干热河谷番茄苗期叶片光合效率的土壤水分阈值效应
2019-12-19李建查闫帮国潘志贤张雷岳学文何光熊樊博史亮涛方海东
李建查 闫帮国 潘志贤 张雷 岳学文 何光熊 樊博 史亮涛 方海东

摘 要 本文以番茄植株为供试材料,采用旱棚盆栽人工给水后自然耗水获得系列土壤水分梯度的方法,利用Li-6400光合测定系统测定不同水分条件下番茄光合生理特性。通過光合参数与土壤水分回归分析,结合数学模型求解,研究了番茄苗期叶片光合效率的土壤水分阈值效应。结果表明:番茄叶片净光合速率(Pn)、蒸腾速率(Tr)、水分利用效率(WUE)及光合参数具有明显的水分临界效应。Pn、Tr、WUE和气孔限制值(Ls)均随土壤水分的降低先升高后下降,但是各指标水分临界值表现不同步,其中Tr和Pn的水分饱和点分别为20.68%和18.18%,WUE最高点为15.83%,Ls最高点为10.74%。当土壤含水率低于10.74%,番茄茎和叶片SOD活性均显著提高。土壤含水率10.74%是番茄叶片光合结构损伤的临界点,也是干热河谷番茄苗期生长所允许的土壤水分最大亏缺程度。而土壤含水率15.83%~18.18%为番茄苗期高产高效水含量,此时番茄幼苗叶片具有较高的光合能力和高效生理用水特性,是确保干热河谷番茄高产高效节水管理的关键区间之一。
关键词 干热河谷;番茄;光合特性;土壤水分;阈值效应
中图分类号 Q945.79 文献标识码 A
Abstract Tomato plants were selected as the experimental material. Soil water gradients were obtained by the balances between water provision and consumption in experimental pots in a covered shed. The Li-6400 portable photosynthesis system was used to measure the photosynthetic efficiency parameters under different soil water conditions. It was found that the net photosynthetic rate (Pn), transpiration rate (Tr), water use efficiency (WUE) and photosynthetic parameters of tomato leaves had clear thresholds of soil moisture. Pn, Tr, WUE and stomatal limiting value (Ls) first increased and then decreased with decreasing soil water content, but their thresholds were different. The water saturation points of Pn and Tr were 18.18% and 20.68% respectively, the water efficiency point of WUE was 15.83%, and the maximum soil moisture of Ls was 10.74%. SOD activities of tomato roots and leaves increased significantly when soil water moisture was below 10.74%. The soil water of 10.74% was a critical point that the photosynthetic structure of tomato leaves was impaired, and it was also the maximum thershold point of soil water deficit allowed tomato seedling growth in the dry-hot valley. Soil moisture from 15.83% to 18.18% was classified as the intervals of high productivity and high efficiency; in this range, tomato had high photosynthetic capacity and efficient physiological characteristics for water consumption. This range of soil water meets to tomato production with high productivity and high efficiency in the dry-hot valley.
Keywords dry-hot valley; tomato; photosynthetic characters; soil moisture; threshold effect
DOI 10.3969/j.issn.1000-2561.2019.11.025
光合参数是反映植物对逆境生理过程响应的主要指标,通过研究植物光合作用与土壤水分的关系,有助于阐明植物在环境变化中的生理适应性[1-2]。土壤水分是影响植物生理过程和植物分布的重要生态因子,对植物光合作用和水分利用效率的影响较大[3-5]。根据生态学的限值因子法则,水分过多或过少均影响其光合特性,植物光合生理过程对土壤水分的需求应存在不同水平的阈值。近年来,国内外水分生理学研究主要涉及不同水分亏缺程度下植物生理生化因子的变化及其对水分胁迫的适应特征与机理等[2, 6-7]。
干热河谷高温少雨,蒸发量大,是我国西南地区典型的生态脆弱区。由于光热资源充足,云南元谋干热河谷具有“天然温室”之美称,是我国反季节蔬菜主要生产区之一,具有“金沙江边大菜园”之称。干热河谷水资源短缺,水成为限制该区农业发展的主要生态因子,导致干热河谷成为典型的无灌溉就无农业的灌溉农业区。发展高效节水灌溉农业是干热河谷农业可持续发展的重要战略措施,而研究灌溉土壤水分指标对制定区域高效节水灌溉制度和灌溉定额具有重要意义。番茄是元谋干热河谷主要蔬菜之一,现有研究主要集中在番茄对水分胁迫强度和胁迫时间的生理响应[8-9],缺乏对番茄光合生理特征在多级水分梯度下的连续性变化的研究,番茄生长与土壤水分的定量关系及其光合生理过程也不明确,导致番茄生产过程中灌溉制度的制定缺乏理论指导。为了揭示干热河谷生境番茄光合特性对土壤水分的响应规律,确定维持番茄较高光合能力和水分利用效率的水分阈值,本研究以番茄为试验材料,采用盆栽人工给水和自然耗水相结合的方法,测定不同水分梯度下番茄叶片光合参数及其水分阈值,评价土壤水分与番茄光合生理的关系,为干热河谷番茄水分管理提供理论依据。
1 材料与方法
1.1 试验区概况
云南元谋干热河谷地处滇中高原北部,东经101?35′~102?06′,北纬25?23′~26?06′,平均海拔1350 m。年均温21.9 ℃,无霜期305~331 d,年降雨量611.3 mm,蒸发量是降雨量的5~6倍。光热资源充足,年平均日照时数为7.3 h/d。土壤为砂壤土,土壤容重1.44 g/cm3,田间持水量为19.42%,pH 6.4,土壤有机碳0.61%,全氮0.05%,碱解氮39 mg/kg,全磷0.188 g/kg,有效磷30.38 mg/kg,全钾7.44 g/kg,速效钾129 mg/kg。
1.2 试验材料和设计
本研究于2016年10月7日—11月4日在云南省农业科学院热区生态农业研究所遮雨棚内开展。采用盆栽试验法,试验塑料盆长15 cm、宽15 cm、高15 cm。采集元谋干热河谷农田0~40 cm土层土壤,自然风干,过2 mm土样筛。然后称重装盆,每个盆装约1 kg土,并记录实际质量数据,共装15盆。从当地育苗公司购买品种名称为“拉比”的标准番茄苗,选择子叶完整、茎秆粗壮、叶片深绿、无病斑、无虫害、节间短、株高在16~18 cm、5~6片真叶的番茄幼苗移栽,每盆种植1株,进行正常水分管理。待番茄度过缓苗期,恢复正常生长时,选择长势一致的10盆番茄,其中5盆为对照组,正常浇水。另外5盆番茄一次性灌水至土壤水分饱和,第2天开始监测自然耗水过程,每天称重,获取土壤水分含量(W),并进行光合参数测定,获得系列水分梯度以及水分梯度下的光合特征值。直到植株萎蔫时停止,水分处理历时8 d。植株萎蔫时分别采集试验组和对照组的番茄根、茎和叶片测定各器官丙二醛含量和保护酶系统。番茄生育期纯氮施用量为9 g/株、P2O5 8 g/株、K2O 17 g/株,其中50%纯氮、60%P2O5、30%K2O作为底肥,苗期追施10%纯氮、12%P2O5、8%K2O,花果期追施40%纯氮、28%P2O5、62%K2O。
1.3 方法
光合参数的测定:利用Li-6400便携式光合仪对番茄的光合生理指标以及响应的生态环境要素进行测定,测定时间为9: 00—11: 00,测定每盆番茄新展开的叶子,从上往下数第3片叶子。光照强度控制为1500 μmol/(m2?s),仪器自动记录净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)等生理参数以及大气温度、湿度、光合辐射等生态环境因子。叶片水分利用效率WUE=Pn/Tr;气孔限制值Ls=1-Ci/Ca,其中Ci/Ca为胞间CO2浓度与环境CO2浓度的比值,由光合仪直接获取。
丙二醛含量和保护酶系统的测定:丙二醛(MDA),硫代巴比妥酸法;超氧化物歧化酶(SOD),氮蓝四唑法;过氧化物酶(POD),愈创木酚比色法;过氧化氢酶(CAT),紫外吸收法。
土壤水分含量测定采用称重法,土壤水分测定时间与光合作用观测时间一致。
1.4 土壤水分阈值分级
基于光合作用的土壤水分有效性分级,采用限值求解法[10]。主要依據光合生理指标与土壤水分含量之间的定量关系,通过数学模型求解光合参数的水分低限值和高限值,对土壤水分有效性进行划分。引入积分求解Pn和WUE的平均值,再用数学模型求解2个参数平均值对应的水分点作为中等光合效率和中等水分利用效率的分界值。将Pn和WUE获得最高、最低及平均值的土壤水分临界值作为土壤水分有效性分界点,结合Pn和WUE随土壤水分的响应规律,建立以Pn和WUE大小为标准的光合效率水分阈值分级。光合作用是作物产量形成的生理基础,因此,该分级标准采用番茄叶片Pn和WUE的土壤水分临界效应值,并赋予其明确生产意义,用Pn和WUE代替农业中的产和效的概念。
1.5 数据处理
利用SPSS 19.0软件对光合参数与土壤水分进行回归分析,通过数学模型求解,得最大净光合速率、蒸腾速率和水分利用效率等,并对水分胁迫下不同器官保护酶活性进行方差分析和显著性检验。
2 结果与分析
2.1 番茄叶片净光合速率和蒸腾速率的水分响应特征
适宜土壤水分条件下,番茄叶片Pn和Tr均先随着土壤水分含量增加而增加,先后达到最高水平后2者均下降(图1)。这表明净光合速率和蒸腾速率对土壤水分具有阈值效应。Pn(P<0.01)和Tr(P<0.01)对土壤水分的响应过程均符合二次方程模型,由模型可以求解Pn和Tr最大值时的土壤水分含量分别为18.18%和20.58%。Pn为0时对应的土壤水分为2.30%。根据拟合方程的积分式求得Pn平均值对应的土壤水分含量为27.05%和9.30%,Tr平均值对应的土壤水分含量为29.72%和11.64%。由此确定番茄叶片光合和蒸腾中等以上水平的土壤水分含量分别为9.30%~ 27.05%和11.64%~29.72%。这表明土壤水分含量过高或过低都影响番茄净光合速率和蒸腾速率,进而影响番茄产量。
2.2 番茄叶片水分利用效率的水分响应特征
番茄叶片WUE随土壤水分含量的增加先增加后降低,WUE对土壤水分的响应过程符合二次方程模型(P<0.01),由此模型可以求解WUE最大值时的土壤水分含量为15.93%。根据拟合方程的积分式求得WUE平均值对应的土壤水分含量分别为23.42%和8.58%,由此确定番茄叶片水分利用效率中等以上水平的土壤水分为8.58%~ 23.42%。表明低土壤水分和高土壤水分条件下番茄叶片水分利用效率都有降低趋势,造成农业水资源浪费。
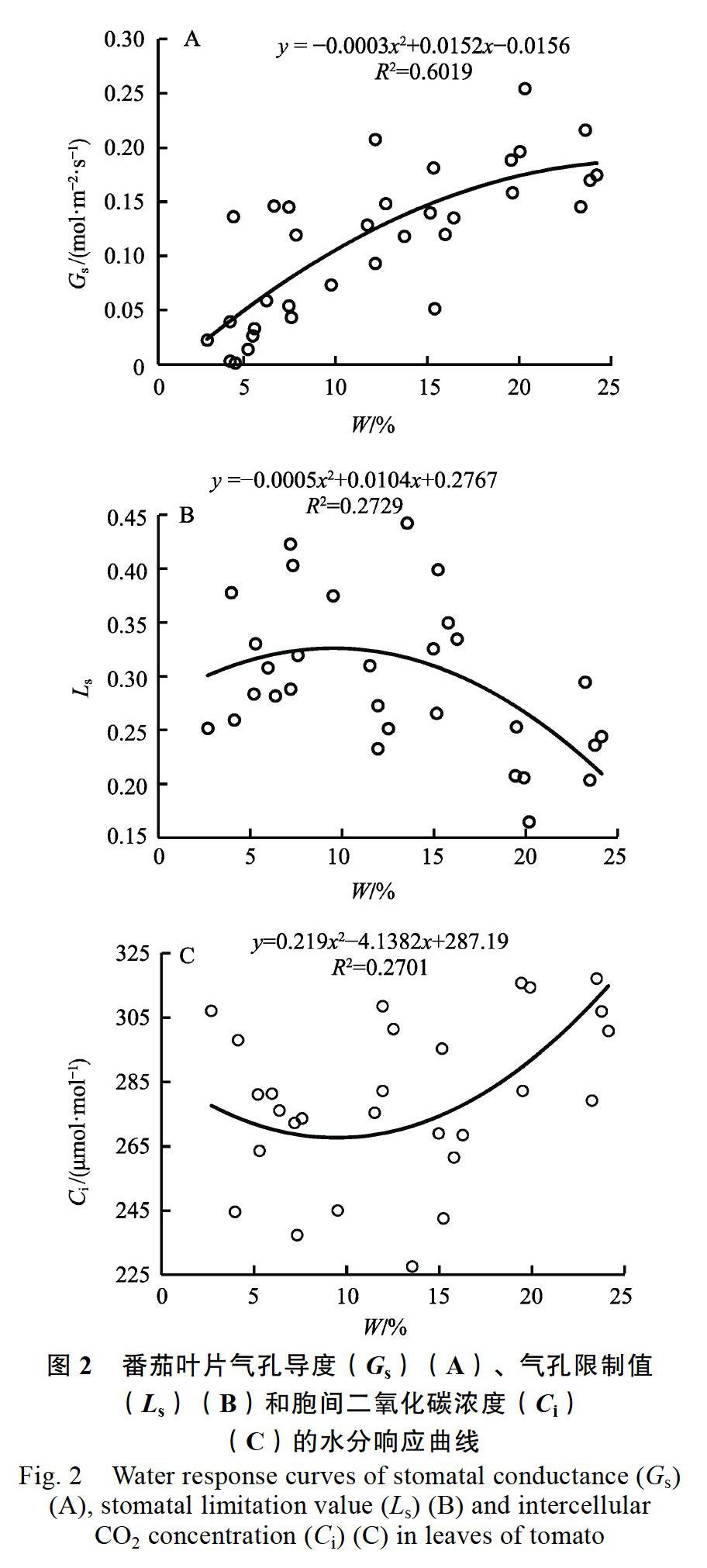
2.3 番茄叶片气孔导度、胞间CO2浓度和气孔限制值的水分响应特征
番茄叶片气孔导度Gs(P<0.01)、Ls(P<0.05)和Ci(P<0.05)的土壤水分响应过程均符合二次方程模型,Gs和Ls最大时土壤水分含量分别为25.33%和10.40%,Ci最低时土壤含水率为9.45%(图2)。土壤水分含量高于10.40%时,随土壤水分含量的增加,Gs和Ci表现为上升趋势,而Ls表现为下降趋势,即水分条件转好时番茄叶片净光合速率上升主要受气孔运动控制。而土壤水分含量低于10.40%时,随土壤水分含量降低,Gs和Ls表现为下降,而Ci表现为上升趋势,此时番茄叶片净光合速率下降主要由非气孔限制引起。在实测值土壤水分含量为10.74%时,出现Ci最低值和Ls最高值,这与模型计算值10.40%很接近,即土壤水分含量为10.74%是干热河谷番茄叶片净光合速率下降由气孔限制转为非气孔限制的转折点。
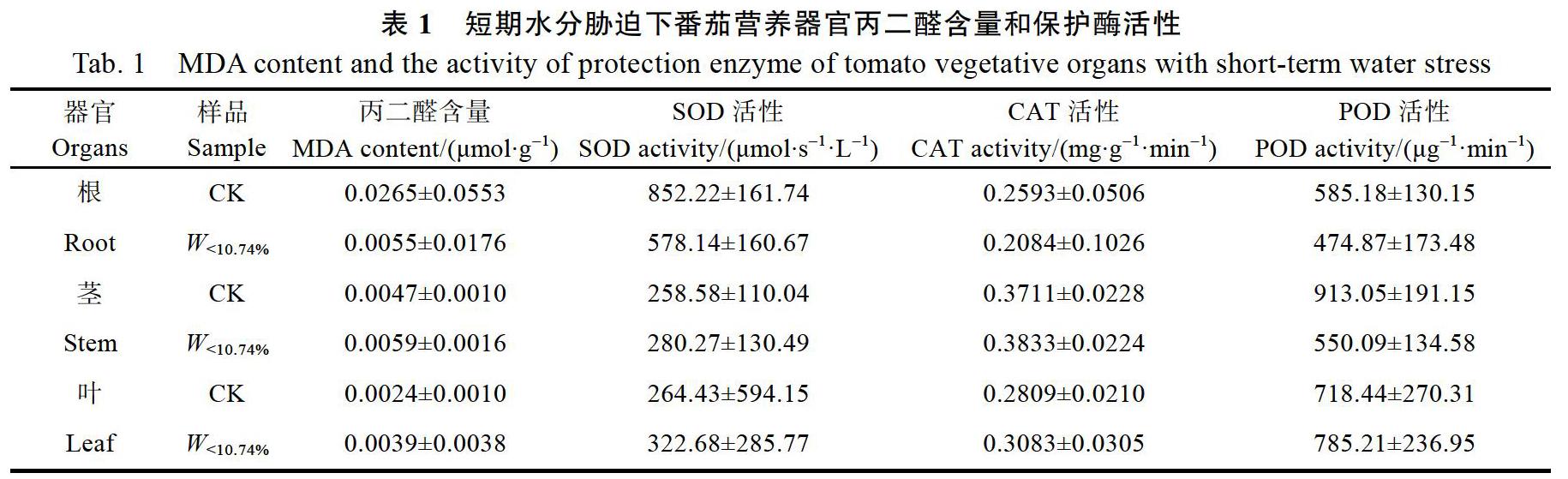
2.4 番茄各器官保护酶系统对短期水分胁迫的响应特征
土壤水分持续降低过程中,土壤水分含量由光合气孔限制转折点10.74%逐渐降至4.24%,此时番茄植株发生萎蔫,历时3 d。对土壤水分含量降至光合作用非气孔限制点之后番茄植株抗氧化系统变化特征深入分析结果显示,与对照组(CK)植株相比,萎蔫时(W<10.74%)植株各器官保护酶系统发生不同程度的变化(表1)。萎蔫时番茄根系的丙二醛含量、SOD活性、CAT活性和POD活性均降低,但变化趋势不显著,短期低土壤水分胁迫对番茄根系的保护酶系统没有产生明显影响。茎除了POD活性降低外,其他保护酶系统指标均升高,其中SOD活性(P<0.05)显著提高8.4%。叶片的丙二醛含量、SOD活性、CAT活性和POD活性均升高,其中SOD(P<0.05)显著提高22%。
2.5 番茄叶片光合效率的水分阈值分级
土壤水分低于2.30%时,Pn为0,不能形成产量,可以描述为无产无效水。高产高效水是指较高的Pn和WUE对应的土壤水分,Pn和WUE最大值时的土壤水分含量分别为18.18%和15.83%,土壤水分含量在15.83%~18.18%范围内可以描述为高产高效水。中产中效水主要依据Pn和WUE与土壤水分之间的积分式求解得2个参数的平均值来确定的,土壤水分含量在9.30%~15.93%和18.18%~23.43%范围内可以描述为中产中效水。低产低效水以低于Pn和WUE平均值对应的土壤水分来确定,土壤水分含量在2.30%~8.58%范围内可以描述为低产低效水。
3 讨论
3.1 光合效率的土壤水分阈值效应
水分胁迫对植物生长发育及光合作用影响较大,适宜的土壤水分条件下植物获得最大光合效率,而土壤水分含量过高或者过低都会降低植物光合作用效率[11-12]。番茄叶片光合参数对土壤水分表现出明显的临界效应,随着土壤水分含量减少,Gs降低,限制了CO2进入叶片组织,表现为Ci降低,Ls上升,Pn和Tr也随之降低,番茄叶片采取关闭气孔的方式来减少蒸腾失水[13]。当10.74% 干旱胁迫诱发植物体内产生更多活性氧,导致膜脂过氧化,造成氧化伤害[14]。而植物体内活性氧自由基代谢失调而引发的生物膜结构与功能的破坏,是光合作用非气孔限制的主要原因[15]。因此,深入分析土壤水分含量降至光合非气孔限制点之后番茄植株抗氧化系统变化特征,可以初步探讨番茄叶片光合作用非气孔限制的生理机制。当土壤水分含量由番茄叶片光合非气孔限制临界点10.74%逐渐降至萎蔫点4.24%时,番茄根系的丙二醛含量、SOD活性、CAT活性和POD活性变化均不显著。短期的水分胁迫下,茎和叶MDA含量增加,差异不显著,且茎和叶片SOD活性均显著提高,分别提高8.4%和22%。这与安玉艳等[16]研究结果一致。这表明土壤含水率10.74%是干热河谷番茄苗期叶片光合结构损伤的临界点,是干热河谷番茄生长所允许的土壤水分最大虧缺程度,土壤长期处于此水分临界值以下,膜系统容易遭受损伤,光合等生理过程受到明显伤害。本研究还发现根系忍耐干旱胁迫的能力较强[17-18],叶片最先呈现保护酶系统显著变化,茎的保护酶系统对土壤水分亏缺的响应程度介于根系和叶片之间。 Gs、Tr、Pn和WUE均随着土壤水分含量增加先增加后降低,这与郎莹等[19]研究结果一致,这表明净光合速率、蒸腾速率和水分利用效率对土壤水分具有阈值效应。Gs、Tr、Pn和WUE最大值时的土壤水分含量分别为25.33%、20.68%、18.18%和15.83%,随着土壤水分含量增加,番茄叶片最先达到WUE最大值,随后Pn达到最大值,此时Gs和Tr都处于上升状态,随后Tr达到最高水平,最后Gs也达到最大值,这与裴斌等[20]研究结果一致。这表明随着干旱的发生发展,气孔导度、蒸腾速率、光合速率和水分利用效率依次受到影响,而调节气孔导度是优化光合与蒸腾关系以提高植物生理用水效率的重要策略。光合与蒸腾呈现非线性关系,光合速率最高时的蒸腾速率为临界值,超出该值即为奢侈蒸腾[21],当土壤水分含量过高时,植物叶片进行奢侈蒸腾[22],造成农业水资源浪费。有研究表明,气孔导度受根系吸水速率与叶片蒸腾速率平衡的反馈制约,土壤水分含量成为影响叶片气孔导度的主导因子[23],通过适度调控土壤水分,抑制奢侈蒸腾并不影响光合生产,还能提高水分利用效率。因此,控制土壤水分含量不超过18.18%,有效控制番茄叶片奢侈蒸腾,提高单位蒸腾水的番茄产量。在一定土壤水分条件下,植物叶片还能通过牺牲部分光合能力以提高水分利用效率[24],导致植物水分利用效率最高时土壤水分含量为15.83%,低于光合速率最高点。这表明15.83%~18.18%为干热河谷番茄苗期高效节水管理适宜土壤水分含量参考值。气孔是植物控制叶片与大气之间碳水交换的重要门户[25],植物通过调节气孔导度来平衡、优化光合作用和蒸腾速率的关系[26-28]。当土壤水分含量高于18.18%时,随着土壤水分持续增加,净光合速率降低。植物可能为了获得生长所需碳而有限地消耗土壤水分,使叶片以有限的水分散失来获得最大的CO2同化量,表现出气孔最优行为[13],致使气孔逐渐关闭以降低蒸腾失水,优化光合作用和蒸腾速率的关系。气孔关闭减少了CO2的吸收,光合能力随之降低,但是番茄叶片降低Gs和Tr水分散失的同时降低Ci加强CO2同化,致使WUE达到最大值。 3.2 光合效率的土壤水分有效性分级及评价 依据维持Pn和WUE低限值、高限值和中等值进行光合效率水分阈值分级与评价。适宜土壤水分含量能够提高植物光合速率和水分利用效率,黄土丘陵区土壤含水率在12.9%~20.5%和12.8%~19.5%时分别为山杏[5]和黄刺玫[1]“高产高效水”,黄河三角洲贝壳堤岛旱柳“高产高效水”为16.5%~14.1%[8]。干热河谷区土壤水分低于2.30%时为番茄无产无效水,在2.30%~8.58%时为低产低效水,在9.30%~15.93%和18.18%~23.43%时为中产中效水,在15.83%~18.18%时为高产高效水,此时番茄叶片获得较高的光合能力,奢侈蒸腾受抑制,生理用水得到提高。与黄土丘陵区相比,元谋干热河谷番茄高产高效水范围较小;而与黄河三角洲相比,其范围较大,这可能与番茄需水特性有关,也可能与干热河谷土壤持水性有关。元谋干热河谷土壤含水率12.69%~13.88%为甜玉米高产高效水[29],明显低于番茄高产高效水范围,这主要与番茄的需水量较大有关。有研究表明,土壤水分含量在15.6%~20.4%时温室番茄光合速率和水分利用效率较高[30],此番茄光合适宜土壤水分上限值较本研究结果高,这可能是因为温室内CO2浓度较高,提高番茄最大CO2同化量,根据气孔最优行为调控气孔导度,从而提高光合适宜土壤水分上限。因此,深入研究不同干旱区不同植物土壤水分阈值对当地农林业水分管理具有重要意义。 参考文献 [1] 张淑勇, 夏江宝, 张光灿, 等. 黄刺玫叶片光合生理参数的土壤水分阈值响应及其生产力分级[J]. 生态学报, 2014, 34(10): 2519-2528. [2] 高 丽, 杨 劼, 刘瑞香. 不同土壤水分条件下中国沙棘雌雄株光合作用、蒸腾作用及水分利用效率特征[J]. 生态学报, 2009, 29(11): 6025-6034. [3] Zhang G C, Xia J B, Sha H B, et al. Grading woodland soil water productivity and soil bioavailability in the semi-arid Loess Plateau of China[J]. Clean-Soil, Air, Water, 2012, 40(2): 148-153. [4] Sofo A, Dichio B, Montanaro G, et al. Photosynthetic performance and light response of two olive cultivars under different water and light regimes[J]. Photosynthetica, 2009, 47(4): 602-608. [5] 夏江宝, 张光灿, 孙景宽, 等. 山杏叶片光合生理参数对土壤水分和光照强度的阈值效应[J]. 植物生态学报, 2011, 35(3): 322-329. [6] 杨 劼, 李国强, 曹 云. 皇甫川流域中国沙棘光合特征分析[J]. 水土保持学报, 2004, 18(2): 148-151. [7] 韩蕊莲, 梁宗锁, 侯庆春, 等. 黄土高原适生树种苗木的耗水特性[J]. 应用生态学报, 1994, 5(2): 210-213. [8] 郝舒雪, 曹红霞, 王虎兵, 等. 水分胁迫对番茄幼苗生理特性的影响[J]. 排灌机械工程学报, 2018, 36(10): 943-947. [9] 张纪涛, 马大炜. 番茄幼苗对水分胁迫的生理响应[J]. 北方园艺, 2015, 43(7): 46-49. [10] 夏江宝, 张淑勇, 赵自国, 等. 贝壳堤岛旱柳光合效率的土壤水分临界效应及其閾值分级[J]. 植物生态学报, 2013, 37(9): 851-860. [11] 张淑勇, 夏江宝, 张光灿, 等. 黄河三角洲贝壳堤岛叶底珠叶光合作用对CO2浓度及土壤水分的响应[J]. 生态学报, 2014, 34(8): 1937-1945. [12] Xiao J B, Zhang S Y, Zhang G C, et al. Critical response of photosynthetic efficiency in Campsis radicans (L.) Seem to soil water and light intensities[J]. African Journal of Biotechnology, 2011, 10(77): 17748-17754. [13] Farquhar G D, Sharkey T D. Stomatal conductance and photosynthesis[J]. Annual Review of Plant Physiology, 1982, 33(1): 317-345. [14] Montanaro G, Dichio B, Xiloyannis C. Shade mitigates photoinhibition and enhances water use efficiency in kiwifruit under drought[J]. Photosynthetica, 2009, 47(3): 363-371. [15] 张仁和, 郑友军, 马国胜, 等. 干旱胁迫对玉米苗期叶片光合作用和保护酶的影响[J]. 生态学报, 2011, 31(5): 1303-1311. [16] 安玉艳, 郝文芳, 龚春梅, 等. 干旱-复水处理对杠柳幼苗光合作用及活性氧代谢的影响[J]. 应用生态学报, 2010, 21(12): 3047-3055. [17] Selote D S, Khanna-Chopra R. Antioxidant response of wheat roots to drought acclimation[J]. Protoplasma, 2010, 245(1-4): 153-163. [18] Lee B R, Li L S, Jung W J, et al. Water deficit-induced oxidative stress and the activation of antioxidant enzymes in white clover leaves[J]. Biologia Plantarum, 2009, 53(3): 505-510 [19] 郎 莹, 汪 明. 春、夏季土壤水分对连翘光合作用的影响[J]. 生态学报, 2015, 35(9): 3043-3051. [20] 裴 斌, 张光灿, 张淑勇, 等. 土壤干旱胁迫对沙棘叶片光合作用和抗氧化酶活性的影响[J]. 生态学报, 2013, 33(5): 1386-1396. [21] 王会肖, 刘昌明. 作物光合、蒸腾与水分高效利用的试验研究[J]. 应用生态学报, 2003, 14(10): 1632-1636. [22] 李茂松. 作物奢侈蒸腾及其调控基础研究[D]. 北京: 中国农业科学院, 2010. [23] 左应梅, 陈秋波, 邓权权, 等. 土壤水分、光照和空气湿度对木薯气孔导度的影响[J]. 生态学杂志, 2011, 30(4): 689-693. [24] 王海珍, 韩 路, 徐雅丽, 等. 土壤水分梯度对灰胡杨光合作用与抗逆性的影响[J]. 生态学报, 2017, 37(2): 432-442. [25] Hetherington A M, Wondward F I. The role of stomata in sensing and driving environmental change[J]. Nature, 2003, 424(6951): 901-908. [26] 范嘉智, 王 丹, 胡亚林, 等. 最优气孔行为理论和气孔导度模拟[J]. 植物生态学报, 2016, 40(6): 631-642. [27] 孙安安, 智颖飙, 姜平平, 等. 西鄂尔多斯四种荒漠植物光合作用特征与差异性[J]. 生态学报, 2019, 39(13): 4944-4952. [28] 侯慧芝, 张绪成, 尹嘉德, 等. 覆盖对西北旱地春小麦旗叶光合特性和水分利用的调控[J]. 应用生态学报, 2019, 30(3): 931-940. [29] 李建查, 孫 毅, 赵 广, 等.干热河谷不同土壤水分下甜玉米灌浆期光合作用光响应特征[J]. 热带作物学报, 2018, 39(11): 2169-2175. [30] 王晓森, 常 晓, 孟兆江, 等. 不同灌水下限与底肥施用量对温室番茄光合特性、产量和品质的影响[J]. 灌溉排水学报, 2016, 35(3): 45-50.
