铁膜和磷作用对水稻根尖细胞壁耐铝性的影响
2019-12-23王智怡李文静邢承华王亚超陈超发蔡妙珍
王智怡,李文静,邢承华,李 方,王亚超,陈超发,蔡妙珍*
(1.浙江师范大学地理与环境科学学院,浙江 金华321004;2.金华职业技术学院农业与生物工程学院,浙江 金华321007)
铝毒是酸性土壤中限制作物生长最普遍的非生物因素之一[1]。作物在长期的进化过程中,逐渐形成了如分泌螯合物(有机酸、酚类和黏液)[2]或磷酸根[3]、细胞壁固定[4]、根系pH 提高[5]、根尖边缘细胞[6]以及根表铁膜[7]等耐铝能力以适应铝毒环境。根尖细胞壁作为植物响应铝毒的第一道屏障,既是铝毒害的最初位点,也是铝(Al)积累的主要部位,其在植物耐铝毒中的作用倍受关注[8-9]。细胞壁主要由多糖(果胶、半纤维素)组成,其中果胶的基本结构是聚半乳糖醛酸,其上大量带负电荷的羧基是Al 结合的主要位点[10]。现有的研究中已经发现,豌豆(Pisumsativum)[11]、紫花苜蓿(Medicago sativa)[12]和水稻(Oryza sativaL.)[13]根尖的细胞壁果胶含量与根尖Al 积累显著正相关。Yang 等[5]指出拟南芥(Arabidopsis thaliana)细胞壁结合的Al 有75%分布在半纤维素,20%分布在果胶。可见,植物细胞壁的多糖组分对根系上Al 的积累起到了至关重要的作用。
淹水环境中水生植物根系具有特殊的通气组织能将地上部的氧气运输至根系,释放的氧气把根系周围的Fe2+氧化形成铁氧化物胶膜(铁膜,IP)附着在植物根表[14-15]。铁膜属于两性胶体,与自然界中的铁氧化物具有相似的特征,它可以通过吸附、共沉淀等作用,改变Cd、Pb、As等重金属[16-17]及氧化铜纳米颗粒[18]的存在形态,影响其生物有效性。研究已经证明,铁膜能将Al 固定在水稻根表,阻止根系对Al 的吸收以及向地上部的运输[19]。磷(P)作为植物生长发育的必需元素之一,在植物Al 的吸收和运输中也起到了一定的作用,如于姣妲等[20]发现磷能够改变杉木(Cunninghamia lanceolata)幼苗细胞内抗氧化酶的活性以及离子转运,增加Al向液泡的转移,降低Al在细胞壁中的积累。此外,细胞壁组分也参与了和磷相关的抗铝毒过程。例如,在缺磷状态下,水稻根系的磷脂和果胶的含量降低或者酸性磷酸酶(APA)活性提高以提供更多的磷与Al结合钝化,由此提高耐铝性[21]。磷充足时,磷通过与Al 在荞麦根细胞壁中形成不溶性沉淀[22]或通过降低耐铝性水稻细胞壁多糖含量以减少Al结合位点[23],表明铝毒胁迫下磷能通过改变根系细胞壁组分提高耐铝性。
以往的研究主要侧重于铁膜或磷单一因素对水稻耐铝性的影响,而铁膜以及吸附于铁膜上的磷对水稻根尖细胞壁组分和耐铝性的影响及其相互作用需要进一步探索。因此,本研究以耐铝能力有显著差异的两个水稻基因型为材料,研究铝毒作用下铁膜和覆铁膜供磷对水稻生长以及细胞壁多糖含量的影响,从而进一步探究铁膜和磷对水稻铝耐性的交互作用。
1 材料与方法
1.1 试验材料
水稻(Oryza sativaL.)品种选用菲优多系1 号(耐铝毒基因型)和红良优166(铝毒敏感基因型),种子由三才种业公司提供[19]。
1.2 植株培养及试验处理
种子经0.1%的H2O2消毒30 min后用蒸馏水漂洗并浸种24 h,转移到28 ℃、湿度为80%的培养箱中进行催芽,露白后将长势良好的幼苗转移到含有4 L 的全营养液[24]中进行水培育苗,待水稻长至两叶时转移到16 孔的塑料板上,将塑料板放入含10 L 全营养液的塑料桶中培养,每孔种2 株,每3 d 更换一次营养液。15 d后取长势一致的水稻,先用去离子水浸泡水稻根系3 次,再用0.05 mmol·L-1CaCl2浸泡30 min,然后用去离子水清洗3 次去除根表的CaCl2溶液后进行铁膜、磷和铝毒诱导,过程分为如下5组:(1)CK,全营养液(pH5.0)培养;(2)Al,50 μmol·L-1Al3+溶液(含0.05 mmol·L-1CaCl2,pH5.0)处理48 h;(3)IP+Al,40 mg·L-1Fe(Ⅱ)的全营养液(pH5.0)处理24 h 后,用去离子水清洗根部3 次,再用0.05 mmol·L-1CaCl2浸泡30 min,接着用去离子水清洗3次去除根表的CaCl2溶液,再全部转移到50 μmol·L-1Al3+溶液(含0.05 mmol·L-1CaCl2)中,处理48 h;(4)IP+Al+P15,40 mg·L-1Fe(Ⅱ)的全营养液处理24 h 诱导铁膜形成,经去离子水和0.05 mmol·L-1CaCl2溶液清洗完全后转移至15 mg·L-1磷水平的NaH2PO4溶液(pH5.0)中处理24 h,之后继续用去离子水和CaCl2溶液洗净根部,再进行50 μmol·L-1Al3+溶液(含0.05 mmol·L-1CaCl2)处理48 h;(5)IP+Al+P30,40 mg·L-1Fe(Ⅱ)的全营养液处理24 h诱导铁膜形成,经去离子水和CaCl2溶液清洗,再转移至30 mg·L-1磷的NaH2PO4溶液(pH5.0)中处理24 h,之后继续用去离子水和0.05 mmol·L-1CaCl2溶液洗净根部,再进行50 μmol·L-1Al3+溶液(含0.05 mmol·L-1CaCl2)处理48 h。Al3+均以AlCl3·6H2O 形态供应,Fe(Ⅱ)均以Fe(Ⅱ)-EDTA 形态供应,磷均以NaH2PO4·2H2O 形态供应,各处理溶液pH 每日均用1 mol·L-1NaOH 和1 mol·L-1HCl 调节至pH5.0,各处理组水稻总生长天数一致。
1.3 测定方法
1.3.1 水稻生物量测定
水稻收获后,用去离子水冲洗干净,分离地上部和根系两部分,地上部于105 ℃杀青30 min 后,放在70 ℃烘干至恒质量,称量即为地上部干质量。根系先用连二亚硫酸钠-柠檬酸钠-碳酸氢钠(DCB)浸提除去铁膜后用去离子水洗净,杀青烘干至恒质量,称量即为根系干质量,每个处理重复3次。
1.3.2 水稻根尖Al测定
每个处理选取30个1 cm 长的根尖经去离子水和0.05 mmol·L-1CaCl2清洗后置于1.5 mL 的离心管中,加入1 mL 2 mol·L-1HCl 溶液振荡浸提24 h。浸提液经ICP-AES测定Al含量。每个处理重复3次。
1.3.3 根表铁膜的Fe含量测定
水稻先用去离子水和0.05 mmol·L-1CaCl2去除根表残留的营养液后,选取每组中长势相同的3 株水稻根系,经DCB 处理提取铁膜,将去除铁膜后的3 株水稻根用去离子水反复冲洗干净后烘干,称量其干质量。采用ICP-AES测定DCB提取液中的Fe含量。用铁膜中的Fe含量表示铁膜含量,单位为g·kg-1DW。
1.3.4 根尖细胞壁多糖组分的测定
水稻先用去离子水和0.05 mmol·L-1CaCl2去除根表残留的营养液后,参照Zhong 等[25]的细胞壁组分的提取方法。取各组提取所得的上清液150 μL 进行细胞壁多糖组分测定,加入250 μL 冰乙酸中和后加入硫酸-硼砂2.4 mL,100 ℃下水浴5 min,在冰中立即冷却后加入40 μL 0.15%间羟基联苯,放置20 min 后在520 nm 下比色,标准空白以0.5% NaOH 代替间羟基联苯。果胶、半纤维素含量用糖醛酸含量表示。每个处理重复3次。
1.3.5 根尖酸性磷酸酶(APA)活性测定
水稻先用去离子水和0.05 mmol·L-1CaCl2去除根表残留的营养液后,参照秦丽凤APA 活性的测定方法,酶活性以单位时间内每克蛋白水解对硝基苯磷酸二钠(ρ-NPP)生成对硝基苯酚(NP)的量(gNP·g-1Pr·h-1)表示[26]。每个处理重复3次。
1.4 数据处理
根据1.3中各指标测定所得数据计算平均值和标准误差,采用Excel 2013 软件制图,并用SPSS 21.0 进行统计分析。
2 结果与分析
2.1 铁膜和磷对铝毒下水稻生物量的影响
Al 处理下菲优多系1 号和红良优166 地上部和根系生长受到显著抑制,根系干质量分别比对照下降了32.4%和35.8%,地上部干质量分别比对照下降了26.5%和37.9%,其中铝敏感基因型红良优166 降幅较大(图1)。IP+Al 处理的红良优166 根系和地上部干质量显著高于Al 处理组水稻。较IP+Al 处理,15 mg·L-1磷处理后的两基因型水稻的地上部和根系干质量变化差异不明显,而当30 mg·L-1磷处理时根系干质量达显著差异,分别增加了20.4%和22.6%,表明水稻根表铁膜形成后能有效缓解铝毒,较高浓度的外源磷处理覆铁膜水稻,能进一步降低Al 对水稻根系的损伤。
2.2 铁膜和磷对铝毒下水稻根尖Al含量的影响
单Al 处理下菲优多系1 号和红良优166 根尖Al含量明显增多,均比对照增加433.3%(图2)。IP+Al处理下菲优多系1 号和红良优166 的根尖Al 含量分别比单Al 处理降低了15.6%和33.3%,说明铁膜有效降低了水稻根尖Al 含量,这种改善的效果在敏感型水稻红良优166 中更明显。同时,随着外源磷处理浓度增加,菲优多系1号的根尖Al含量也逐渐降低,IP+Al+P30 处理时菲优多系1 号根尖Al 含量比IP+Al 处理降低55.6%,表明铁膜与外源磷的联合处理能有效降低根尖Al含量,该种作用对菲优多系1号更有效。
2.3 铁膜和磷对铝毒下水稻根表铁膜厚度的影响
外源Fe(Ⅱ)处理下,IP+Al、IP+Al+P15、IP+Al+P30 的水稻根表铁膜中的Fe 含量显著高于无Fe(Ⅱ)处理的CK 和Al 处理,与Al 处理相比菲优多系1 号和红良优166 各组的增幅达434.5%和488.8%、621.3%和265.5%、295.3%和344.3%,红良优166 根表的铁膜厚度高于菲优多系1号(图3)。表明Fe(Ⅱ)处理诱导根表形成大量铁膜。
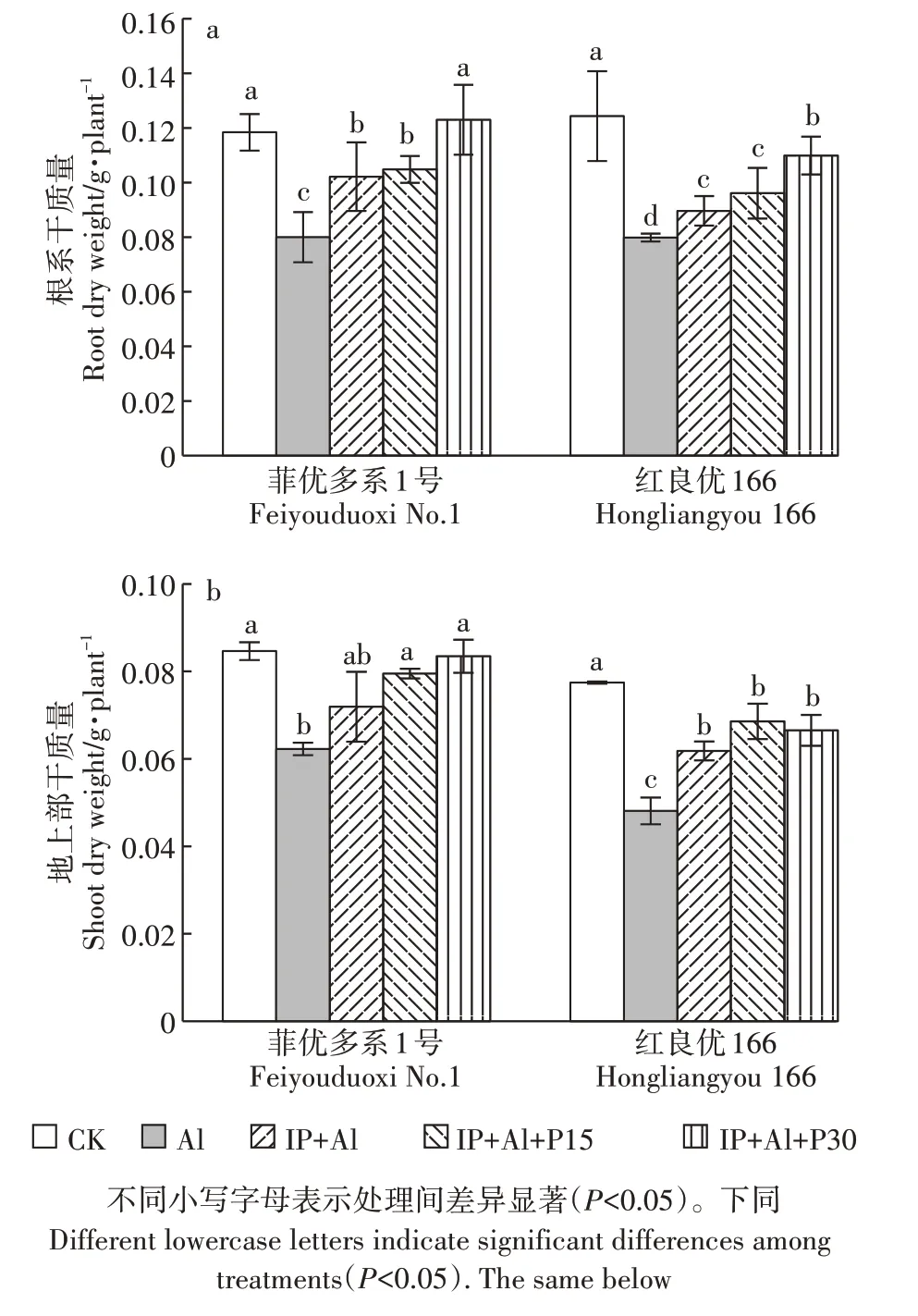
图1 铁膜和磷对铝毒下水稻根系和地上部干质量的影响Figure 1 Effects of iron plaque and P on dry weight of rice roots and shoots under aluminum toxicity
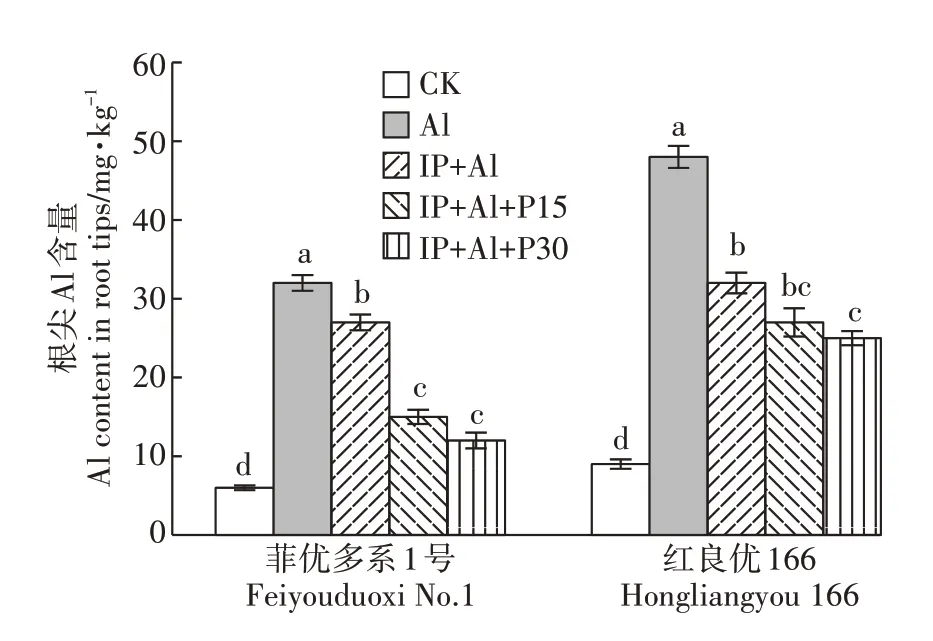
图2 铁膜和磷对铝毒下两个基因型水稻根尖Al含量的影响Figure 2 Effects of iron plaque and P on Al content in root tips of rice under aluminum toxicity
2.4 铁膜和磷对铝毒下水稻根尖果胶含量的影响
Al 处理下菲优多系1 号和红良优166 根尖细胞壁果胶含量分别比对照增加18.6%和27.8%(图4)。IP+Al处理时两基因型水稻根尖细胞壁果胶含量分别比Al 处理减少了22.6%和29.9%。随着外源磷的加入,两基因型水稻根尖细胞壁果胶含量均显著降低,与Al 处理组相比,菲优多系1 号和红良优166 在IP+Al+P15 处理下根尖果胶含量分别下降了33.7%和45.7%,在IP+Al+P30 处理下根尖果胶含量降幅达43.3%和57.8%,表明根表有铁膜存在时,水稻根尖的细胞壁果胶含量随着磷处理浓度增加而降低,水稻根尖果胶含量的变化同时说明了磷能够有效降低覆铁膜水稻根尖细胞壁果胶含量。
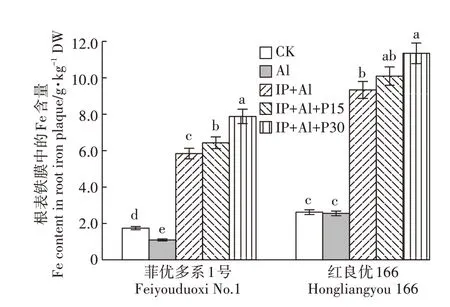
图3 铁膜和磷对铝毒下两个基因型水稻根表铁膜中Fe含量的影响Figure 3 Effect of Fe(Ⅱ)and P treatment on iron plaque formation on root surface of rice under aluminum toxicity
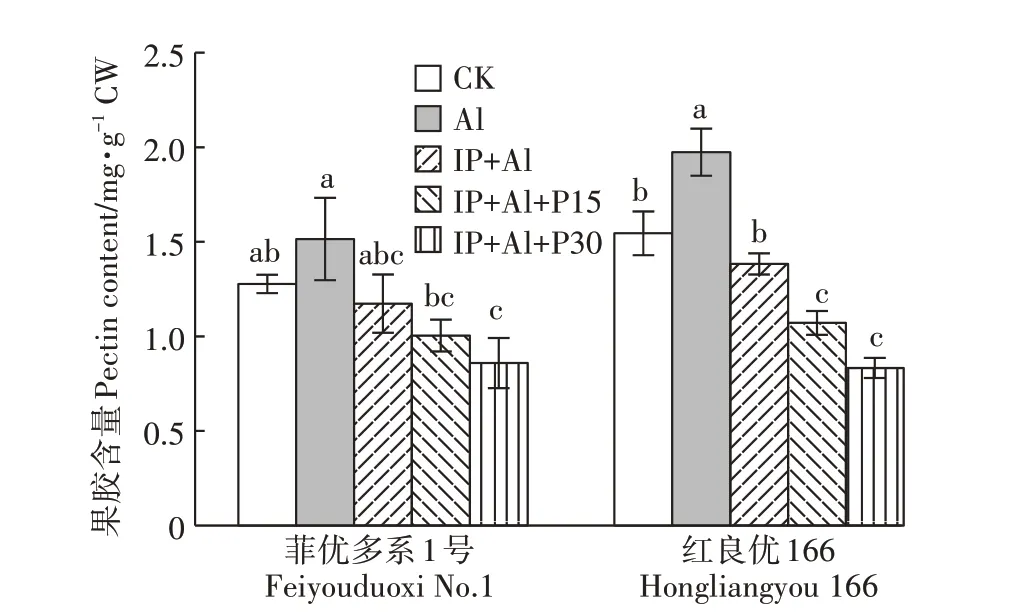
图4 铁膜和磷对铝毒下水稻根尖细胞壁果胶含量的影响Figure 4 Effects of iron plaque and P on pectin content in cell wall of root tips under aluminum toxicity
2.5 铁膜和磷对铝毒下水稻根尖半纤维素1含量的影响
与对照相比,单Al处理下菲优多系1号和红良优166 的半纤维素1 含量分别增加了66.1%和182.7%,红良优166 增幅远大于菲优多系1 号(图5)。IP+Al处理的根尖细胞壁半纤维素1 含量较Al 处理下降22.6%和29.9%,铁膜和磷共同处理下根尖细胞壁半纤维素1 含量比铁膜单独处理时降幅更大,其中IP+Al+P15 处理组降幅分别为21.0%和41.5%,IP+Al+P30 处理组降幅分别为20.5%和40.2%,表明铁膜能够降低Al处理下根尖细胞壁半纤维素1的产生,外源磷能进一步降低覆铁膜水稻半纤维素1的含量。
2.6 铁膜和磷对铝毒下水稻根尖半纤维素2含量的影响
根尖细胞壁半纤维素2含量变化与半纤维素1含量变化基本一致(图6)。单Al处理下,根尖细胞壁半纤维素2含量与对照相比显著上升,但与单Al处理相比,菲优多系1 号和红良优166 根尖细胞壁半纤维素2 含量在IP+Al 处理下分别下降了28.4%和28.1%,IP+Al+P15 处理降低42.7%和43.0%,IP+Al+P30 处理降低了44.8%和45.3%。表明根表铁膜和外源磷处理能有效降低Al诱导的细胞壁半纤维素2含量的增加。
2.7 铁膜和磷的作用对铝毒下水稻根尖APA 活性的影响
Al 处理48 h 后菲优多系1 号的APA 活性在各处理组均无显著性差异,IP+Al 处理96 h APA 的活性显著增加(图7),而红良优166 的IP+Al 处理组的APA活性在Al 处理48 h 和96 h 下均显示出明显的增强,表明低磷状态是水稻根系APA 活性急剧增加的主要原因,较短时间内覆铁膜的菲优多系1 号能更有效地吸收可溶性磷来对抗低磷环境,同时耐受型水稻显示出更强的抗低磷能力。
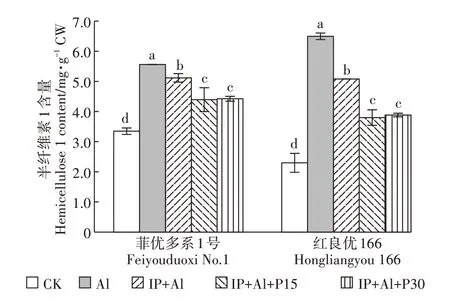
图5 铁膜和磷对铝毒下水稻根尖细胞壁半纤维素1含量的影响Figure 5 Effects of iron plaque and P on hemicellulose 1 content in cell wall of root tips under aluminum toxicity
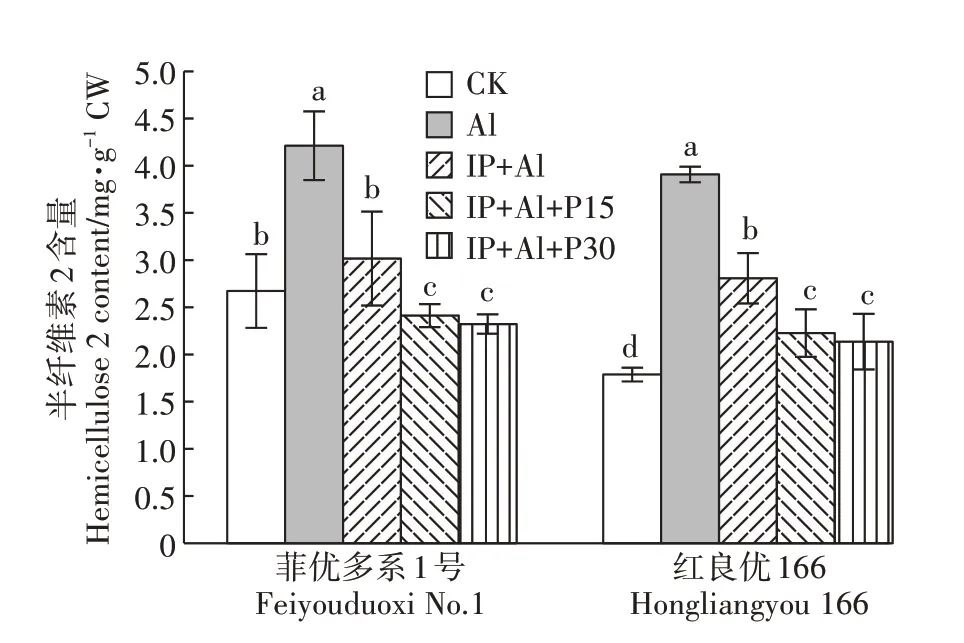
图6 铁膜和磷对铝毒下水稻根尖细胞壁半纤维素2含量的影响Figure 6 Effects of iron plaque and P on hemicellulose 2 content in cell wall of root tips under aluminum toxicity

图7 铁膜和磷对铝毒下水稻根尖APA含量的影响Figure 7 Effects of iron plaque and P on APA content in rice root tips under aluminum toxicity
3 讨论
铁膜能吸附Cd、As 等有害金属[27],有效缓解其对植物生长的损伤[28-29]。Chen 等[30]发现水稻根表铁膜可以将Al 吸附在根表,同时铁离子能预饱和水稻根尖Al 的结合位点,减轻根系对Al 的吸收。本实验中经Fe(Ⅱ)处理后水稻根表铁膜大量形成(图3),显著减少了根尖Al含量(图2),缓解了铝毒导致的水稻根系以及地上部生物量的降低(图1),说明铁膜的存在能有效阻止Al 进入水稻根尖,减缓铝毒对水稻生长的抑制作用。Dai等[31]在对红树林的研究中发现施加外源磷增强了铁膜对Cd 的吸附能力。因此,本实验采用覆铁膜水稻经不同浓度外源磷处理来分析铁膜对水稻耐铝的作用。结果表明外源磷处理明显降低铝耐受型水稻菲优多系1号的根尖Al含量,磷浓度达30 mg·L-1时,两基因型水稻根系干质量显著增加,菲优多系1 号的根尖Al含量进一步降低(见图1~图3),由此表明,外源磷降低了Al 在耐受型覆铁膜水稻根尖的积累,较高浓度的磷能缓解Al 对覆铁膜水稻根系生长的抑制,增强了水稻的耐铝效应。
水稻根表铁膜是一个磷储藏库,能从贫营养环境吸附磷从而满足植物生长的需要[32]。Batty 等[33]提出Al 能与磷酸盐共沉淀形成难溶的磷酸铝,Al 胁迫下铁膜的形成和磷处理后铁膜上吸附磷的增加为磷和Al 在铁膜表面的反应与沉积提供了条件,因此在菲优多系中观察到磷处理后根尖Al 含量的降低可能是由于铁膜上的磷酸盐和Al 反应形成稳定的铝磷酸盐络合物[34],而降低了根尖Al的积累。
细胞壁在植物应对铝毒时起着至关重要的作用。Yang等[35-36]发现在Al处理后,铝敏感水稻细胞壁中的果胶以及Al 含量显著高于耐铝水稻,在五加科植物三七[Panaxnotoginseng(Burk.)F.H.Chen]的根中,83%的Al 结合在根细胞壁的果胶上,Zhu 等[37]分析了CO2对铝毒作用下水稻细胞壁组分的影响,发现水稻培养环境中较高的CO2浓度诱导细胞壁中半纤维素含量降低从而缓解铝毒对水稻根系的损伤,表明细胞壁多糖含量与植物铝耐受性相关,通过外源介质调控细胞壁多糖组分含量能够缓解铝毒。磷处理显著降低了覆铁膜水稻菲优多系1 号的根系细胞壁多糖含量,与IP+Al 相比,IP+Al+P15 处理组的根尖细胞壁果胶含量降幅为33.7%(图3),半纤维素1、半纤维素2 含量的降幅分别为21.0%和41.5%(图4~图5),IP+Al+P30处理使菲优多系的根尖细胞壁果胶含量进一步减少,降幅达到43.3%,根系细胞壁多糖的含量变化直接影响了根系细胞的伸展[38],细胞壁多糖的增加引起细胞壁的硬化[39],从而抑制了根系的伸长。本试验中细胞壁多糖含量降低后根细胞的伸展性得到提高,因此磷缓解了铝毒对根系生长的抑制作用,增强覆铁膜水稻尤其是铝耐受型水稻的耐铝作用。细胞壁多糖是重金属离子的主要结合位点[5,10,35]。从细胞壁多糖的成分来看,果胶含量随着磷处理浓度的增加而降低。果胶富含同聚半乳糖醛酸,在其C6 位上的游离羧基能够与金属阳离子结合[10,40],其含量降低后Al在根尖的结合位点减少,从而缓解了铝毒对水稻根系的损伤。
此外,植物能引发一系列适应性反应来抵御低磷环境,其中包括APA 的诱导和分泌,APA 能将环境中植物无法吸收的有机磷转化为利于植物吸收的无机磷形态[41]。试验中发现菲优多系1 号在48 h 时各个处理组根系的APA 含量无显著差异,在96 h 时IP+Al+P0组的APA含量显著增加,而对于红良优166,处理48 h时IP+Al+P0组的APA 含量已经呈现出显著的差异(图7),表明磷在铝敏感型水稻抗铝过程中具有重要作用,在低磷环境中水稻通过分泌大量APA 来增加可用的磷,从而提高覆铁膜水稻的铝耐受性。
4 结论
(1)50 μmol·L-1Al 处理显著降低水稻根系和地上部干质量,增加根尖Al 含量和细胞壁果胶、半纤维素1以及半纤维素2含量。
(2)根表覆铁膜处理下,菲优多系1 号和红良优166 通过降低根尖细胞壁的果胶、半纤维素1 和半纤维素2 含量,减轻Al 对水稻生长的抑制,增加水稻根系和地上部干质量。
(3)覆铁膜水稻经磷处理,根尖细胞壁的果胶、半纤维素1 和半纤维素2 的含量显著降低,从而减少Al在根尖的沉积,减轻根生长受抑程度。
