季节性冻融期长白山森林溪流中凋落叶N、P的释放动态
2020-01-08陈鸽王璐宫雨薇谭颖张振兴杨海军
陈鸽,王璐,宫雨薇,谭颖,张振兴,杨海军
1. 东北师范大学草地科学研究所/植被生态科学教育部重点实验室,吉林 长春 130024;2. 云南大学高原湖泊生态与治理研究院,云南 昆明 650000
源头溪流不同于陆地生态系统,其郁闭度高,水温低,藻类和水生植被的初级生产力低下,河岸带植被的凋落叶作为外源有机物,成为了源头溪流中主要的能量来源(Vannote et al.,1980;Manuela,2001;Riskin et al.,2017)。在溪流中,凋落叶作为粗颗粒有机物(CPOM)通过快速的淋溶过程、微生物定殖分解、底栖动物摄食分解和物理破碎被分解为细颗粒有机物(FPOM),是河流物质循环和能量流动的关键过程(Wallace et al.,1996;Moore et al.,2004)。其中凋落叶氮(N)、磷(P)元素的释放动态更是土壤-水体营养元素流动的关键纽带(张川等,2015)。
不同凋落叶具有不同的分解速率及元素动态(Djukic et al.,2018),凋落叶质量(结构、营养含量等理化性质)(Peng et al.,2019;Raposeiro et al.,2018;Hector et al.,2000)、环境因子(pH、温度、流速等)(Alida,2018;Martinez et al.,2014)和分解者(底栖动物、微生物)(Hieber et al.,2002)是影响分解过程的 3个关键驱动因素。三者之间的相互作用以及对凋落叶分解和元素动态的影响在空间尺度和时间尺度上具有很大差异(Sariyildiz et al.,2003;Ferronato et al.,2019;Haapala et al.,2001),探究三者间的关系是河流凋落叶分解研究的关键。
北方凋落叶主要在秋季大量输入溪流(张新平等,2008),冬季河床凋落叶一方面能为水生生物提供食物来源(Vannote et al.,1980;Whiles et al.,1997;Lamberti et al.,2000),另一方面能为水生生物提供越冬的栖息地和避难所(Wallace et al.,1999),因此北方溪流冬季凋落叶的分解过程尤为重要。在北方寒冷区,季节性冻融特征显著,季节性冻融过程能通过影响水流、水温、溶解氧、浊度等理化因子直接影响凋落叶的分解(Ruedadelgado et al.,2006;Leroy et al.,2006),并能通过影响微生物和底栖动物的生存及繁殖间接作用于凋落叶的分解过程(Baldy et al.,2007)。
由于冬季溪流冻结,采样不便,关于冻结期凋落叶分解动态的研究匮乏,国外有限的研究集中于冬季河流中凋落叶分解的质量损失动态,及其与底栖动物的定殖关系(Ferreira et al.,2014;Iii et al.,1994),较少有研究关注到了凋落叶分解过程中的元素动态。有限的研究指出了冬季冻结溪流中河床凋落叶仍能维持较高的分解速率,且底栖动物在凋落叶分解过程中起到了较大的作用(Boyero et al.,2011)。国内相关研究仅见在岷江开展的关于高山森林冬季溪流凋落叶分解过程质量变化及分解过程中N、P等养分元素动态特征的研究(岳楷等,2015;张川等,2016),以及针对长白山地区溪流冬季冻结初期凋落叶分解与底栖动物定殖关系的研究(王璐等,2018)。尚未见关于季节性冻融期凋落叶元素动态与底栖动物作用的研究。
本研究旨在通过对冬季长白山森林源头溪流内两种不同质量植物色木槭(Acer mono)和蒙古栎(Quecus mongolica)凋落叶分解过程的研究,揭示季节性冻融过程凋落叶的分解动态及元素释放动态。基于前期研究,我们提出的科学假设如下:(1)冬季长白山源头溪流凋落叶仍能分解,色木槭凋落叶分解速率显著高于蒙古栎;(2)冬季长白山源头溪流凋落叶分解过程中不同凋落叶的N、P元素释放动态存在差异;(3)底栖动物能显著促进凋落叶分解及元素释放过程。
1 研究地概况
试验溪流位于吉林省通化市辉南县龙湾国家级 自 然 保 护 区 内 ( 42°19′22″— 42°19′43″N ,126°28′20″—126°30′7″E),地处长白山北麓龙岗山脉中段。该地区属于北温带大陆性季风气候,历年年平均降水量约为704.2 mm,降水多集中在夏季。历年年平均气温约4 ℃,最低月均温为-18 ℃,出现在1月,最高月均温为22.4 ℃,出现在7月(杨一,2017)。该地区冬季漫长寒冷,每年11月至次年4月为溪流结冰期,约70%的水面被冰层覆盖。年水温0—15 ℃,冬季冰面下水体仍维持流动性,冰下水体温度约为0.5 ℃。研究地流域森林覆盖率大于75%,为地带性红松针阔叶混交林,河岸带以色木槭(Acer mono)、蒙古栎(Quecus mongolica)、山杨(Populus davidiana)、紫椴(Tilia amurensis)等为主要优势阔叶树种。试验断面位于保护区溪流源头,无人为干扰和旅游开发,能较好地反映自然条件下凋落叶的分解动态。
2 材料与方法
2.1 试验设计及样品处理
基于前期调查及文献调研,选择当地河岸带最优势植物色木槭和蒙古栎为研究对象,其中色木槭分解速率相对较高一般属于中、快速分解组,而蒙古栎分解较慢属慢速分解组(Canhoto et al.,1996;Eggert et al.,2003;刘昕等,2008),选择这两种分解速率不同的凋落叶,进而研究凋落叶自身性质对分解及元素动态的影响。供试凋落叶收集于2017年9月末,在试验地河岸带,收集新鲜老熟落叶,选取叶片完整、无明显破损和菌食的叶片带回实验室。蒸馏水清洗除掉表明杂质后,于通风干燥处风干,2种凋落叶分别准确称取6 g的风干样各5袋,置于65 ℃鼓风干燥箱中烘干至恒质量,由此确定样品的含水率,计算其初始质量。为了比较分解袋中底栖动物的存在对凋落叶分解是否具有影响,本实验设计了2种孔径的尼龙凋落叶分解袋(15 cm×20 cm):一种是允许底栖动物自由出入,孔径为5 mm的网袋;另一种为底栖动物不易进入,仅允许微生物定殖的孔径为0.3 mm的网袋。称取(6±0.01) g凋落叶分别封入两种孔径的分解袋内,2种凋落叶用粗细分解袋各装15袋,共60袋。4个凋落叶分解袋为一组(2种凋落叶×2种孔径),随机依次缝于安全绳上,相邻凋落叶分解袋间隔不小于0.4 m,共15组。
于2017年10月7日,在试验河段内选择生境条件较一致的 10个缓流浅滩生境,间距不小于 20 m,进行凋落叶分解袋的投放。每个缓流浅滩生境顺水流方向放置一组或平行放置两组,两组分解袋间距不小于0.4 m,将缝有凋落叶分解袋的安全绳顺水流方向放置于水体中,并用钢筋将两端固定于河底。
样品放置后,分别在第46天(2017年11月21日,冻结初期)、97天(2018年1月11日,冻结期)、170天(2018年3月25日,融化期)取样,每次从10个样点取样随机取出5组凋落叶分解袋,取出后迅速分装于塑料封口袋中,放入装有冰袋的保温箱中低温保存,带回实验室。将样品袋放在500 μm网筛上小心冲洗,洗去沉积物。洗净凋落叶上的泥沙和杂物后,分别封入纸袋,置于鼓风干燥箱,65 ℃烘干至恒量后称取其干质量并测定养分质量分数。全氮及全磷用硫酸高氯酸消煮后使用Smartchem450全自动化学间断分析仪测定,每个样品重复测定3次。
2.2 水理化指标测定
在凋落叶分解袋布设和收集的同时,进行水环境指标的测定。使用米尺测定河宽,水深探测棒测定水深,便携式流速分析仪(欧瑞卡LS300A)测定断面0.6倍水深处的流速,使用便携式水质分析仪(哈纳HI9819T)测定pH值、溶解氧、电导率等水质指标,每个指标重复测 3次。为了表征整个分解过程中凋落叶分解袋内的温度变化,随机选择 2个分解袋,各放入1个纽扣式温度计同步记录温度的变化。
2.3 数据处理与分析
采用凋落叶质量剩余率表示凋落叶的质量损失情况,质量剩余率为分解过程中凋落叶的剩余质量占初始质量的百分率。凋落叶分解曲线的模拟依据 Olson 指数衰减模型(Olson,1963):mt=m0×e-kt,其中t为分解时间(d),mt为时间t时凋落叶剩余的干质量(g),m0为凋落叶的初始干质量(g),k为凋落叶质量损失的分解速率常数,单位为d-1。凋落叶失质量速率V0(%)=(mt-1-mt)/D△t,(mt-1-mt)为相邻采样时间凋落叶分解袋内凋落叶质量残留量之差(t=1, 2, 3),D△t为相邻采样时间间隔天数。凋落叶养分元素的 释 放 率E=[(mt-1×Ct-1-mt×Ct)/(m0×C0)]×100%,式中,m0为放置凋落叶袋时袋内凋落叶的干质量(g),mt为第t次采样时凋落叶袋内凋落叶的干质量(g)(t=1, 2, 3),C0为初始N、P养分质量分数(mg·g-1),Ct为第t次采样时期 N、P养分质量分数(mg·g-1),当E>0,表示该元素出现释放,E<0时,表示该元素出现富集。
数据统计与分析采用SPSS 21.0和Origin 9.1软件完成。采用单因素方差分析(One-way ANOVA)和最小显著差异法(LSD)分析冬季不同时期凋落叶质量剩余率及凋落叶N、P质量分数。采用t检验分析各时期底栖动物对凋落叶质量损失及 N、P释放动态的影响。
3 结果
3.1 溪流特征
2017年10月—2018年3月,记录研究地冻结初期、冻结期、融化期3个时期的溪流特征,如表1所示。表1显示,研究地溪流温度随着冻结期的到来逐渐降低,融化期重新上升。流速在冻结期最缓,随着冰雪消融流速越来越高。水深在冻结期急剧下降,融化期雪水造成水位迅速回升。温度的高低、流速的变化及水位的明显涨落可能会对凋落叶分解速率造成影响。

表1 研究地溪流特征Table 1 Stream characteristics of the study site
3.2 凋落叶分解动态
根据凋落叶指数衰减模型拟合得到凋落叶的分解速率(表2),粗细凋落叶分解袋中蒙古栎的分解速率均显著低于色木槭(P<0.05),同树种凋落叶粗细分解袋间对比结果显示,色木槭在粗凋落叶分解袋中的凋落叶的分解速率显著高于细凋落叶分解袋(P<0.05)。

表2 两种树叶在不同分解袋类型中的分解速率Table 2 Decomposition rates of two species of leaves in two types of litterbags
凋落叶分解动态如图 1所示。经过一个冬季(170 d)的分解,粗分解袋中的蒙古栎和色木槭分别完成了63.74%和90.24%的分解,细分解袋中的蒙古栎和色木槭分别完成了45.28%和62.92%的分解。冻结初期,2种凋落叶在粗细分解袋内的质量损失均无显著差异。冻结期,色木槭凋落叶在粗分解袋内的质量损失显著高于细分解袋(P<0.05)。融化期,蒙古栎及色木槭凋落叶在粗分解袋内的质量损失均显著高于细分解袋(P<0.05)。
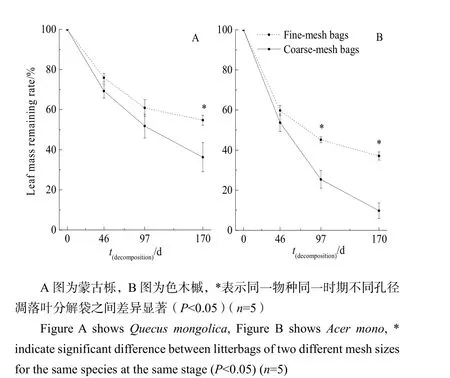
图1 分解过程中蒙古栎和色木槭凋落叶质量剩余率Fig. 1 Mass remaining rate of Quecus mongolica and Acer mono during leaf litter decomposing
整个冬季,凋落叶在不同时期的失质量速率表现出较一致的规律(表3)。均显著表现为冻结初期>冻结期>融化期(P<0.05)。此外,色木槭凋落叶在冻结初期的分解速率显著高于蒙古栎(P<0.05),而冻结期仅粗分解袋中色木槭凋落叶的分解速率显著高于蒙古栎(P<0.05)。同树种凋落叶粗细分解袋间对比结果显示,冻结期粗分解袋中的色木槭凋落叶失质量速率显著低于细分解袋(P<0.05)。

表3 两种凋落叶的失质量速率Table 3 Mass loss rate of two leaf litters %

图2 蒙古栎和色木槭凋落叶N和P质量分数动态Fig. 2 Dynamics of N and P mass fraction of Quecus mongolica and Acer mono leaf litter
3.3 凋落叶养分元素质量分数动态
图2为蒙古栎和色木槭凋落叶N、P质量分数随时间变化的情况。对于蒙古栎,在冻结初期粗细分解袋内的凋落叶都经历了 N质量分数的迅速增加,但两凋落叶袋间没有显著性差异。随后,粗分解袋内N质量分数开始下降。在冻结期,粗分解袋内凋落叶N质量分数显著低于细分解袋(P<0.05)。而细分解袋内N质量分数的上升时间更长,直到融化期才开始下降,但170 d时仍比初始质量分数增加了64.41%。色木槭凋落叶N质量分数随时间变化的规律与蒙古栎凋落叶相似,除了冻结期,融化期粗分解袋内凋落叶 N质量分数也显著低于细分解袋。
凋落叶 P质量分数在冻结初期总体呈上升趋势。蒙古栎凋落叶在粗分解袋内于冻结初期达到最高质量分数,随后开始下降,最终接近初始值水平;而在细分解袋内,P质量分数持续上升,冻结期达到最高值后才开始下降,并在170 d时接近初始质量分数。色木槭凋落叶在粗细分解袋内的质量分数变化规律基本一致,冻结初期后,在冻结期持续上升并达到最高值,于融化期开始下降,170 d时与初始值相比粗分解袋中P质量分数增加了9.20%,细分解袋中P质量分数增加了19.67%。
3.4 凋落叶养分元素释放(富集)率动态
由图3可见,粗细分解袋中蒙古栎凋落叶的N元素均在冻结初期表现为微量富集现象,在冻结期及融化期均释放,呈现出富集-释放-释放的模式。粗细分解袋中色木槭凋落叶的N元素表现为明显的释放现象。整个季节性冻融期间,粗细分解袋内两种凋落叶的P元素均表现为明显的释放现象。
整体分解过程中,冻结期两种凋落叶N、P元素的释放率显著高于其它两个时期(P<0.05),其中色木槭N元素在此时期的释放率高达66.07%,P元素释放率高达46.77%。蒙古栎凋落叶的N释放(富集)率在粗细分解袋中无显著差异,粗分解袋中的P释放率在冻结期显著高于细分解袋(P<0.05)。色木槭凋落叶的N、P释放(富集)率在冻结初期的粗细分解袋中均无显著差异,冻结期粗分解袋内凋落叶N、P释放率显著高于细分解袋(P<0.05),而融化期细分解袋内凋落叶N、P释放率显著高于粗分解袋(P<0.05)。
4 讨论
4.1 凋落叶分解动态
大量研究表明,凋落叶的质量是影响凋落叶分解的关键因子(Konig et al.,2014;Leroy et al.,2006),低韧性的凋落叶往往分解速率较高(Graca,2015;Mathuriau et al.,2002)。本研究也证明了这一点,根据Petersen et al.(1974)的划分标准,粗凋落叶分解袋中的色木槭树叶属于快速分解组,蒙古栎树叶属于中等组;细凋落叶分解袋中色木槭属于中速分解组,而蒙古栎树叶属于慢速分解组(表2)。两种分解袋中,质量较高的色木槭凋落叶的分解速率均显著高于蒙古栎凋落叶(表3),同时由于本地生物长期以2种优势树种凋落叶为食,所以2种凋落叶仍维持了相对较高的分解速率,符合假设一“冬季长白山源头溪流凋落叶仍能分解,色木槭凋落叶分解速率显著高于蒙古栎”。

图3 蒙古栎和色木槭凋落叶N和P释放率动态Fig. 3 Dynamics of N and P release rate in decomposing leaf litter of Quecus mongolica and Acer mono
凋落叶在溪流中的分解过程整体来讲一般分为3个阶段(Petersen et al.,1974):淋溶、微生物定殖、底栖动物定殖及物理破碎。淋溶作用在分解初期占主导地位,通常在短期内导致凋落叶质量的大量损失(Menendez et al.,2003)。本研究中,初始的46 d(冻结初期)凋落叶分解速率最高,达到15.71%—30.21%(表3)。随着分解的进行,相对于冻结初期,冻结期及融化期凋落叶的分解速率降低很多。这主要是因为冻结初期通过淋溶作用,一些未完全分解的有机组分从凋落叶中大量释放(Webster et al.,1986),导致凋落叶质量损失明显。但由于淋溶作用主要发生在凋落叶浸入水中后的短时间内(Shieh et al.,2008),冻结期及融化期淋溶作用不再是凋落叶分解的主要影响因素(张川等,2016),微生物及底栖动物的缓慢分解占据了质量损失的主要地位,因此冻结期和融化期凋落叶的质量损失小于冻结初期。
溪流中底栖动物的定殖和取食活动会引起凋落叶的机械破碎,导致凋落叶更快的分解(Bird et al.,1992;Xu et al.,2005)。本研究中,经过整个冬季过程后,粗分解袋内2种凋落叶的质量剩余率显著(P<0.05)低于细分解袋(图1),说明虽然冬季水温低,底栖动物在寒冷条件下仍能维持较高的活性。冻结初期,粗细凋落叶分解袋内质量剩余率差异不显著,表明分解初期底栖动物对凋落叶质量损失的贡献率较低,而微生物是主要的分解者。大量研究也表明,底栖动物往往倾向于取食被微生物分解定殖过的凋落叶(Frainer et al.,2015;Gessner et al.,1994)。进入冻结期,底栖动物对色木槭凋落叶的分解作用更显著(表3),这可能由于分解蒙古栎凋落叶的主要分解者仍是微生物。融化期结束后,底栖动物对色木槭分解的贡献为27.32%,对蒙古栎凋落叶分解的贡献仅为18.46%。这表明了底栖动物对食物具有选择性,其更倾向于摄取质量较好、可快速分解的色木槭凋落叶,并且对于色木槭而言,冻结期是底栖动物作用的最主要时期,符合假设三“底栖动物能显著促进凋落叶分解过程”。
4.2 凋落叶养分元素释放动态
在170 d的分解过程中,蒙古栎凋落叶的N元素表现出富集-释放-释放的模式(图3),与Shieh et al.(2008)的研究一致,而色木槭凋落叶的N元素在各个时期均表现为释放现象,这说明N的释放动态在不同凋落叶中具有不同的释放规律,胡霞等人的研究也表明凋落叶 N元素的动态变化是无规律的(胡霞等,2012)。整体季节性冻融期分解过程中,2种凋落叶的P元素在各个时期均呈现出明显的释放模式,与Peng et al.(2018)及Elwood et al.(1981)研究的结果不一致,这可能是由于 P元素的释放在不同时期、不同地点具有不同的动态。本研究中两种凋落叶的N元素动态有所差异,但P元素的释放动态基本一致,仅在一定程度上符合假设二“冬季长白山源头溪流凋落叶分解过程中不同凋落叶的N、P元素释放动态存在差异”。
在长白山源头溪流冻融期的分解过程中,底栖动物对凋落叶 N质量分数的影响较 P质量分数更为显著(图2)。冻结期底栖动物的存在显著降低了2种凋落叶的N质量分数(P<0.05)。同时,冬季分解过程中底栖动物对溪流中凋落叶的P质量分数没有显著影响,说明了底栖动物与凋落叶P质量分数的变化无显著关联。而凋落叶元素释放(富集)率的差异可能是由于基质质量导致的(岳楷等,2015)。冻结初期,河岸带植被叶片大量脱落,是凋落叶输入溪流的高峰期(Leff et al.,1990),淋溶作用会导致 N、P元素从凋落叶中大量释放(Qiu et al.,2002),淋溶后微生物迅速定殖,并在二次生产中加入营养物质加速叶片破碎(Graca,2015;Pozo et al.,1998),这可能是底栖动物在此时期对凋落叶的元素动态影响不显著的原因。随着分解的进行温度逐渐下降,冻结期溪流表面部分冻结,在冰层覆盖的情况下,由于流速减缓,同时低温阻止了融雪径流,物理磨损达到最低(Bird et al.,1992)。虽然进入冬季温度降低,但溪流水温仍能维持在0 ℃以上(表 1)且温差波动小,相对稳定的环境为微生物及底栖动物的活性提供了条件,分解者在此时期仍能对凋落叶进行缓慢的分解(Polunin,1984)。冻结期底栖动物作用导致2种凋落叶P元素及色木槭N元素呈现更显著的释放现象(P<0.05),这说明冻结期是底栖动物影响元素释放动态的关键时期。Iii et al.(1994)的研究也指出低温下微生物活性较低,微生物介导的凋落叶分解率会随纬度的增加而降低,在高纬度上底栖动物更为重要,这在一定的程度上也支持了我们的研究。融化期温度回升,溪流冻结部分融化及融雪径流导致凋落叶分解及营养元素浸出(Pettit et al.,2012),考虑到温度上升,微生物的活性及作用也可能相应的增强,同时由于细分解袋中凋落叶的质量高于粗分解袋,因此研究结果显示底栖动物在此时期对凋落叶N、P释放的影响较低。以上符合假设三“底栖动物能显著促进凋落叶元素释放过程”。
5 结论
为探究冻融期凋落叶的元素释放动态及底栖动物的贡献率,我们在长白山森林源头溪流用色木槭、蒙古栎凋落叶进行了试验研究。研究发现,在冻融期,不同凋落叶的分解速率不同,不同凋落叶的元素释放动态存在一定差异,N、P元素整体表现为释放现象,底栖动物对凋落叶的分解及元素释放动态起到了显著的作用。因此河岸带植被多样性的保护是具有重要意义的,维持河流底栖动物的多样性也是河流保护的关键。这些结果为深入了解长白山源头溪流底栖动物及物质循环过程的关系提供了一定的基础数据。
