镉胁迫对高低积累型水稻幼苗非蛋白巯基含量的影响
2019-12-23李冬琴王丽丽李智鸣黎华寿陈桂葵
李冬琴,王丽丽,李智鸣,黎华寿,陈桂葵*
(1.岭南师范学院生命科学与技术学院,广东 湛江524048;2.华南农业大学农业农村部华南热带农业环境重点实验室,广州510642;3.广州草木蕃环境科技有限公司,广州510640)
镉(Cadmium,Cd)是生物非必需元素,其毒害效应已受到人们的广泛关注[1-2]。Cd进入土壤环境中容易被植物吸收,可通过食物链进入人体,不仅造成贫血、高血压、骨痛病等疾病,而且还具有潜在的致癌作用,严重威胁食品安全、人体健康及农业的可持续发展[3-4],是被公认的对人类最具威胁的主要有毒金属之一。由于Cd的移动性强、毒性高、污染面积大而被称为“五毒之首”[5]。长期以来,植物对重金属污染的抗性(耐性)问题一直是污染生态学等研究领域的热点问题。
水稻(Oryza sativaL.)是我国,尤其是南方地区主要的粮食作物,且品种繁多[6]。据研究,水稻品种不同对Cd 的吸收积累存在很大差异[7]。Cd 胁迫诱导植株体内产生活性氧(Reactive oxygen species,ROS),使细胞受到伤害,导致代谢紊乱,因此破坏植物正常的生理生化功能。为了抵御Cd 的毒害作用,植物进化了一系列抗性机制,如抗氧化酶和抗氧化剂清除活性氧的毒害;抗坏血酸-谷胱甘肽循环(AsA-GSH)系统维持细胞氧化还原平衡:液泡的区室化和外排机制;植物体内螯合物与Cd结合解除Cd的毒害[8-10]。
非蛋白巯基(Non-protein thiols,NPT)化合物主要包括还原型谷胱甘肽(Reduced glutathione,GSH)、植物螯合肽(Phytochelatins,PCs)和半胱氨酸(Cysteine,Cys)等,是植物体内的一类多肽。植物体内的GSH、PCs、Cys 以及金属硫蛋白(Metallothionein,MTs)等巯基物质可与细胞内游离的Cd离子配位结合转化成络合态存在于细胞质或运输到液泡中[11],从而减轻细胞质受到的Cd 毒害。PCs 在对Cd 的解毒、防御Cd引起的氧化胁迫和代谢方面具有重要作用。早在1990 年Verkleij 等[12]的报道中就表明,根系所吸收的Cd 至少有60%是以PC-Cd 复合物形式存在[12]。在水稻(Oryza sativa)、谷子(Setaria italicaL.)、拟南芥(Arabidopsis thaliana)、小麦(Triticum aestivum)、蓖麻(Ricinus communis)、大豆(Glycine max)等植物中证实了巯基物质在Cd 的螯合、固定等方面起着重要作用[13-18]。PCs是在植物螯合肽合成酶的作用下以GSH为底物合成的多肽。GSH是低分子量的抗氧化剂,对植物新陈代谢的维持起到很重要的作用[19]。GSH 具有清除自由基、抵抗过氧化损伤、保护酶和结构蛋白及膜系统的功能[11]。此外,与GSH代谢密切相关的解毒酶——谷胱甘肽硫转移酶(Glutathione S-transferase,GST)在Cd 解毒过程中也发挥了重要作用[20]。GSH 在GST 的作用下可与细胞内的Cd离子络合形成复合物,将有毒物质转移到液泡内[21-22],以达到缓解Cd 毒害的作用。范业赓等[23]发现,甘蔗叶片在Cd 胁迫下GSH 和NPT 含量有上升的趋势,产生的GSH 和NPT可以有效地清除活性氧及自由基,阻止膜脂过氧化。高可辉[24]的研究表明,水稻幼苗体内的GSH含量随处理时间的延长而逐渐增加,同样说明GSH 对水稻解除Cd 的毒害发挥重要作用。然而,水稻品种不同在Cd 胁迫下的解毒过程不同,巯基物质含量对水稻Cd 污染较敏感[25]。Cd 胁迫对Cd 高低积累型水稻非蛋白巯基含量的影响机制研究不足,不利于指导解决水稻种植过程中遭受Cd胁迫的问题。
本实验基于前期研究筛选出的高低Cd积累型水稻品种[26],通过营养液沙培的方式,研究高低Cd 积累型水稻品种Cd 含量与非蛋白巯基化合物之间的关系,以及在Cd 胁迫下NPT(包括GSH 和PCs)和GST活性的动态变化规律,为揭示不同积累型水稻Cd 解毒的生理机制提供理论依据,也可为耐Cd 胁迫且低Cd 积累的水稻品种的选用和选育提供有效的生物学途径。
1 材料与方法
1.1 实验设计
本实验选取的材料为前期实验筛选出的Cd高积累型水稻品种欣荣优2045 和低积累型水稻品种优I2009[26]。采用营养液沙培方法,探讨Cd 胁迫下不同耐性水稻中NPT、GSH、PC、GST、过氧化氢(H2O2)和硫代巴比妥酸反应产物(TBARS)的动态变化。培养基质为经干热灭菌的河沙(150 ℃灭菌2 h,灭菌后50 ℃烘干备用)。将灭菌后的河沙装入双层嵌套使用的塑料杯(透明塑料杯的体积为700 mL,规格为9 cm×6.5 cm×10.5 cm),其中里面的塑料杯底部穿孔以方便更换营养液,外层塑料杯四周和底部用黑色遮光袋包住,以免光照对根系生长造成影响。
挑选饱满的水稻种子用10%的H2O2消毒10 min,用无菌水清洗干净,以除去H2O2,再用蒸馏水浸泡48 h,期间每日换水4 次。待种子露白后在温度为25 ℃,相对湿度为75%的人工气候箱暗箱中催芽24 h,挑选发芽一致的水稻种子均匀播种于装好河沙的塑料杯中(1 kg·杯-1)。营养液参照国际水稻研究所(IRRI)推荐的水稻常规营养液配方配制(表1)。将水稻放置于人工气候箱中进行培养,条件为:光照时间为14 h·d-1,光照强度为3000 lx,昼/夜温度为28 ℃/25 ℃,昼/夜相对湿度为80%/75%。每2 d更换一次营养液。待幼苗长至四叶一心时,进行CdCl2处理。根据文献研究设定Cd2+处理浓度为50 μmol·L-1。样品分别在处理前(0,对照)和第1、3、5、7 d收获。每个处理重复3次,每杯种植水稻2株。
1.2 测定方法
1.2.1 植物样品的前处理
水培实验结束后,小心采集水稻幼苗样品,用自来水洗净根系河沙及地上部杂物后用去离子水彻底清洗,然后将幼苗从基部剪开,分为根部和地上部。样品一部分用于Cd含量的测定,另一部分存于-80 ℃超低温冰箱冷藏供各项生理生化指标测定。
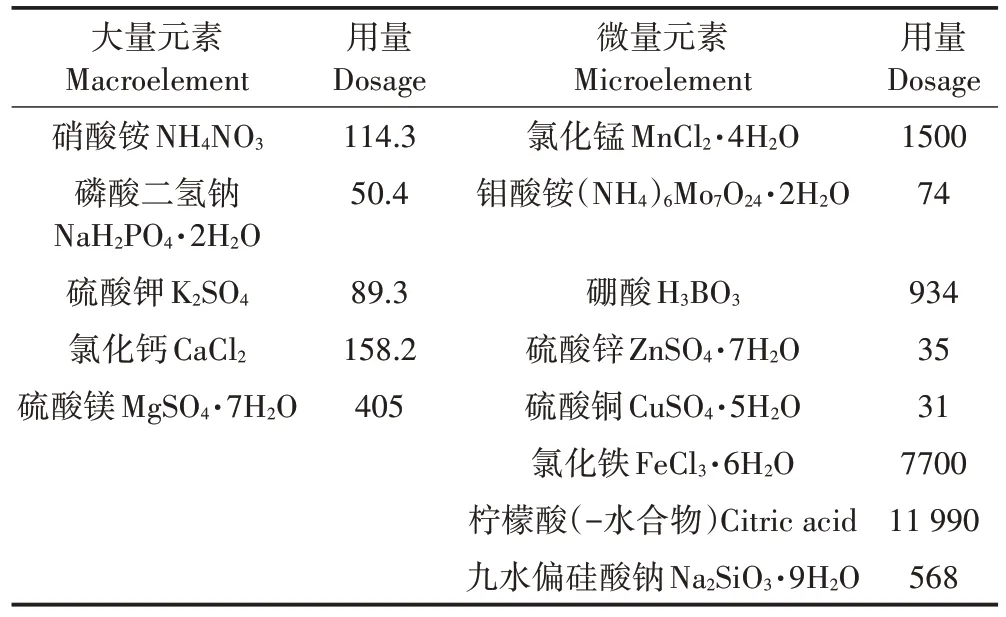
表1 水稻常规营养液配方(mg·L-1)Table 1 Conventional nutrient solution formula for rice(mg·L-1)
1.2.2 植物样品中Cd含量的测定
称取0.1 g 鲜样,用硝酸和高氯酸的混酸(4∶1)消解后,用石墨炉原子吸收(ZEEnit700P,德国耶拿)测定样品中Cd含量。
1.2.3 GSH和GST活性的测定
GSH 和GST 活性采用南京建成试剂盒推荐方法测定。蛋白含量测定用考马斯亮蓝染色法[27],以结晶牛血清蛋白(BSA)为标准蛋白。组织中GST活性(U·mg-1prot):规定每毫克组织蛋白在37 ℃反应1 min,扣除非酶促反应,使反应体系中GSH 浓度降低1 μmol·L-1为一个酶活力单位。
1.2.4 NPT 和PCs 含量的测定
称取1.00 g 水稻鲜样,加4 mL 5%磺基水杨酸(SSA)冰浴研磨,12 000g离心15 min(4 ℃),上清液定容至10 mL。分袋保存。
NPT 物质含量测定采用Rama Devi 等[28]的方法:反应体系为1 mL 上清液与1.85 mL 0.2 mol·L-1的Tris-HCl(pH 8.2)和0.15 mL 10 mol·L-1的5,5-双二硫(2-硝基苯甲酸)(DTNB),反应20 min 后用多功能酶标仪于412 nm 处测定吸光值,以不加DTNB 为空白。以GSH为-SH标样,做标准曲线。
PCs含量用差减法计算[29],即:PCs=NPT-GSH
1.2.5 H2O2和TBARS含量的测定
称取1.00 g 鲜样,加4.8 mL 5%三氯乙酸(TCA)冰浴研磨,12 000g离心15 min(4 ℃),上清液定容至10 mL。分装保存。
H2O2的测定参照Jana 等[30]的方法:取1 mL 上清液,加0.3 mL 0.1%(m/V)硫酸钛[溶于20%(V/V)硫酸],410 nm下读数(25 ℃)。
TBARS 含量参照Heath 等[31]的测定方法:取2 mL上清液,加2 mL 0.5%(m/V)TCA 于试管,摇匀后保鲜膜封口,95 ℃水浴30 min,冰浴终止。由532 nm 和600 nm下吸收度的差值计算TBARS含量。
1.2.6 数据分析
实验数据以平均数±标准误(Mean±Stand error)表示,采用Microsoft Excel 2010和SPSS 17.0进行数据计算和统计分析,并用Duncan′s 检验法进行显著性分析。
2 结果与分析
2.1 Cd胁迫下高低积累型水稻品种间Cd吸收积累量的差异
随Cd 胁迫时间的延长,两品种水稻对Cd 的吸收积累均呈显著上升趋势(图1)。在处理的第7 d 时,欣荣优2045 和优I2009 地上部对Cd 的吸收量均达到最大值,分别为59.80 mg·kg-1FW 和20.74 mg·kg-1FW,与第1 d 相比,增加了1.92 倍和3.10 倍;欣荣优2045 和优I2009 的根系对Cd 的吸收量也在第7 d 时达到最大值,分别为252.84 mg·kg-1FW 和126.54 mg·kg-1FW,与第1 d 相比,增加了2.26 倍和1.84 倍。而且各个处理时间段,无论是地上部还是根系,欣荣优2045对Cd的吸收积累均显著高于优I2009。
2.2 Cd胁迫下高低积累型水稻品种间H2O2含量的差异
如图2所示,两品种水稻的H2O2含量随处理时间的延长呈逐渐上升趋势,第7 d 时,欣荣优2045 和优I2009 地上部的H2O2含量达最高值,分别为6.57 μmol·g-1和9.19 μmol·g-1,相较于对照,分别上升了45%和78%;欣荣优2045 和优I2009 根系H2O2的含量在第7 d 时也均达到最高值,分别为4.89 μmol·g-1和6.28 μmol·g-1,相较于对照,分别上升了1.14倍和1.32倍。综上所述,优I2009 体内H2O2含量上升的幅度大于欣荣优2045,说明随处理时间的延长,优I2009 受Cd的毒害作用更大。
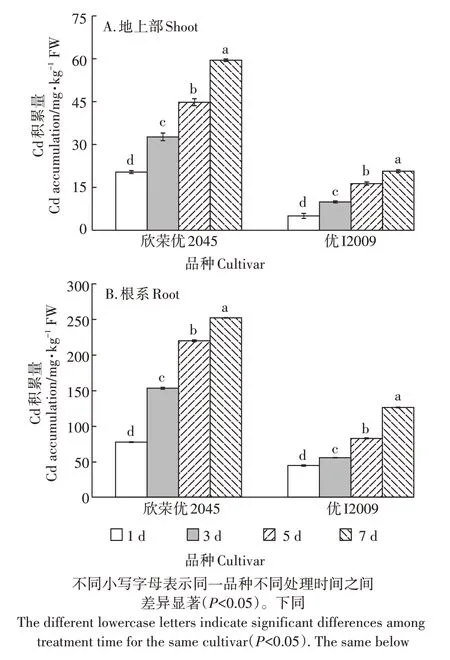
图1 水稻幼苗积累Cd的动态变化Figure 1 Dynamic changes of Cd accumulation in rice seedling
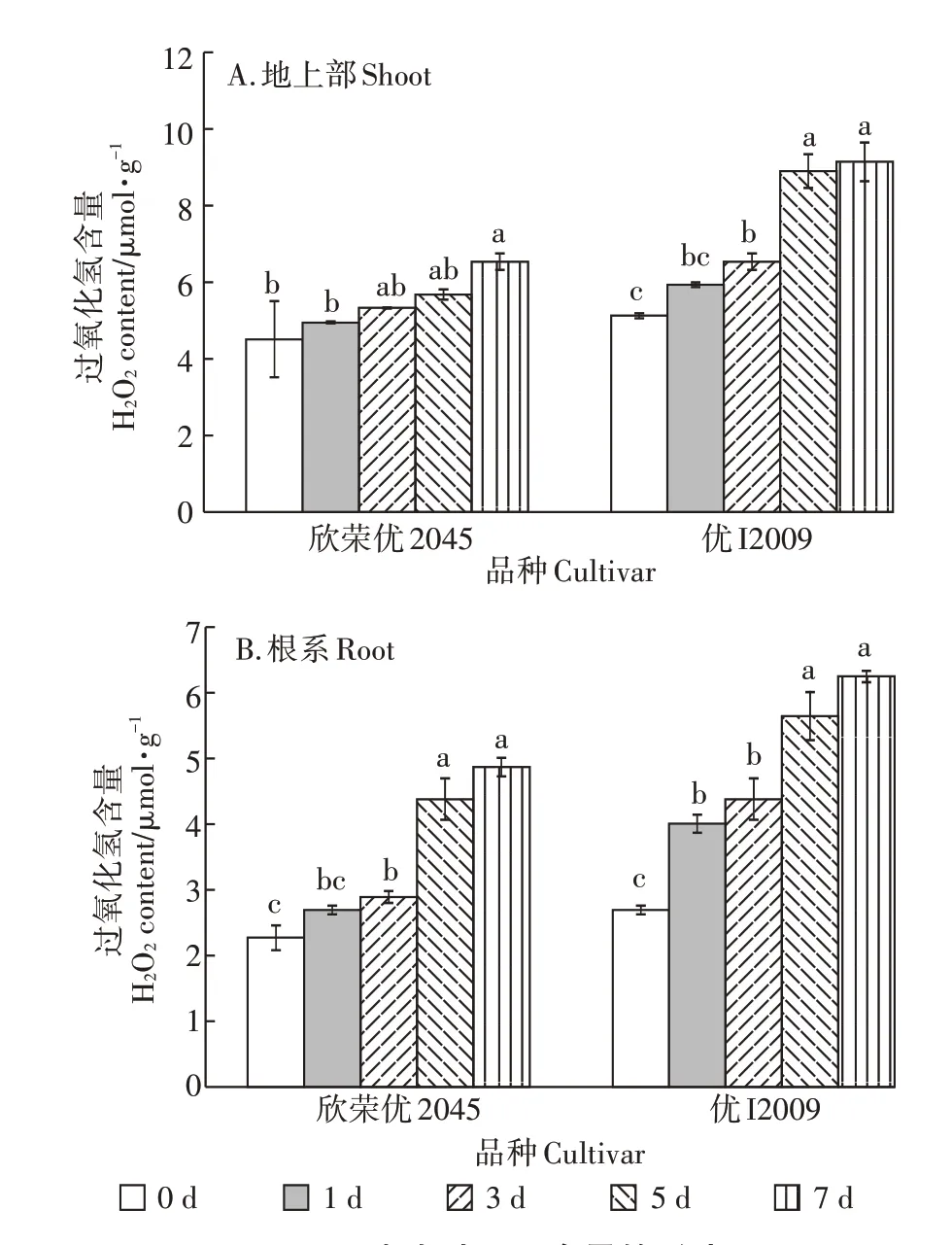
图2 Cd胁迫对H2O2含量的影响Figure 2 Effect of cadmium stress on H2O2content
2.3 Cd 胁迫下高低积累型水稻品种间TBARS 含量的差异
随时间延长,两品种水稻地上部和根系的TBARS 含量的变化趋势与H2O2相似(图3)。欣荣优2045 地上部的TBARS 含量在第1、3、5、7 d 比对照分别增加了7%、29%、59%和74%,优I2009 地上部的TBARS 含量分别上升了12%、56%、72%和120%;欣荣优2045 根系TBARS 的含量在第1、3、5、7 d 比对照分别上升了6%、19%、28%和61%,优I2009 根系的TBARS 含量分别上升了22%、32%、53%和64%。综上所述,优I2009体内TBARS 含量上升的幅度大于欣荣优2045,同样说明优I2009 水稻品种对Cd 的耐受性相较于欣荣优2045更弱。
2.4 Cd胁迫下高低积累型水稻品种间NPT含量的差异
如图4 所示,随Cd 处理时间的延长,两品种水稻地上部和根系NPT 的含量呈先升后降的趋势。NPT含量在第5 d 时达到最大值,与其对照相比,欣荣优2045 和优I2009 地上部的NPT 含量分别增加了107%和48%,根系含量增加了1.38 倍和5.13 倍。在第7 d时,两品种水稻的NPT 含量均显著下降。在各个Cd处理时间段,欣荣优2045 的NPT 含量(尤其是根系)均高于优I2009。
2.5 Cd胁迫下高低积累型水稻品种间GSH含量的差异
随Cd 胁迫时间的延长,欣荣优2045 和优I2009体内GSH 含量的变化趋势差异很大(图5)。欣荣优2045 体内的GSH 含量呈逐渐上升趋势,第7 d 时达到最大值,地上部和根系分别为178.43 nmol·g-1和161.97 nmol·g-1,相较于对照分别增加了2.09 倍和1.06 倍;而优I2009 体内的GSH 含量则呈先升后降的变化趋势,第3 d时达到最大值,地上部和根系分别为118.35 nmol·g-1和149.23 nmol·g-1,相较于对照增加了86%和2.33 倍,第7 d 时,地上部和根系含量分别降为79.97 nmol·g-1和64.48 nmol·g-1,相较于第3 d 分别下降了32%和57%。
2.6 Cd胁迫下高低积累型水稻品种间PCs含量的差异
两种水稻地上部和根系PCs 含量的变化与NPT含量的变化趋势一致(图6)。欣荣优2045和优I2009地上部的NPT 含量在第5 d 时达到最大值,分别为420.35 nmol·g-1和437.76 nmol·g-1,与其对照相比增加了97%和47%;而欣荣优2045 和优I2009 根系的PC 含量分别为720.05 nmol·g-1和508.38 nmol·g-1,与其对照相比增加了1.51 倍和8.37 倍。在第7 d 时,两个水稻品种的PCs含量均显著下降。
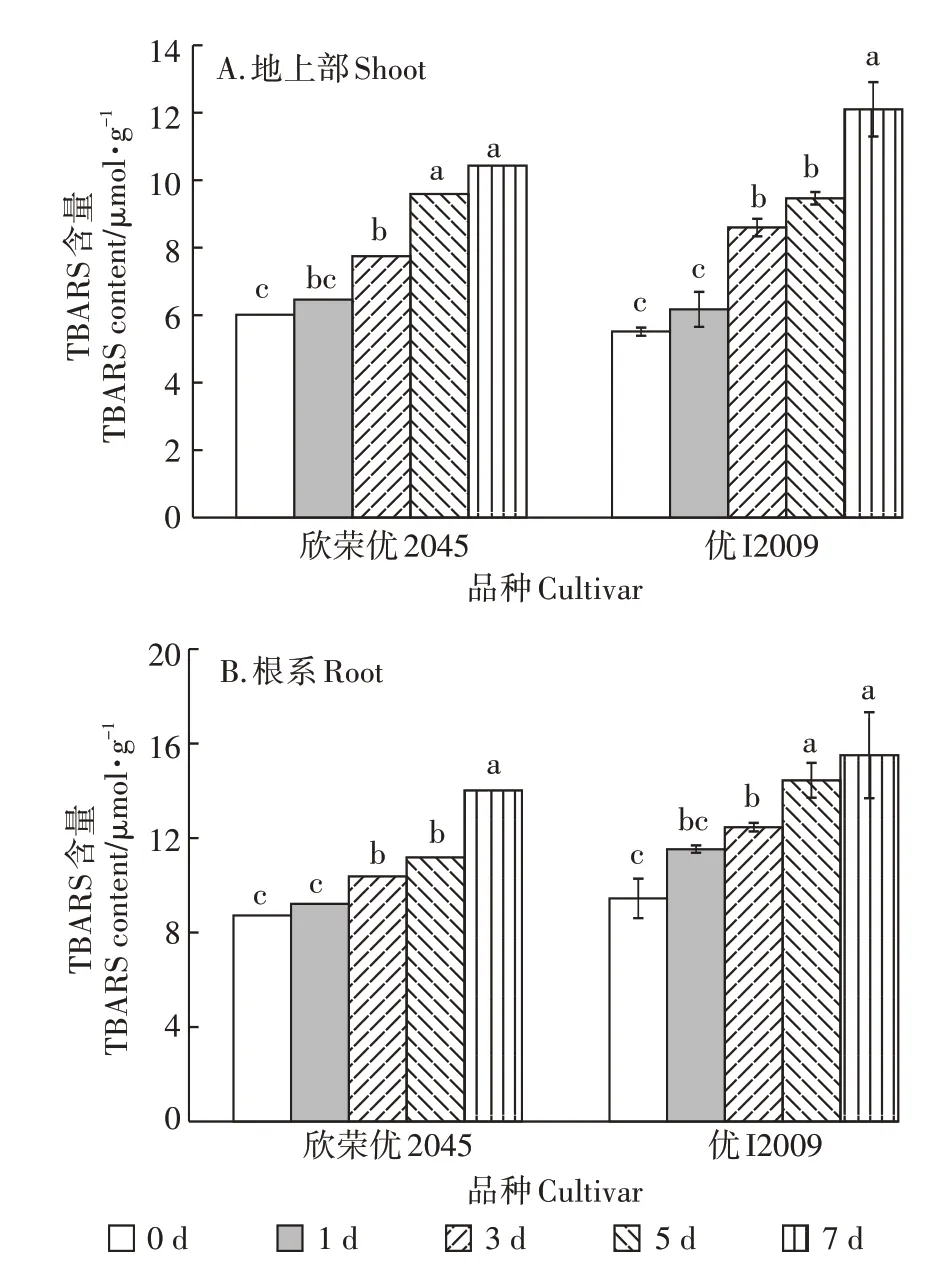
图3 Cd胁迫对TBARS含量的影响Figure 3 Effect of cadmium stress on TBARS content
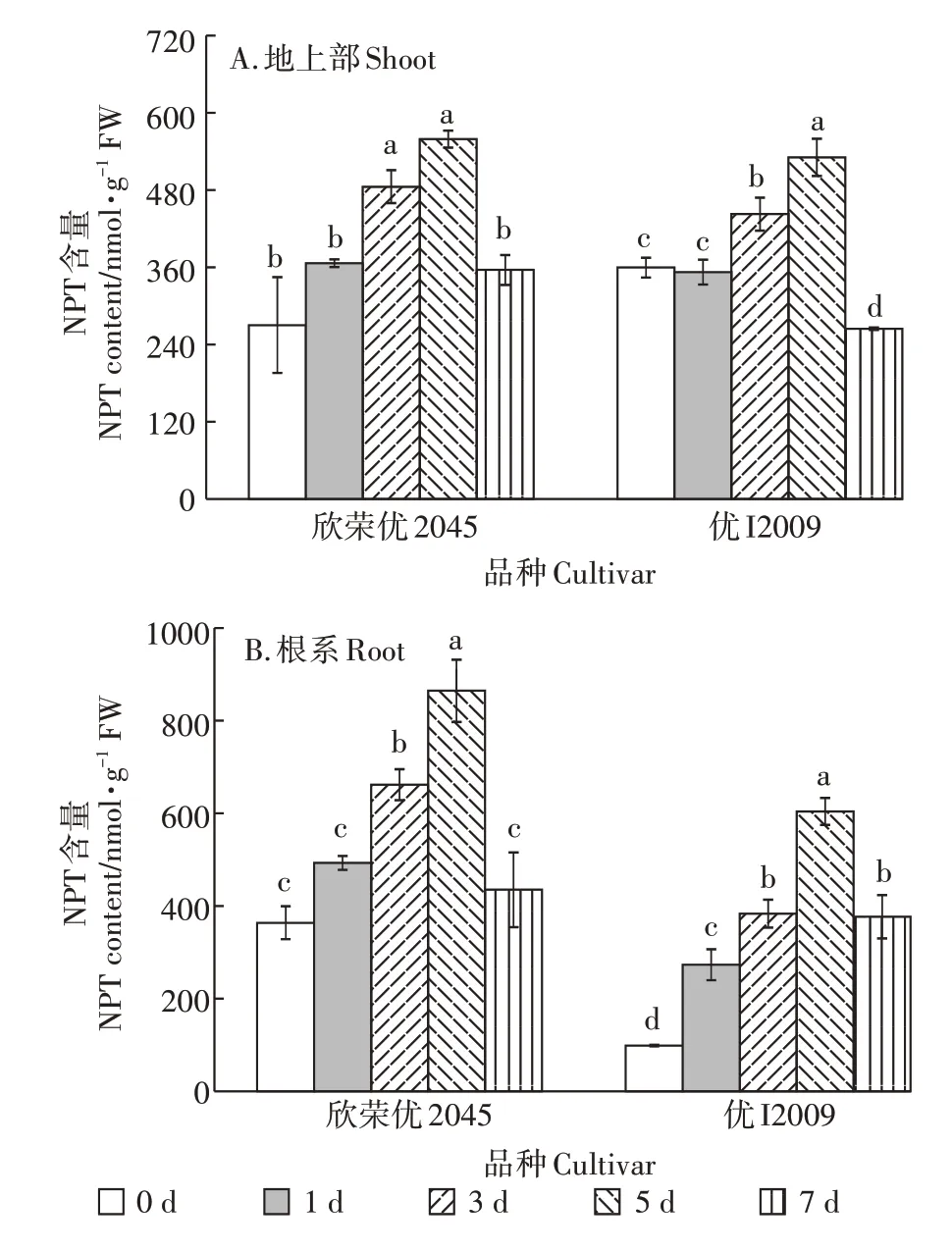
图4 Cd胁迫对NPT含量的影响Figure 4 Effect of cadmium stress on NPT content
2.7 Cd胁迫下高低积累型水稻品种间GST活性的差异
随Cd 胁迫时间的延长,两品种水稻地上部GST活性都呈先升后降的趋势(图7),第3 d 时活性均达到最高,为145.90 U·mg-1(欣荣优2045)和146.67 U·mg-1(优I2009),比对照上升了70%(欣荣优2045)和129%(优I2009),第3 d 后GST 活性显著下降,到第7 d时,下降了31%(欣荣优2045)和51%(优I2009)。
两品种水稻根系GST活性亦呈先升后降的趋势,其中GST 活性最大的是第3 d 和第5 d,欣荣优2045的GST 活性分别为287.33 U·mg-1和282.20 U·mg-1,比对照增加了95%和91%;优I2009 的GST 活性分别为264.01 U·mg-1和266.94 U·mg-1,比对照增加了1.11倍和1.13 倍。到第7 d 时,两品种水稻的GST 活性均显著下降,相较于第5 d,欣荣优2045 和优I2009 分别下降了36%和50%。
3 讨论
同一条件下,不同品种水稻间和不同器官间Cd的吸收和积累存在差异。这种差异主要是由水稻内外部形态结构的差异、不同品种对Cd 的生理生化反应机制以及水稻植株对Cd 吸收转运机制的不同所致[32]。本研究中,随Cd 胁迫时间的延长,两品种水稻对Cd 的吸收积累量为欣荣优2045>优I 2009,且根部>地上部,这与我们前期的研究一致[26]。
在Cd 胁迫下,植物体内产生的大量ROS 导致植物体内代谢紊乱。植物细胞产生的H2O2是ROS 的主要产物,其含量可反映机体内脂质过氧化程度和细胞损伤程度,而TBARS 是植株体内脂类过氧化物的最终产物,其含量水平通常可用来反映植株遭受氧化胁迫的水平。随Cd 胁迫时间的延长,两品种水稻的H2O2和TBARS 含量逐渐上升,而Cd 低积累品种优I2009 体内的H2O2和TBARS 含量上升的幅度大于欣荣优2045(图2 和图3),说明优I2009 受Cd 的毒害作用比欣荣优2045更大。
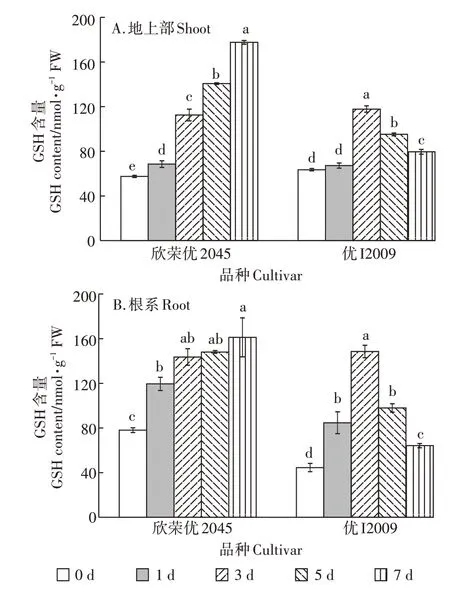
图5 Cd胁迫对GSH含量的影响Figure 5 Effect of cadmium stress on GSH content
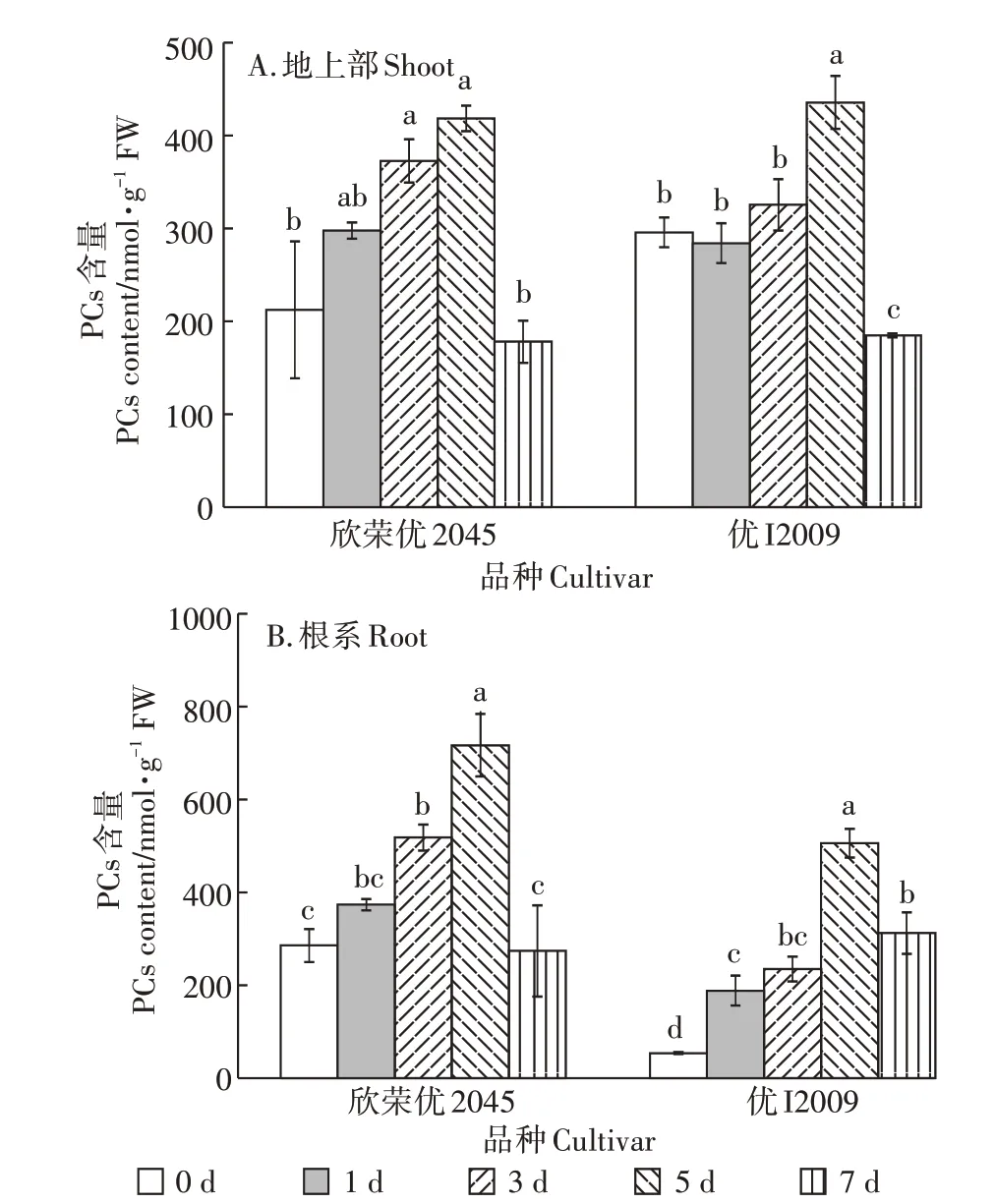
图6 Cd胁迫对PCs含量的影响Figure 6 Effect of cadmium stress on PCs content
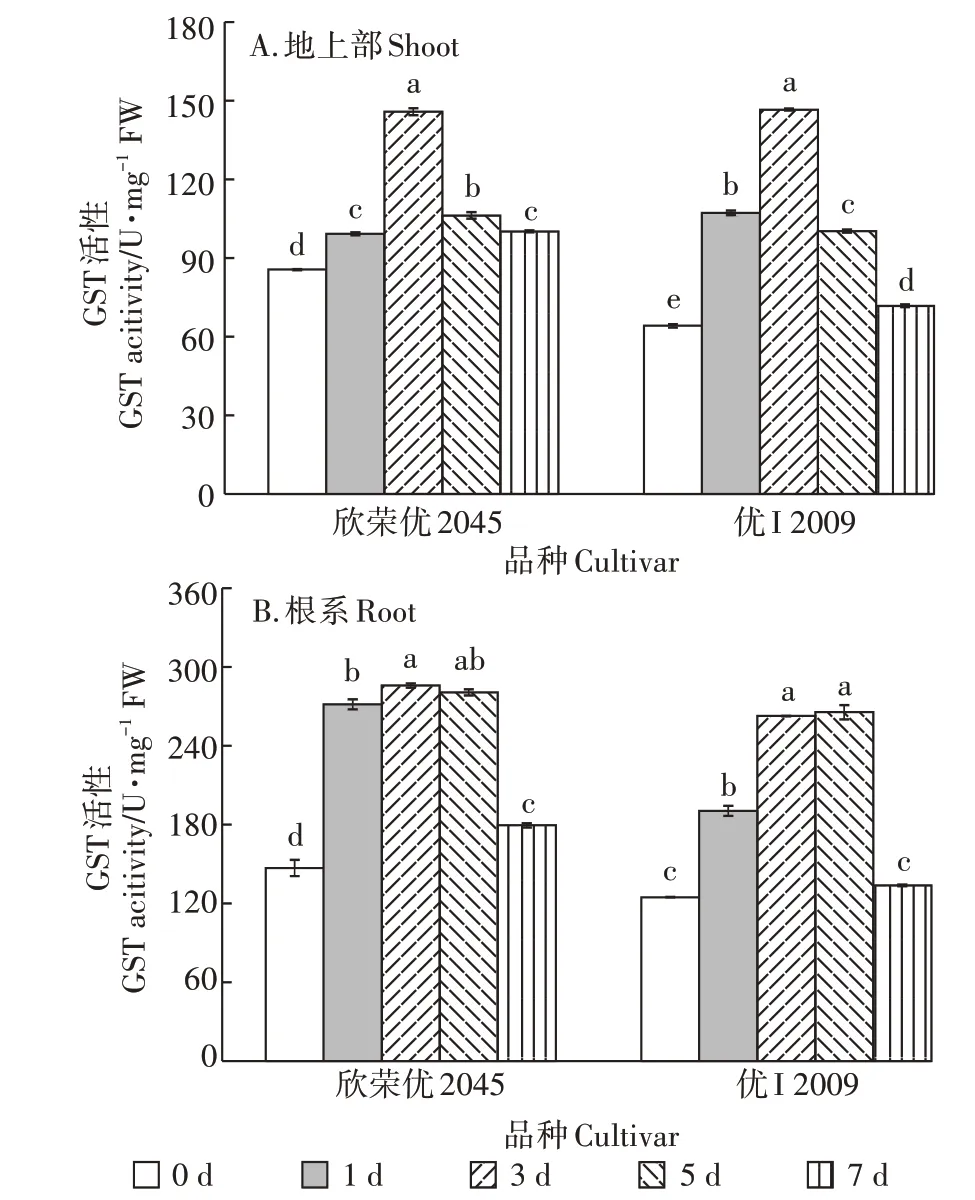
图7 Cd胁迫对GST含量的影响Figure 7 Effect of cadmium stress on GST content
为抵御Cd 胁迫,植物体内产生的非蛋白巯基物质(GSH、PCs、Cys 等)络合Cd 是重要的解毒机制之一[11]。巯基物质与Cd 形成的络合物减少了细胞质中自由Cd离子的浓度,或将其运输到液泡中区隔,形成区室化作用[21-22,33]。GSH是植物体内重要的抗氧化剂和信号物质[11],是在生物体胞内代谢过程和遭受氧化胁迫时产生的H2O2的最有效清除剂之一[34]。本研究中,随Cd 胁迫时间的延长,欣荣优2045 的GSH 含量逐渐上升,与H2O2活性呈正相关,而优I2009 表现出先升后降的趋势,说明欣荣优2045 在清除活性氧的功能方面表现出更强的解毒能力,而优I2009在Cd胁迫前期表现出对Cd 胁迫的应激反应,但在Cd 胁迫后期可能是因为过多ROS 的生成超出了其清除范围,因此GSH含量逐渐下降。两品种水稻随Cd胁迫时间的延长,NPT 和PCs 含量呈先上升后下降的趋势,说明Cd 胁迫后水稻幼苗可通过诱导前期(1~5 d)NPT和PCs 的合成来减轻Cd 对水稻幼苗的毒害作用,这属于水稻幼苗应对Cd胁迫的自救机制。并且欣荣优2045的NPT含量高于优I2009,说明Cd高积累型品种合成更多的NPT 与Cd 络合,解毒能力更强[25]。Cd 胁迫后期,NPT 和PCs 的含量下降,可能的原因是随Cd胁迫时间的延长,Cd 引起的过氧化损伤不断加剧,打破了植物体内ROS 产生和清除的动态平衡,使其诱导NPT 和PCs合成的能力下降,说明两种水稻NPT 和PC的合成能力存在一个Cd胁迫时间的阈值。
GST是生物体内重要的解毒酶,与GSH代谢密切相关,可保护细胞免受Cd的毒害[20]。GST还具有过氧化物酶的功能,能平衡H2O2的产生和清除[35]。本研究中,两品种水稻随Cd 胁迫时间的延长,GST 活性呈先上升后下降的趋势,因为在Cd胁迫下,H2O2等活性氧不断生成,而在植物防御机制中起着重要作用的GST活性也随之升高,以减轻Cd 的毒害作用,但随Cd 胁迫时间的延长,GST 活性下降,表明水稻抗氧化系统对Cd 的耐受性有一定的时间限值,超过这一时间限值时,未被清除的活性氧会对细胞产生不可逆的损伤,GST 的解毒作用被抑制,致使其酶活性下降。通过比较两品种水稻的GST 活性(图7)发现,同一时间段欣荣优2045 高于优I2009,说明不同Cd 积累型水稻的GST 活性对Cd胁迫的应答机制存在一定的基因型差异,这可能与两品种GST 同功酶的组成、表达和功能不同有关[20,36-37]。
4 结论
随Cd 胁迫时间的延长,两品种水稻的H2O2和TBARS 含量逐渐增加,同时也不同程度地刺激了两品种水稻NPT(包括GSH 和PCs)的合成和GST 的生成,且Cd 低积累型品种优I2009 遭受的氧化胁迫水平和膜脂过氧化程度大于Cd 高积累型品种欣荣优2045。但是,当Cd 胁迫超过一定的时间限值时,两品种水稻所合成的NPT 和生成的GST 活性均被抑制。此外,Cd 高积累型水稻品种欣荣优2045 可以通过合成更多的NPT 与Cd 络合来清除活性氧,相较于Cd 低积累型水稻品种,优I2009 表现出更强的解毒能力。
