;徐芷君等,2019);诸多研究还表明,山体垂直带土壤颗粒组成中的粘粒含量一般随海拔上升而减少、砂粒比率增大,易形成土壤团聚体,使得较高海拔土壤有机C的风化发育程度有所降低,且高海拔处坡面径流可携带大量土壤颗粒和有机质运送到中山地带(廖琪等,2017;万丹等,2018),故高海拔土壤的C꞉N呈一定的下降趋势。
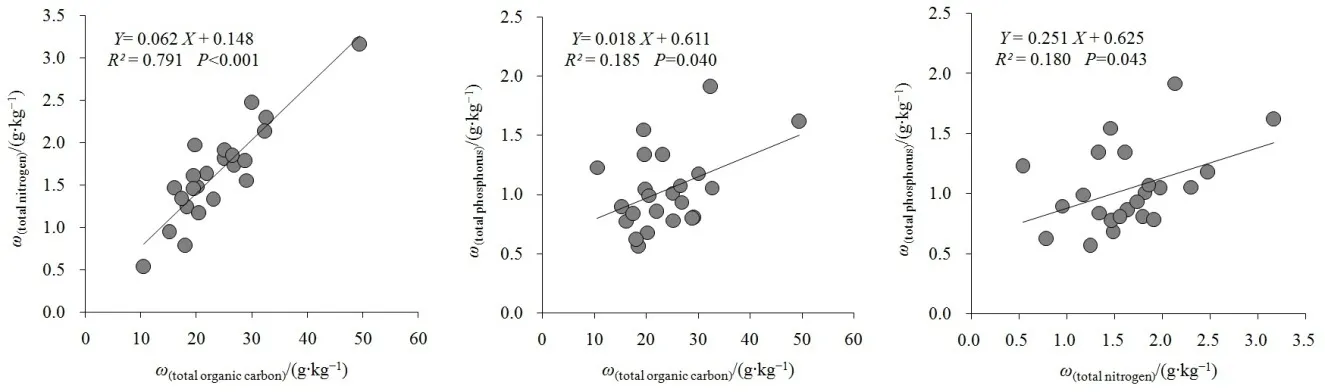
图3 不同海拔土壤总有机碳、总氮、总磷元素的回归拟合关系Fig. 3 Regression relationships between soil C, N, P at different elevations
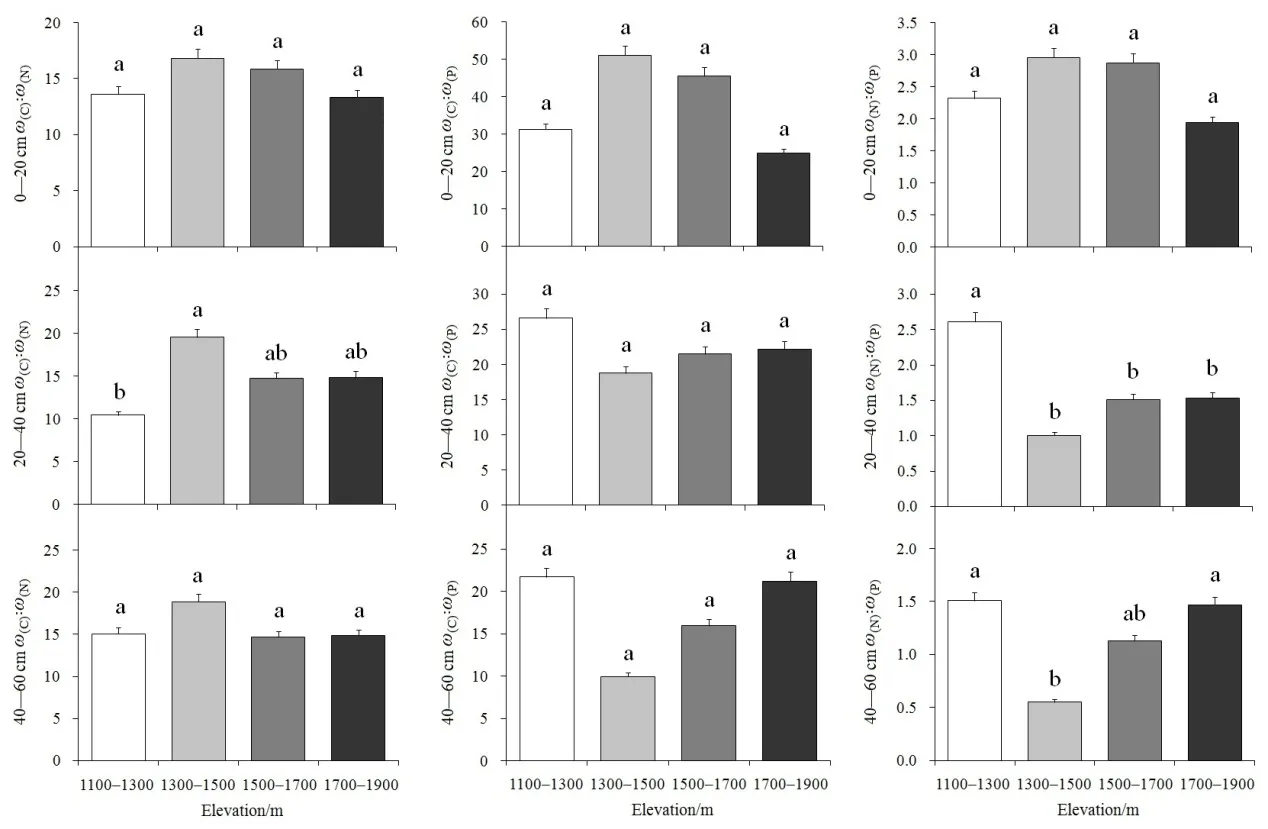
图4 不同土层土壤的C、N、P化学计量特征沿海拔梯度的多重比较Fig. 4 Multiple comparisons of C, N, P eco-stoichiometry along elevations in different soil layers
土壤 C꞉P和 N꞉P反映了土壤养分的供应能力(徐芷君等,2019)。本研究中、高海拔土壤 N꞉P小于低海拔处,可能是由于中、高海拔针叶凋落物较多,其分解释放N的速率相对于阔叶树种较慢,且针叶凋落物分解导致的土壤酸化阻碍了微生物对P的矿化,导致N꞉P下降(Qiao et al.,2014;张雨鉴等,2019)。土壤P主要来自于岩石风化、其空间迁移性较差,且土壤C、P释放不同步(秦海龙等,2019),故C꞉P沿海拔梯度无明显变化。研究区土壤 C꞉N(14.906)略高于全国森林土壤平均水平(13.700),C꞉P(24.081)和 N꞉P(1.657)远小于全国平均水平(C꞉P为44.500,N꞉P为3.200),这有利于土壤微生物的有机磷净矿化,可为植物生长提供充足的磷酸盐(李喜霞等,2018),据此推测研究区土壤的限制性营养元素为N。前人研究还发现海拔升高导致的降水量变化与土壤C꞉P和N꞉P之间具有显著相关性,是由于高海拔区较多的降水会加速P元素的淋溶过程,改变土壤中P的有效性,进而造成微生境内P含量差异性较大(李红林等,2015;李路等,2018),这可能也是本研究林地土壤总P以及N꞉P在高海拔段具有较大变异系数的原因之一。此外,高海拔处森林土壤C꞉N的降低也与高海拔降水增加、温度降低导致的土壤淋溶和水土流失作用的增强有关(Khan et al.,2015;王霖娇等,2018);因此,全球环境变化背景下的气候变暖和降雨量增加可能加剧森林土壤养分流失,未来应更重视高海拔森林土壤的保育工作。
4.2 土层深度对土壤C、N、P化学计量特征的影响
森林土壤C、N主要来源于凋落物分解合成的有机质,表层土壤的枯枝落叶分解较多,腐殖层厚,使得表层土壤比深层土壤更为疏松,具有明显的C、N表层聚集现象,之后其随水分向下层扩散,形成由上至下逐渐减少的分布格局(Jiang et al.,2017;万丹等,2018)。土壤P主要来源于岩石风化,为沉积性元素(李相楹等,2016;汝海丽等,2016),且混交林中松树的根系垂直分布较深,可通过细根周转等方式摄取和改变P的分布,有效减少其沿土壤剖面的渐变趋势(秦海龙等,2019;徐芷君等,2019),故不同土层间P含量比较稳定。此外,秦岭南坡 900—2400 m的山地土壤多为黄棕壤,其铁、铝氧化物含量较高,可促进P的固定作用,且P易与钙质形成难溶性磷酸钙,进一步降低其在土层中的垂直流动性(吴昊,2015 a)。土壤容重随着土层的加深而增加,其紧实度和板结程度加强,植物根系数量减少,导致有机质积累量也随之减少(冯嘉仪等,2018);土层增加也会导致土壤孔隙度减小、通透性变差,使得参与分解有机质的微生物数量和活性随土层加深而降低(邱新彩等,2018),这些原因共同导致研究区表层土壤的 C꞉P和N꞉P显著高于中下层土壤,而20—40 cm、40—60 cm土层各自的C꞉P、N꞉P之间无明显差异。类似的C、N表层聚集现象也发现于广西猫儿山常绿针阔叶混交林和东北红松阔叶混交林土壤中(李喜霞等,2018;秦海龙等,2019)。
研究区土壤总C、总N之间呈极显著的正向耦合关系,而总P变化滞后于二者,与李红林等(2015)关于塔里木盆地绿洲土壤、李丹维等(2017)关于太白山土壤和张雨鉴等(2019)关于滇中亚高山土壤的研究结果相类似。这是由于土壤微生物在利用N时首先需要将有机物的碳骨架破环,使得森林凋落物在分解时同步释放 C、N(Olander et al.,2000),且二者作为结构性元素已经在消耗和积累过程中形成动态平衡(徐芷君等,2019),这种紧密的共变关系也导致C꞉N在不同土层之间无显著变化。研究区 20—40 cm土层中 1300—1500 m处的土壤C꞉N显著高于低海拔段,是由于该处地势较为平缓,山民在此广泛种植中草药猪苓(Polyporus umbellatus),且研究区20—40 cm土层是植物根系最为集中的区域(吴昊等,2012),多种植物对20—40 cm土层N元素的剧烈争夺使得其总N含量锐减,导致1300—1500 m海拔处的中、下层土壤C꞉N上升而N꞉P下降。
综上可知,研究区油松—锐齿槲栎混交林土壤的 C、N、P及其化学计量比在海拔梯度及不同土层中的变异程度较大,应注重对该区土壤的深层施肥,以补充其N、P含量,维持较高水平的土壤肥力;对于海拔1300—1500 m人类活动较为频繁的地段,应尽快合理规划林区内居民生产用地的范围,统筹兼顾,切实维护好林地资源。此外,若在大空间尺度上考虑多地形因子对土壤化学计量的综合效应,则海拔的影响作用将可能被削弱。如:坡度影响水热条件及土壤物质运移堆积,最终影响土壤元素分配(连纲等,2008);坡位影响土壤母岩组成及其砂石比例,可导致上坡位土层表现出明显的养分聚集效应(盛茂银等,2013);坡向制约生境光照和水分状况,导致土壤微生物的生长和繁殖速率发生变化,进而影响土壤养分的转化及归还(Maren et al.,2015)。本研究采用典型取样法,仅探讨了海拔梯度对松栎林土壤的生态效应,而未对样地群落所处的坡度、坡位和坡向等因素进行精确设置和分析,关于多个地形因素对秦岭松栎林土壤的综合效应尚需采用冗余分析(Redundancy Analysis,RDA)等数量生态学技术进一步研究。
5 结论
秦岭油松-锐齿槲栎混交林土壤的 C꞉N、C꞉P、N꞉P平均值分别为14.906、24.081和1.657,变异系数较小。其中,C꞉N略高于全国森林土壤的平均水平,C꞉P和N꞉P均低于全国平均水平,据此推测研究区林地土壤的限制性营养元素为N。松栎林土壤C꞉N、N꞉P随海拔增加分别呈现出较为明显的“先升后降”和“先降后升”变化趋势,C꞉P则无明显变化;0—20 cm土层的C꞉P、N꞉P显著高于20—40 cm和40—60 cm土层,而不同土层间的C꞉N无显著差异;土壤C-N的正向相关性高于C-P和N-P;1300—1500 m海拔处的C꞉N、N꞉P明显异于其他海拔段。全球环境变化背景下,应更加注重高海拔区域松栎林土壤的保育工作,关于研究区“植物—凋落物—土壤”生态系统整体的化学计量特征有待深入探讨。
