膳食多酚的糖稳态调节作用
2019-12-19龙晓珊邹宇晓胡腾根黎尔纳沈维治凡U
王 晨 龙晓珊 - 邹宇晓 - 李 倩 胡腾根 - 黎尔纳 - 沈维治 - 刘 凡U
(1. 广东省农业科学院蚕业与农产品加工研究所,广东 广州 510610;2. 广东省农业科学院农业部功能食品重点实验室,广东 广州 510610;3. 广东省农产品加工重点实验室,广东 广州 510610)
机体糖稳态是生命体维持正常新陈代谢活动的重要基础,糖稳态失衡会增加糖尿病、肥胖和代谢综合症等代谢性疾病的发生或发展风险,控制糖稳态一直是该类疾病的重要防治策略。因此,通过“一日三餐”补充富含天然降糖因子的膳食干预方式对于控制糖代谢稳态具有十分重要的意义。多酚作为一类重要的植物次生代谢物,广泛存在于水果、蔬菜、葡萄酒、茶、咖啡等植物性食物中[1],大量源于动物和人体的研究结果显示膳食多酚在维持机体糖代谢稳态的过程中扮演了重要角色[2]。文章在查阅文献的基础上,综述膳食多酚种类和分布、调节糖代谢功效及作用机制,旨在为利用膳食多酚开发天然辅助降血糖保健食品提供理论依据。
1 膳食多酚的种类和分布
多酚类化合物的基本化学结构特征是苯酚与一个或多个羟基基团相连形成的共轭结构(图1)。已经报道的膳食多酚类化合物种类繁多,从简单的酚酸到分子量>30 kDa的高度聚合化合物有数百种[3]。多酚类化合物按化学结构的差异可分为酚酸类、黄酮类、二苯乙烯、香豆素、单宁5大类(图2)[4]。
咖啡酸和阿魏酸是常见的酚酸类化合物的代表性单体,咖啡酸主要分布于诸多蔬菜和水果中,是咖啡的主要成分,阿魏酸则多见于谷物外壳中,交叉结合在细胞壁的多聚糖上,影响细胞壁的完整性。黄酮类化合物的结构特征是2个芳香环和3个碳原子结合在一起形成的含氧杂环,根据杂环的类型又可分为黄酮、黄酮醇、黄烷醇、黄烷酮、异黄酮和花青素6类。最常见的黄酮是属于黄酮醇类槲皮素,在洋葱、茶叶、苹果中含量丰富;异黄酮类主要分布于豆类及其制品中,如染料木黄酮、黄豆苷元,被称为植物雌激素[5]。黄烷醇类的儿茶素广泛存在于茶叶和水果中;橙色素则是一种存在于柑橘中的黄烷酮类;矢车菊素、天竺葵素、飞燕草素等是常见的花色素单体,常与糖类分子结合形成花色苷,赋予桑椹、黑醋栗、蓝莓和覆盆子鲜艳的色泽。
图1 儿茶素和羟基苯甲酸的化学结构式
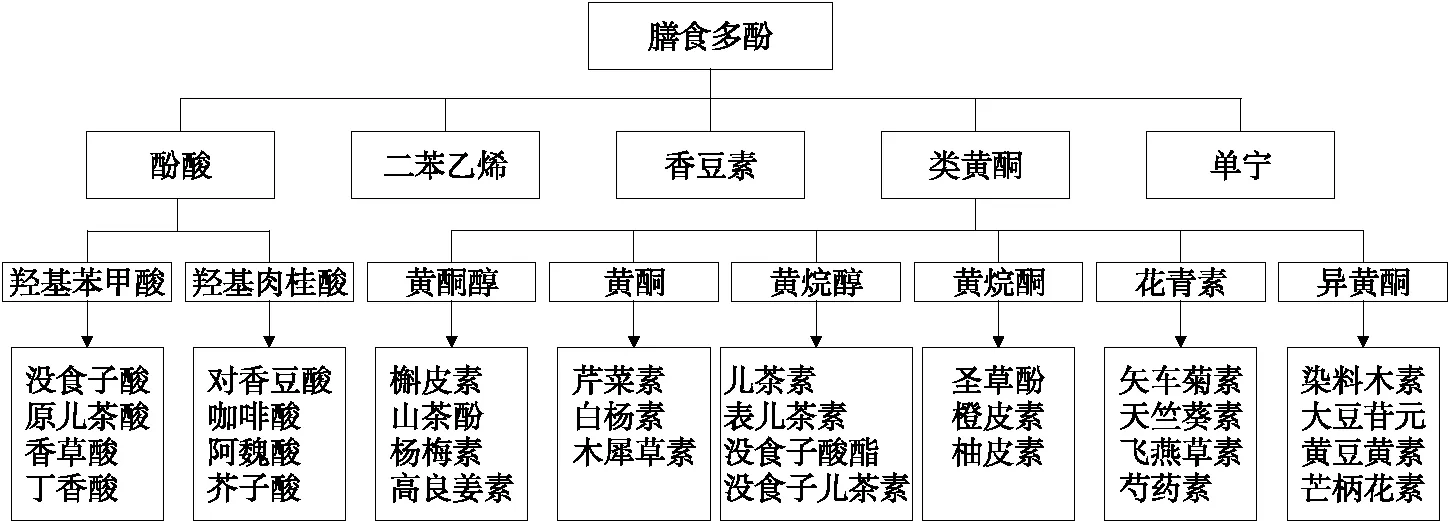
图2 膳食多酚的分类[4]
在果蔬中的多酚物质主要包括黄酮类、酚酸、单宁3大类。不同种类的植物其酚类物质组成千差万别,但种属分类接近的植物其酚类组成相似度更高,如表儿茶素和儿茶素等缩合单宁常见于葡萄和蓝莓等浆果中[6]。橙皮苷和柚皮苷两种重要的黄烷酮类物质主要分布在柑橘柚类果实中[7]。谷物中常见的酚类物质包括酚酸和黄酮,其中阿魏酸和p-香豆酸是分布最广泛的单体酚酸[8]。茶多酚是近年来研究食品领域的研究热点,是一类有代表性的多酚混合物,主要包括黄烷醇(儿茶素为主)、花色素(茶黄素、茶红素等)、黄酮及黄酮醇、酚酸和缩合酚酸5大类。其中儿茶素占茶多酚总量的60%~80%,包括表儿茶素(EC)、表没食子儿茶素(EGC)、表儿茶素没食子酸酯(ECG)、表没食子儿茶素没食子酸酯(EGCG)[9]。
在植物的萌芽、生长及加工过程中多酚的组成亦会发生复杂的变化。成熟的苹果主要含绿原酸、儿茶素、原花青素等,而未成熟的苹果还含有较多的二羟查耳酮、槲皮酮等化合物[10]。这是由于苹果成熟过程中植物激素水平和多酚合成、转化或代谢酶的水平变化共同造成的。发芽是谷物的一项重要生理活动,在此过程中由于多种酶的活性变化导致其酚类物质组成发生改变,在中国野生稻(菰)的发芽试验中,通过KEGG数据库分析探明了调控多酚代谢关键酶表达变化,发现发芽120 h时4种苯丙氨酸脱氨酶、1种4-香豆酸辅酶A连接酶、1种肉桂酰辅酶A还原酶、2种肉桂醇脱氢酶的表达水平远高于发芽36 h,由此促进了香豆酸和阿魏酸在120 h的高水平表达[11]。茶叶的加工方式是影响其酚类物质组成的重要原因。根据茶叶是否发酵及发酵工艺的差异,中国茶叶分为绿茶、黄茶、乌龙茶、红茶、白茶、黑茶6大茶系,茶多酚含量与发酵程度呈显著负相关。高海荣等[12]报道了中国16种常见茶叶中茶多酚含量的测定结果,发现样品间多酚含量差异极大(91.59~376.60 mg/g),分析可能是茶叶植物品种与加工工艺的差异造成的,从茶叶品种来说多酚含量差异为绿茶>白茶>黄茶>青茶(乌龙茶)>红茶>黑茶,就加工工艺而言多酚含量差异为未发酵茶>半发酵茶>发酵茶。由此可见,茶叶不发酵或发酵程度越轻,其多酚含量越高。
总体来说,膳食多酚组成的差异,一方面取决于基因等内源性的因素,另一方面植物在生长和萌发等生理活动的过程中,伴随着体内复杂的激素和代谢酶的变化,酚类组成也会随之改变。在食物加工过程中外界理化因子如温度、光线、微生物发酵等,也会直接影响植物的酚类组成,因此在研究植物多酚的健康效应时,应该关注内外因素对酚类成分的影响规律,提出适宜的原料采收期和加工方式。
2 膳食多酚对调节糖代谢功效
近年来,随着膳食多酚化学结构和生物活性研究的不断深入,越来越多源于动物和人体试验[13-14]结果显示,膳食多酚以多种方式参与机体的葡萄糖吸收与代谢,是一种极有潜力的糖稳态调节功能因子。针对糖尿病、肥胖等多种方式诱发的糖稳态失衡状态,增加膳食多酚摄入均显示出良好的改善效果。
2.1 膳食多酚对肥胖诱发的糖稳态失衡的调节作用
自1999年世界卫生组织(WHO)正式宣布肥胖是一种疾病以来,其相关的发病机理和防治方法就已成食品和医学领域科学研究的重要方向。肥胖患者通常都伴随糖代谢紊乱,通过饮食干预纠正糖代谢紊乱已经成为控制肥胖的有效措施。长期高脂膳食的摄入亦会导致机体的糖稳态失衡。白藜芦醇(30 mg/kg·BW)及其衍生物3′-羟基白藜芦醇(14 mg/kg·BW)可显著降低高脂饲料喂养的C57Bl/6小鼠的餐后血糖水平,效果与抗糖尿病药物阿卡波糖接近[13]。一项以28名肥胖患者为研究对象的随机双盲试验[14]结果显示,经过35 d日本虎杖提取物(富含白藜芦醇酚类成分)膳食干预,与安慰剂组相比肥胖患者的2 h糖耐量曲线下面积显著减少,胰岛素敏感性显著提高,糖稳态调节能力显著增强。这些源于动物和人体的试验结果表明,肥胖人群膳食中适当补充酚类物质,有助于改善肥胖导致的糖代谢紊乱,以及由此引发的胰岛素抵抗、糖耐量受损等,对于机体保持糖稳态具有十分重要的意义。
2.2 膳食多酚对糖尿病伴随糖稳态失衡的调节作用
糖尿病是以持续高血糖为病理特征的慢性代谢性疾病,病程长且并发症多,目前中国的发病率已经超过10%,因长期服用西药带来的器官损伤和低血糖等毒副作用是科学家们不得不面对的棘手问题,越来越多源于临床研究的证据[15-16]表明,增加膳食中多酚的摄入比例或许是解决这一问题的有效办法。连续两周给予糖尿病大鼠150,300 mg/g赶黄草多酚提取物,可显著降低大鼠血清中糖化血红蛋白A1C(HbA1c)的水平并升高胰岛素的水平,在口服淀粉耐量试验中发现600 mg/kg多酚提取物可将大鼠餐后血糖含量降低42%[17]。通过体内外模型结合并相互验证的研究方法,发现苹果[18]、藤茶[19]、罗汉果[20]和乌梅[21]等植物的酚类提取物显示良好的α-葡萄糖苷酶抑制活性,且有利于维持动物血糖的稳态化。
大量研究[22-24]结果显示,膳食多酚的摄入量与糖尿病发病风险呈负相关,但其有效性与多酚来源及单体组成有密切关系。以41 334位男性为研究对象的健康专业随访研究[22]结果表明,多酚的摄入量与2型糖尿病的发病风险呈显著负相关性,这项研究为膳食多酚预防2型糖尿病提供了直接的依据。此外,科学家们还开展了多酚种类及单体与2型糖尿病发病风险的研究。Hui等[23]研究结果表明,膳食多酚摄入水平高的人群发生2型糖尿病的风险显著降低(RR:0.89,95% CI:0.82~0.96),呈显著的量效关系,而且发现摄入类黄酮种类差异亦会导致2型糖尿病发病风险的差异,其中效果由高到低的类黄酮种类依次为:花色苷>黄烷-3-醇>黄酮醇>异黄酮。多酚单体的差异亦会导致2型糖尿病发病风险降低的差异。一项以1 111例女性为研究对象前瞻性研究[24]报道了膳食多酚摄入量与糖尿病发病风险的密切关系,通过比较调查对象尿液中的酚类物质组成,发现在早期随访期间(样本采集后4.6年),黄烷酮(柚皮素和橙皮素)和黄酮醇(槲皮素和异鼠李素)以及酚酸(咖啡酸) 可将T2DM风险降低39%~48%,进一步证实了多酚单体结构的差异与其降低糖尿病发病风险能力强弱存在密切关联。
随着肥胖、糖尿病等慢性疾病发病率的逐年增高,由此引发的社会健康问题受到普遍关注,通过日常膳食干预纠正肥胖、糖尿病引发的糖代谢紊乱是重要的防治策略之一。膳食多酚分布广泛且种类繁多,在不同食物中其组成和含量均不相同,综合已报道的动物试验和人体试验研究结果,发现多酚的种类及组成与其生物活性功能密切相关,它们以不同的方式参与到机体的糖稳态调节作用中,作用靶点和机制各不相同,从侧面说明构效关系是多酚生物活性研究的重要内容,如何确定活性更好的多酚组分乃至单体,都需要经过系统的体内外生物活性评价数据加以支持。
3 膳食多酚影响糖代谢的机制探讨
食物中的碳水化合物对机体发挥健康效应需经过消化、转运吸收和代谢等多个环节,其间涉及到诸多酶、激素、细胞因子及基因的复杂调控,综观前人的研究[25-26]结果,膳食多酚维持机体糖稳态的活性机制主要表现在抑制碳水化合物消化酶的活性和小肠葡萄糖转运蛋白表达、改善胰岛素抵抗等方面。
3.1 抑制碳水化合物消化酶的活性
α-葡萄糖苷酶是一类位于人体小肠黏膜刷状缘的酶,包括麦芽糖酶、蔗糖酶、乳糖酶等,进入人体的碳水化合物必须在消化道内经α-葡萄糖苷酶水解产生葡萄糖、果糖、半乳糖才能被人体吸收[27-28]。抑制α-葡萄糖苷酶的活性可有效抑制人体对葡萄糖的吸收速率,降低餐后高血糖,因而成为控制餐后血糖的重要靶点,阿卡波糖和伏格列多糖等均是基于该靶点开发的糖尿病治疗药物。
在一些体外化学模拟、细胞和体内试验[29-32]均报道了膳食多酚及其组成单体都具有良好的α-葡萄糖苷酶抑制活性。梁洁等[33]发现龙眼叶乙酸乙酯部位分离得到的没食子酸乙酯、山柰酚、木犀草素、槲皮素、紫云英苷等酚类成分均有很好的α-葡萄糖苷酶抑制活性,优于对照阿卡波糖,构效关系分析发现C环上C 3位的取代基种类和数目对化合物的抑制活性有影响。Striegel等[34]报道了红茶水提取物和红茶渣丙酮提取物均能够显著抑制酵母源α-葡萄糖苷酶和猪源胰腺淀粉酶的活性,成分分析结果显示二者含有高分子儿茶素聚合产物是其共同的物质基础。如前所述,食物的发酵等加工过程会引起酚类物质组成变化,发酵大豆经过发酵处理后,由于微生物的活动产生了大量的酶,引发食品中多酚含量及其单体组成发生显著变化,并可能引起生物活性的改变。日本科学家[35]通过14周的动物试验发现,一种日本传统发酵大豆产品可有效改善糖尿病小鼠糖代谢紊乱,推测发酵大豆多酚类成分的α-淀粉酶和α-葡萄糖苷酶抑制作用为其降血糖作用的核心机制。此外,在谷物和坚果多酚的类似研究[36]中也发现了对碳水化合物消化酶显著抑制作用,并且某些多酚的抑制效果可以和降血糖药物相匹敌。这种不同结构的酚类化合物对不同来源的α-葡萄糖苷酶相互作用所表现出来的差异普遍存在,这些差异性的研究结果主要是由于学者们未采用统一的评价方法所造成的,提示研究者在以α-葡萄糖苷酶抑制活性为靶点筛查活性化学成分时,尽可能选用化学、生物学特性接近人体小肠α-葡萄糖苷酶的类似物进行研究。
3.2 抑制葡萄糖转运
葡萄糖被肠道细胞吸收进入血液的过程须借助两种转运体介导,首先是由小肠上皮细胞刷状缘GLUT-2介导的易化扩散,其次是钠—葡萄糖共同转运体(SGLT-1)介导的主动转运。通过调控葡萄糖转运蛋白的表达和转位,可实现对小肠葡萄糖的转运吸收速率和数量的调节,进而实现对血糖稳态的调控目的。
一些广泛分布于果蔬类食物中黄酮被证实可以抑制GLUT-2和SGLT-1介导的葡萄糖和果糖的转运吸收,而且同一单体对不同的转运体抑制活性各不相同,表现出选择性抑制。Schulze等[37]结合人源SGLT1的cDNA在爪蟾卵母细胞中重组表达和小鼠外翻肠囊试验,明确苹果多酚提取物及其对SGLT1介导的葡萄糖转运的抑制作用,且不同的单体表现的抑制活性存在差异,根皮苷显示出最强的抑制作用。此外,当单糖随血液到达各组织器官后,膳食多酚还会对分布于各组织器官的特异性的单糖转运体的转运和表达产生影响,继续发挥对组织器官糖代谢稳态的调控作用。通过爪蟾卵母细胞中进行的类似试验[38]系统探究了类黄酮对葡萄糖转运体(GLUT2和 GLUT7)及果糖转运体(GLUT2、GLUT5和GLUT7)的抑制情况,结果发现GLUT2介导的葡萄糖和果糖转运被槲皮素、表没食子儿茶素没食子酸酯(EGCG)和芹黄素抑制,而GLUT5介导的果糖转运为EGCG和芹黄素所抑制,但不受槲皮素的影响。GLUT7介导的葡萄糖和果糖的摄入仅被芹黄素所抑制,不受槲皮素和EGCG影响。由此可见,膳食多酚与小肠上皮或各组织器官细胞上的葡萄糖、果糖转运体——SGLT1和GLUTs的相互作用极为复杂,主要体现在不同的酚类物质对不同类型的转运体的选择性抑制,而就与转运蛋白的结合机制而言,这些酚类物质与葡萄糖、果糖的相互关系又涉及竞争性或非竞争性,目前尚无一致性的研究结果。
3.3 改善胰岛素抵抗
当机体糖代谢主要靶组织如脂肪组织、骨骼肌、肝脏对胰岛素的敏感性降低后,胰岛素就不能发挥血糖代谢调节作用,即产生胰岛素抵抗并导致糖稳态失衡[39-41]。因此,通过药物、饮食和运动的干预可以减轻胰岛素抵抗,改善糖代谢稳态。胰岛素经血液循环到达相应靶器官后,首先与细胞膜上的胰岛素受体(IR)结合,激活酪氨酸蛋白激酶(protein Tyrosine kinase,PTK),继而激活胰岛素信号通路下游的系列信号因子,全面调控靶器官糖代谢的正常运行。任何外源或内源性化学物质,一旦引起胰岛素信号转导通路中任一细胞因子表达变化,均会影响胰岛素的生物效应。研究[42-44]表明,多种酚类物质可与该途径的多个分子靶点相互作用后改善胰岛素抵抗,促进葡萄糖的转运与代谢。罗莎藿香(一种维吾尔族药物)的多酚提取物对糖尿病大鼠的摄入试验[42-44]也显示出改善胰岛素抵抗指数和胰岛素敏感性的效果,推测该机制是通过胰岛素信号通路(PI3K/AKT)途径实现的。
多酚能调节胰岛素分泌,改善机体对胰岛素的敏感性。虽然肠道并不是葡萄糖代谢的主要场所,但作为葡萄糖和脂质吸收的重要器官,肠道对胰岛素的敏感性也是影响糖代谢稳态的重要因素。富含原花青素B2的葡萄籽多酚提取物可显著改善自发1型糖尿病db/db小鼠的体重、摄食量、胰岛素抵抗水平和胰岛体积等水平,促进胰岛素分泌,改善胰岛细胞损伤,显著增加胰腺中胰岛素的表达(P<0.05)[45]。膳食多酚对胰岛素分泌的改善作用,可能是由其中的一种或几种特殊单体贡献而来的。比较可可中提取的原花青素低聚物(包括表儿茶素、原花青素B2、原花青素C1和肉桂鞣质A2)对小鼠糖稳态调节激素的影响,发现仅肉桂鞣质A2能够增加GLP1(胰高血糖素样肽-1)和胰岛素的浓度,是提取物中活性最强的组分。
在人体试验中多酚通过改善胰岛素抵抗实现糖稳态调节作用也有大量的数据支持。一项研究[46]采用膳食多酚强化的食物对86名肥胖志愿者进行膳食干预治疗8周,发现多酚饮食可显著增加受试人早期胰岛素分泌(P=0.048)和胰岛素敏感性(P=0.05)。在浆果类的膳食干预试验[47]中也观察到类似的健康效应,该试验以41名超重和肥胖受试者为研究对象,双盲随机平行对照试验结果显示,受试者连续6周每日服用含有草莓和蔓越莓多酚的饮料(strawberry and cranberry polyphenols,SCP;多酚含量333 mg/杯),与对照组相比,胰岛素敏感性增加(P=0.03),并且在测试的前30 min后,SCP组胰岛素和C-肽分泌水平较低,证实富含草莓和蔓越莓多酚的食物干预可以有效改善超重受试者的胰岛素敏感性。
4 展望
高碳水化合物(如淀粉或蔗糖)的膳食会诱发高血糖症、高胰岛素血症。经常食用高糖饮食可能会增加肥胖,2型糖尿病等糖代谢紊乱相关的慢性疾病的患病风险。膳食多酚在控制血糖方面极具潜力,但综合目前的研究进展而言,尚有一些未知或有争论的问题有待进一步的深入研究:① 在动物和体外研究中,多酚对血糖的积极作用已经被大量研究所证实,但是对人类临床和分子机制的研究较少,确切的机制还尚无定论。② 已有的干预研究试验的结果并不一致,可能与不同的统计方式、样本量小(许多是急性研究)、缺少葡萄糖代谢的动态测量有关,应该进行更多具有明确饮食和对照研究设置的人体试验,找到揭示干预研究中细微饮食变化的早期生物标志物,并且对活性代谢物进行药代动力学分析。③ 诸多日常饮食,如茶、咖啡和红酒的多酚含量很高,但生物利用度却很低。使用不同的食物制备方法增加多酚的生物利用度也是一项具有挑战性的工作,生物利用度是促进多酚发挥生理功效的关键因素。④ 膳食多酚种类繁多,其数千种代谢物有许多种不同的定性和定量的组合,了解其吸收、代谢和组织分布、鉴定活性分子是一项非常具有挑战性的任务。⑤ 应该更加关注研究整株植物或者整个食物提取物的作用,以便跟踪同时存在于食物中的不同植物化学化合物的协同生物活性。酚类化合物与其他食物成分如纤维之间的相互作用也是一个值得探究的方向。
