氢过氧化物氧化对核桃蛋白结构和乳化特性的影响
2020-01-02王丹丹毛晓英孙领鸽吴庆智李宝坤程卫东
王丹丹 毛晓英 孙领鸽 吴庆智 李宝坤 程卫东
(石河子大学食品学院 新疆石河子 832003)
脂肪氧化酶(LOX)普遍存在于植物性食品种子中[1]。植物中的LOX可能与脂肪酸过氧化,脂质贮藏,生长调节因子的产生有关[2]。不饱和脂肪酸氧化酸败导致核桃蛋白发生氧化酸败,食品色泽、风味及品质的改变,降低核桃蛋白的营养价值[3]。由于LOX在可控条件下催化亚油酸形成的13-氢过氧化-顺-9,反-11-十八碳二烯酸(HPODE)结构不稳定,在热、紫外线以及金属离子存在的情况下易于分解[4],所以有关HPODE氧化蛋白质的研究较少。Kong等[5]报道,大豆蛋白的氧化程度与脂肪氧合酶的活性有关;Huang等[6]提出大豆蛋白的结构经LOX催化亚油酸反应的产物氧化后发生改变。蛋白质具有良好的两亲性,其可以结合在油水界面上,减少油水界面的表面张力,基于这一性质,蛋白质可以在油水界面上形成乳化液(O/W、W/O)[7]。蛋白质的乳液性质与结构性质有关,氧化引起的结构改变,可能影响乳化性质的改变[8]。Dickinson[9]研究乳化液,发现酪蛋白可在分散的液滴周围形成黏弹性膜,以减少在油水界面的表面张力,这可以保持乳液的稳定性;Wang等[8]研究酪蛋白氧化,发现蛋白质氧化是导致乳液性质恶化的关键因素。
对于LOX-亚油酸-核桃蛋白模拟氧化体系的研究尚未见报道。本研究以LOX催化亚油酸形成的HPODE代表LOX诱导脂质过氧化反应中形成的脂质氢过氧化物。以HPODE氧化的核桃蛋白为研究对象,利用HPODE对核桃分离蛋白(WPI)进行不同程度的氧化修饰,通过分析WPI的氧化结构指标(如羰基含量、表面疏水性、粒径分布和分子质量大小)以及乳化性质(如乳液活性及其稳定性、乳液粒度、乳液荧光)的变化规律,探讨不同亚油酸添加量所致蛋白质氧化对核桃蛋白质结构和乳化特性的影响,为进一步阐明核桃蛋白氧化机理提供理论依据。
1 材料和方法
1.1 材料与试剂
新疆薄皮核桃,购于新疆石河子市农贸市场;脂肪氧合酶(LOX I-B)(98 005 units/mg),东京化成工业株式会社,日本;亚油酸,色谱纯;2,4-二硝基苯肼,1-苯氨基萘-8-磺酸等分析试剂均为分析纯。
1.2 仪器与设备
LGJ-18S冷冻干燥机,北京松源华兴科技有限公司;荧光分光光度计,日立F-7000;EX30生物显微镜,武汉华科达实验设备有限公司。
1.3 方法
1.3.1 WPI的制备 参照毛晓英的方法[10],选取新疆当年产薄皮核桃,将核桃仁在2%碱液中浸泡4 min,后手动去皮、粉碎,以正己烷为溶剂脱脂,制备核桃脱脂粉。核桃脱脂粉醇洗后,溶于去离子水中,碱溶(pH 11)、酸沉(pH 4.5)法得到核桃分离蛋白,用去离子水调节沉淀pH 7.0,冷冻干燥得到核桃分离蛋白,至于4℃冰箱备用。
1.3.2 HPODE氧化蛋白的制备 参照叶林的方法略改[11]。配制底物亚油酸溶液:1.5 g亚油酸、1 mL 5 mmol/L NaOH溶液,2滴Tween 20摇匀混合,用0.2 mol/L pH 9.0的硼酸缓冲液于50 mL容量瓶中定容。
配制酶液:将90 mg LOX溶于50 mL pH 9.0的磷酸盐缓冲液。
模拟反应种类:WPI溶于去离子水,碱液调节pH 9.0。称取100 g溶液于5个锥形瓶中。在5个加入10 mL酶液的100 g溶液的锥形瓶中,加入亚油酸量为 0,3,4.5,7.5,9 mL,振荡摇匀,4℃冰箱培养6 h后取出,以酸液调节 pH至4.5,离心(5 000 g×30 min),蛋白沉淀分散于去离子水中,调节至 pH 7.0,离心(8 000 g×30 min),除去不溶物,将其分别冷冻干燥,得到氧化核桃分离蛋白。至于4℃冰箱备用。
1.3.3 WPI乳状液制备 上述核桃蛋白为原料制备乳状液。1%的蛋白溶液,与体积分数10%的大豆油,高压均质后得到所需乳状液。
1.3.4 结构测定
1.3.4.1 羰基含量的测定 参照Huang等[12]的方法进行测定。将蛋白用去离子水配制为5 mg/mL,用双缩脲法测定上清液中的蛋白质含量。在367nm 处用2,4-二硝基苯肼比色,用22000 M-1cm-1消光系数计算每毫克蛋白质羰基衍生物的物质的量。
1.3.4.2 游离巯基和总巯基含量的测定 根据Huang[12]的方法略改,采用DNTB比色法。核桃蛋白样品溶于0.1 mol/L含有1 mmol/L EDTA和1%SDS pH 8.0的磷酸盐缓冲液中,测定上清液中蛋白浓度。取上清液在412 nm波长下测定吸光度,以13 600 M-1cm-1消光系数计算巯基和总巯基含量。
1.3.4.3 表面疏水性的测定 采用ANS荧光探针法[12]。样品溶解于0.01 mol/L pH 8.0磷酸盐缓冲液中,BCA 法测定上清液中蛋白质量浓度为0.005~0.5 mg/mL之间,加入ANS探针,在激发波长和发射波长分别为395,473 nm下测定荧光强度。荧光强度值为Y轴,蛋白质浓度为X轴作图,蛋白质浓度为0,Y轴得值即为蛋白质表面疏水性指数。
1.3.4.4 粒径分布测定 根据Huang的方法略改[12]。样品溶解于0.01 mol/L pH 7.0磷酸盐缓冲液中,BCA法测定上清液中蛋白质量浓度为1mg/mL,采用纳米粒度分析仪测定核桃蛋白粒径分布。
1.3.5 乳化性质
1.3.5.1 乳液活性及其稳定性 乳化活性(EAI)和乳化稳定性(ESI)方法参考文献[13]。将蛋白样品溶于去离子水中,取9 mL蛋白溶液,加3 mL大豆油,于10 000 r/min均质2 min,立即移取50 μL于5 mL 1%SDS里面,在500 nm下测A0,此时为EAI。10 min 后,再测 A10,此时为 ESI。分别代入以下公式,计算。
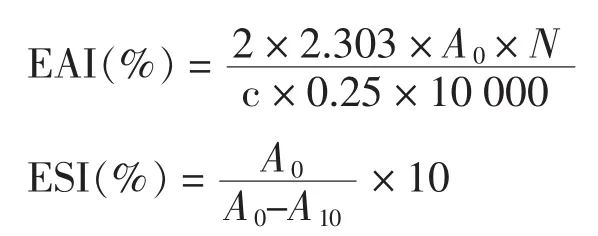
1.3.5.2 乳液内源荧光测定 取乳液样品120 μL用6 mL 0.01 mol/L磷酸盐缓冲溶液(pH 7.0)稀释,混合均匀后在激发波长为290 nm处,扫描300~400 nm发散谱带,狭缝宽均为5 nm。
1.3.5.3 乳状液乳液粒度的测定 乳状液用去离子水稀释至1∶1 000(w/w)粒度分布仪测定乳液的粒子大小及其分布。
d3,2:表面积平均直径(surface area mean diameter)
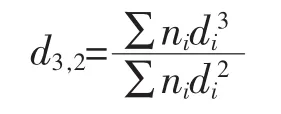
d4,3:体积平均直径(volume mean diameter)
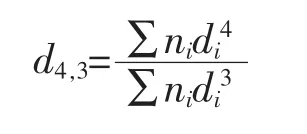
式中ni——直径(μm);di——脂肪球的数量。
1.3.5.4 乳液微观结构 使用光学显微镜(EX30)观察乳液的微观结构,每个样品移取 10 μL制成玻片标本,用 400×的物镜进行观察,根据照片记录进行分析。
1.4 数据处理与统计
每个试验重复3次,结果表示为平均数±SD。显著性分析采用SPSS statisics 17.0分析,数据绘图使用Origin 8.5。
2 结果与讨论
2.1 氧化对结构指标的影响
2.1.1 核桃蛋白羰基含量分析 蛋白质羰基含量是评价蛋白质氧化的最广泛使用的方法之一[14]。在本文中,氢过氧化物引起的氧化用羰基群的形成来表示。结果如表1所示,在LOX催化亚油酸氧化系统中,亚油酸添加量增加到9 mL时,WPI样品羰基从未氧化的2.32 nmol/mg增加到4.23 nmol/mg(P<0.05)。羰基群的形成可以导致蛋白质分子间的共价结合,赖氨酸残基氧化,形成醛类,形成的醛类进一步相互作用,可与游离氨基酸群结合或者在两个羰基群之间发生醇醛缩合[15]。蛋白羰基的形成,表明氧化可引起单个氨基酸残基转化[16],多肽骨架断裂或与非蛋白羰基化合物结合。Huang等[17]研究LOX催化亚油酸对大豆蛋白氧化时发现,LOX诱导的脂质过氧化,较高浓度氧化时,大豆蛋白羰基含量显著增加,增加的羰基含量,暗示着氨基酸组分的变化,将会对大豆蛋白结构改变产生影响。
2.1.2 核桃蛋白巯基值和总巯基含量分析 巯基(SH)的含量是蛋白质氧化的另一个普遍的指示器[14]。不同氧化条件,巯基可以被氧化成可逆(二硫化合物和次磺酸)和不可逆(亚磺酸和磺酸)的两种形式[18]。蛋白质氧化,SH群丢失是最早可观测到的指标,这主要归因于蛋白质内部多肽与多肽之间形成二硫键[19]。不同亚油酸添加量下,WPI游离巯基和总巯基含量如表1所示,底物亚油酸含量增加,WPI的游离巯基含量下降,从2.72 nmol/mg 下降到 1.92 nmol/mg(P<0.05)。半胱氨酸残基是蛋白质氧化最敏感的氨基酸残基[14],其和二硫键对蛋白质结构起到重要的影响[20]。由此可得,蛋白质氧化,可以改变蛋白质的结构特性。Wu等[21]在研究脂质过氧化产物对大豆蛋白氧化机理时指出,当添加HPODE为较高氧化浓度时会导致巯基和总巯基含量显著下降。本研究中,HPODE添加量在低氧化浓度时,巯基和总巯基含量都表现显著下降(P<0.05),此结果可能与原材料不同有关。
2.1.3 核桃蛋白表面疏水性分析 氧化修饰蛋白质表面的氨基酸残基可以对蛋白质的构象和空间结构产生明显的影响,表面疏水性或暴露的疏水群可以表示蛋白质的结构特征[19]。其可以评价蛋白质构象转变[8]。从表1得出,亚油酸添加量达到9 mL时,WPI表面疏水性指数与未氧化的WPI相比下降了5.68%。可见亚油酸氧化处理后,H0下降。这可能是因为亚基之间相互聚合,或者蛋白质分子在疏水作用和二硫键相互作用下重新聚集,暴露的疏水残基共价修饰、引入新的亲水组分[22],或者是由于氧化导致蛋白结构发生解开再聚集的修饰,导致原先暴露的疏水基团更大的被包埋,并且形成新的亲水组分 (例如羰基群)[8]。黄友如在LOX-亚油酸-大豆蛋白模拟体系中发现随着大豆蛋白氧化程度的增加,表面疏水性逐渐下降[23]。
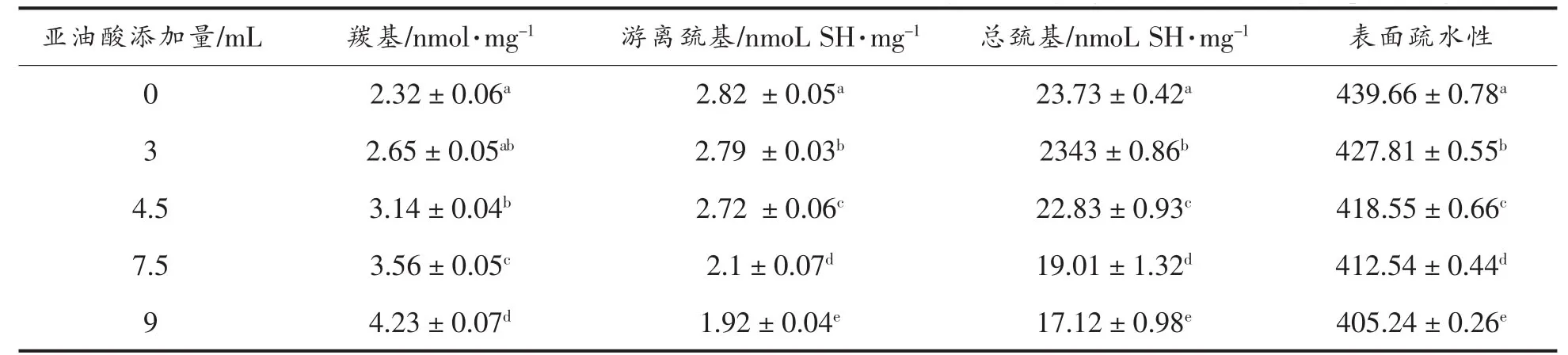
表1 不同亚油酸添加量对WPI羰基、巯基及表面疏水性的影响Table1 Effects of different amount of linoleic acid added on the carbonyl,sulfhydryl and surface hydrophobicity of WPI
2.1.4 核桃蛋白粒径分布分析 不同亚油酸添加量下,核桃分离蛋白粒径分布如图1所示。与没有氧化的样品相比,亚油酸添加量为3 mL时,WPI的粒径分布逐渐减小。当底物亚油酸添加量为9 mL时,WPI的粒度达到最大。粒子尺寸的改变反应蛋白结构的改变。基于图1得到的数据,粒径分布趋势如此可能是由于低浓度氧化引起WPI的分子结构展开,导致平均粒径变小,这可能是由于一些较大的可溶性聚合物分解成更小的可溶性多肽。氧化程度增加,暴露的疏水基团相互作用,生成可溶性聚集体(这与上述疏水性下降一致)。或者是由于二硫键的共价结合,引起WPI的共价聚集(这与游离的SH群减少,表明氧化蛋白质的ss形成保持一致),转变成不可溶性聚集体,致使平均粒径变大。
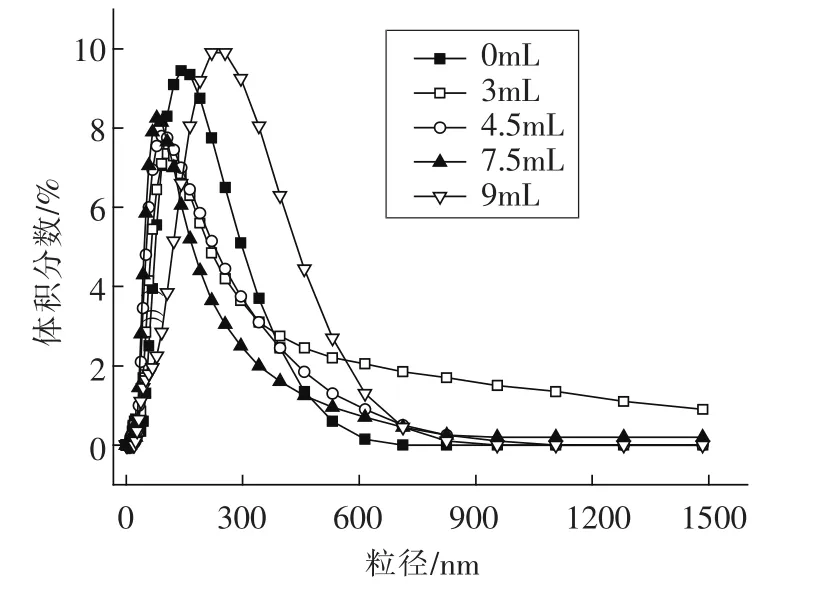
图1 不同亚油酸添加量对WPI粒径分布的影响Fig.1 Effects of different amount of linoleic acid added on the particle size distribution of WPI
2.2 氧化对乳化性的影响
2.2.1 核桃蛋白乳状液活性及其稳定性分析 不同亚油酸添加量对WPI乳化活性及乳化稳定性的影响如图2所示。未氧化的WPI的乳化活性指数(EAI)和乳化稳定指数(ESI)分别为 74.66 m2/g和 20.9%。当亚油酸添加量为4.5 mL时,两项指标均得到改善。此时,EAI和ESI分别增加到88.98 m2/g(P<0.05)和27.49%。亚油酸添加量继续增加,EAI呈现明显降低的趋势,而ESI降低不明显。产生这种结果可能是由于巯基群氧化所形成的二硫键,致使蛋白质分子之间共价交联,形成的大分子聚集体可以在油滴表面形成更稳定的蛋白膜[15],所以适度氧化蛋白质ESI有所增加。
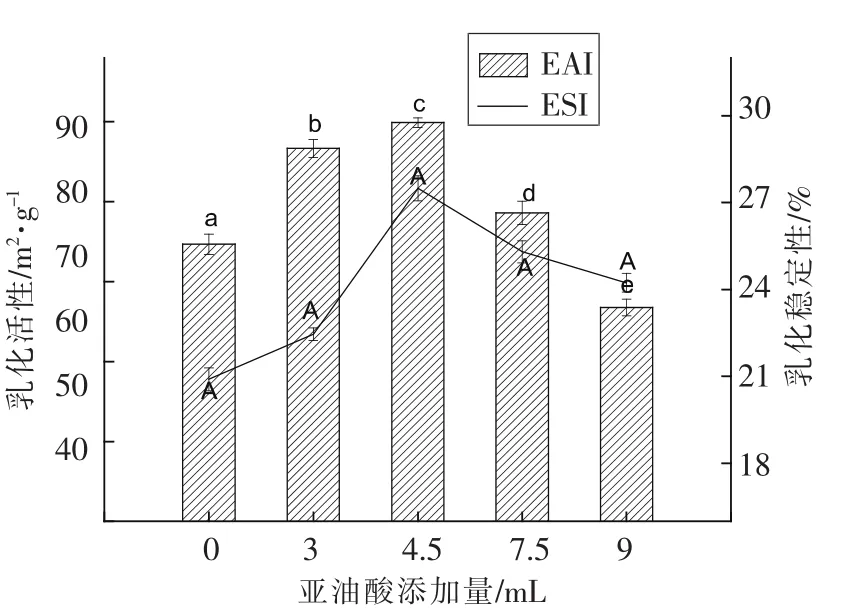
图2 不同亚油酸添加量对WPI乳化活性及乳化稳定性的影响Fig.2 Effects of different amount of linoleic acid added on EAI and ESI of WPI emulsion
2.2.2 核桃蛋白乳状液内源荧光分析 色氨酸残基可以在激发波长290 nm处,于300~400 nm范围内发出荧光[24],荧光光谱分析主要由色氨酸(Trp)残基的极性环境决定,其可监测蛋白-蛋白、脂质-蛋白的构象改变[25]。新鲜核桃蛋白乳液在290 nm处激发得到色氨酸为发射基团的荧光光谱,可以反映乳液中色氨酸残基的氧化程度及其微环境的变化。荧光强度显著降低,指示着蛋白结构逐渐展开[26]。表2所示,未氧化的WPI荧光强度为695.3,当亚油酸添加量为9 mL时,荧光强度下降到664.5。此结果表明,氢过氧自由基氧化破坏了乳状液的结构。Estevez[27]曾报道,氧化降低了肌原纤维蛋白O/W型乳液色氨酸的荧光强度。发色团更多的暴露在极性环境下,最大内源荧光发射光谱红移,发色团与猝火剂相互作用,溶剂和蛋白质荧光的量子产量下降[28]。然而,最大内源荧光(λmax)从 335 移动到 330(蓝移),说明亚油酸添加量增加,乳液中色氨酸转移到更加疏水的非极性环境中[29]。这可能是由于氧化的WPI蛋白中,色氨酸残基被埋藏在疏水基团内部,导致了λmax的下降。
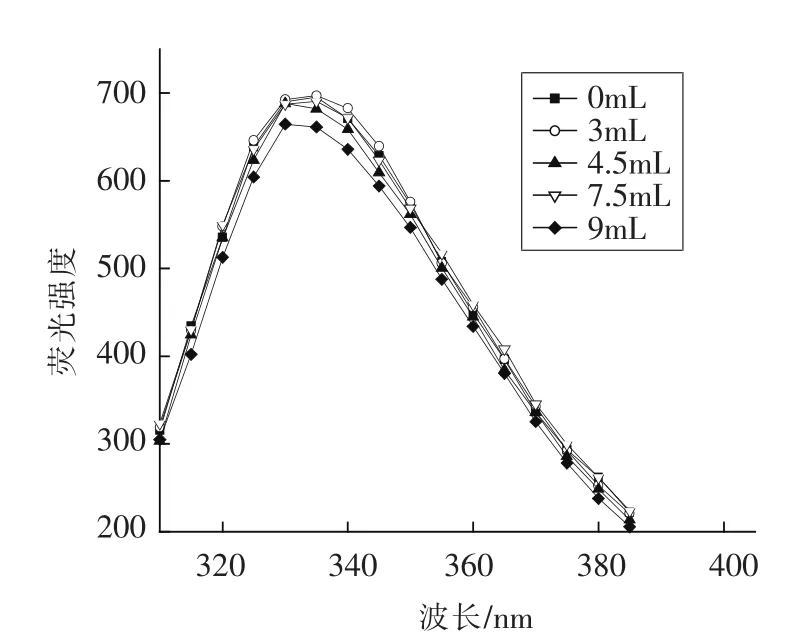
图3 不同亚油酸添加量对WPI乳状液内源荧光的影响Fig.3 Effects of different amount of linoleic acid added on fluorescence intensity of WPI emulsion

表2 不同亚油酸添加量对WPI乳状液荧光λmax和荧光强度的影响Table2 Effects of different amount of linoleic acid added on λmaxand fluorescence intensity of WPI emulsion
2.2.3 核桃蛋白乳状液乳液粒度分析 乳液液滴大小与蛋白质的乳化能力[30]和分子间粒子的疏水作用[31]有关。乳剂的液滴粒径影响乳化稳定性,以及蛋白质乳液的货架期[32]。形成乳液的过程中,蛋白质可以促进油相与水相的结合,乳液中油滴的粒径大小可以直观的展示乳液中油滴的分散状态[33]。d3,2反映乳液中的小颗粒组分,d3,2越小,其乳液所对应的乳化活性较好[34]。d4,3反映蛋白质帮
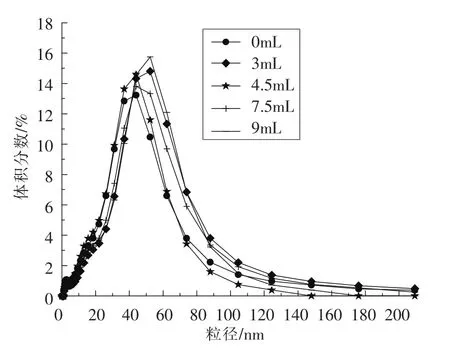
图4 不同亚油酸添加量对WPI乳状液乳液粒度大小的影响Fig.4 Effects of different amount of linoleic acid added on drop size of emulsion of WPI emulsion
2.2.4 核桃蛋白乳状液微观结构分析 电子显微镜显示了蛋白质的微观结构。图5所示亚油酸添加量不同,乳液油滴聚集形态不同。亚油酸添加量在0~4.5 mL之间,液滴大小逐渐增加。随后,随着氧化程度的增加,乳液液滴颗粒又变得密集。当亚油酸添加量达到9 mL时,液滴尺寸变得最大。液滴聚结和絮凝出现在被过度氧化的WPI稳定的乳剂中。这一观察结果正好与上述阐述的乳状液助油相分散于水相的乳化能力,d4,3越小,其乳化能力越好[35]。如图4所示,亚油酸添加量从0~4.5 mL 时,乳状液 d4,3、d3,2的值有所增加,当亚油酸添加量至7.5 mL时,d3,2下降到最小,对应乳化活性最高。由此说明适度的氧化使蛋白质结构展开,暴露出更多的疏水基团,形成可溶性聚合物[24]。这与Poon[34]报道的d3,2越小,乳液的乳化活性越好相一致。液滴大小的结果一致。乳液液滴水相中的蛋白质,在油水界面发生聚集,随着氧化程度的增加,液滴变大,蛋白质聚合程度增加。这表明,氧化影响表面液滴的均匀分布。在乳液中,只有30%~40%覆盖蛋白的界面,因为只有一部分的疏水基团覆盖在油水界面上,其它的蛋白肽存在于水相中[36]。乳化的过程,是在油水界面上由界面张力驱动的分子重组和蛋白质重排的过程[7]。
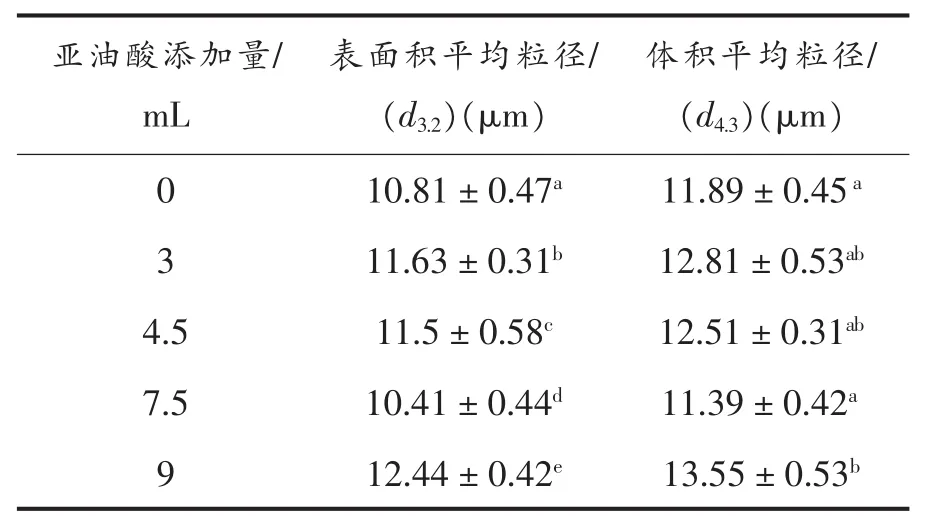
Table3 不同亚油酸添加量对WPI乳状液乳液粒度的影响Table3 Effects of different amount of linoleic acid added on drop size of emulsion of WPI emulsion

图5 不同亚油酸添加量对WPI乳状液微观结构的影响Fig.5 Effects of different amount of linoleic acid added on microstructure of WPI emulsion
2.3 蛋白氧化结构对功能的相关性分析
皮尔逊相关性分析在结构和乳化功能指标之间建立相关联系。皮尔逊相关系数清楚地表明,蛋白质结构如羰基变化,可以影响其它结构之间如巯基值、总巯基值与表面疏水性显著变化,且与EAI呈显著负相关,但与ESI不相关。WU等[37]报道氧化导致大豆蛋白质二级和三级结构的显著变化,从而导致功能属性的增加或下降,但此结果表明,结构的改变,对功能性质的影响并不显著,这可能与HPODE氧化体系对蛋白质的影响活性较低有关。
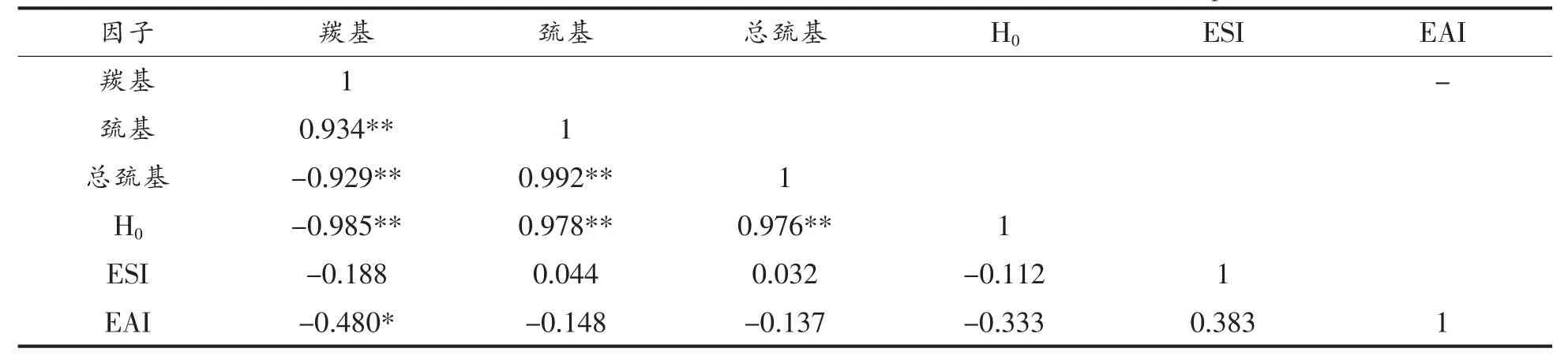
表4 乳化性质和氧化核桃蛋白结构的Pearson相关系数Table4 Pearson correlation coefficient of emulsifying property and oxidized walnut protein structure
3 结论
LOX催化亚油酸形成HPODE,代表LOX诱导脂质过氧化反应中形成脂质氢过氧化物。HPODE与蛋白质分子中含ε-氨基群的赖氨酸残基、含巯基群的半胱氨酸残基反应,生成共价结合物,最终导致蛋白质羰基化,蛋白质巯基群含量下降。表面疏水性由于氧化导致蛋白质聚集,暴露的疏水残基共价修饰,引入新的亲水组分,致使表面疏水性下降。从粒径分布结果显示,随着亚油酸添加量的增加,疏水基团共价修饰及二硫键共价键导致聚集体的形成。由此得出,HPODE氧化会改变蛋白质的结构。HPODE对蛋白质乳化功能的影响也做了进一步研究,EAI/ESI结果显示,适当的氧化浓度有利于提高核桃蛋白的乳化活性和乳化稳定性。内源荧光光谱分析监测了核桃分离蛋白乳液的构象改变,结果显示出核桃蛋白的吸光度改变,由此说明色氨酸的化学环境发生了改变。乳状液乳液粒度中,d3,2反映乳液中的小颗粒组分,其表征样品的乳化活性,当亚油酸添加量至7.5 mL时,d3,2下降到最小,对应乳化活性最高。电子显微镜显示了蛋白质的微观结构,亚油酸添加量不同,乳液油滴聚集形态不同。在结构和乳化功能指标之间建立皮尔逊相关性系数,结果表明蛋白质结构的改变,对功能性质的影响并不显著。
