环状RNA hsa_circ_0005320在三阴性乳腺癌中的表达及其对细胞增殖的影响
2019-12-13郑夏颖陈俊霞
郑夏颖,陈俊霞
重庆医科大学细胞生物学与遗传学教研室,重庆 400016
乳腺癌是全世界最常见的妇女恶性肿瘤,位居女性恶性肿瘤发病率第1位[1]。随着分子生物学及基因测序技术的不断发展,乳腺癌的分子分型不断完善,目前乳腺癌广泛使用的标准是2013年St.Gallen乳腺癌国际会议上规定的乳腺癌分类标准,分为Luminal A型、Luminal B型、人表皮生长因子受体2(human epidermal growth factor receptor 2,HER2)过表达型、基底样型/三阴性型及其他特殊类型乳腺癌[2]。三阴性乳腺癌(triple-negative breast cancer,TNBC)是指雌激素受体(estrogen receptor,ER)、孕激素受体(progesterone receptor,PR)和HER2均为阴性的一类乳腺癌,与其他亚型乳腺癌相比,恶性程度更高,更容易发生转移[3]。TNBC是一类异质性极高的恶性肿瘤,目前的治疗方式以化疗为主,尚缺乏有效的靶向治疗,因此亟待寻找诊断和治疗TNBC的新的生物标志物和靶分子。环状RNA(circular RNA,circRNA)是一类新发现的内源性非编码RNA,呈闭合环状结构,不含5'端帽子和3'端多聚腺苷酸尾巴[4]。研究表明,circRNA在包括乳腺癌在内的多种癌症的发生、发展过程中发挥着至关重要的作用[5-7],但目前有关circRNA在TNBC中的功能及其机制的报道尚少。本研究旨在探讨hsa_circ_0005320在TNBC中的表达及其对细胞增殖的影响。
1 材料和方法
1.1 材料
1.1.1 组织标本
本研究中的20例未接受放疗或化疗的TNBC组织及癌旁组织由重庆医科大学分子医学与肿瘤研究中心保存。该组织样本来自重庆医科大学附属第一医院,20例患者(均为女性)均签署知情同意书后进行手术切除,样本通过病理学检查证实并立即放于液氮中。临床资料和人TNBC标本的收集由重庆医科大学伦理委员会授权和监督。
1.1.2 细胞系及主要试剂
正常人乳腺上皮细胞系MCF-10A和人TNBC细胞系(MDA-MB-231和BT-549)购自美国ATCC细胞库。siRNA-阴性对照(siRNAnegative control,si-NC)和si-circ(si-hsa_circ_0005320)质粒及hsa_circ_0005320探针由广州吉赛生物科技股份有限公司设计合成;转染试剂LipofectamineTM2000购自美国Invitrogen公司;DMEM培养基和胰蛋白酶购自美国Gibco公司;MEBM培养基套装购自瑞士Lonza公司;胎牛血清(fetal bovine serum,FBS)购自重庆博全生物科技有限公司;TRIzol、PrimeScript RT Reagent Kit和TB Green购自日本Takara公司;聚合酶链反应(polymerase chain reaction,PCR)引物购自武汉金开瑞生物工程有限公司;Cell-LightTM EdU Apollo567In VitroKit购自广州锐博生物科技有限公司;细胞计数试剂盒(cell counting kit-8,CCK-8)购自武汉博士德生物工程有限公司;Hoechst 33342试剂购自北京鼎国昌盛生物技术有限责任公司;Tunel一步法测凋亡试剂盒购自上海碧云天生物技术有限公司;Anti-LIF抗体购自美国R&D公司,Anti-STAT3抗体购自英国Abcam公司,Anti-P-STAT3抗体购自美国CST公司。
1.2 方法
1.2.1 RNA测序
通过收集4对TNBC组织及癌旁组织,对其进行总RNA的提取并检测总RNA的质量和总RNA的完整度,质检合格后进行后续实验。在广州吉赛生物科技股份有限公司进行RNA测序(构建RNA测序文库、上机测序以及差异基因分析),根据测序结果筛选出TNBC组织及癌旁组织中差异表达的circRNA,并使用微阵列对包括hsa_circ_0005320在内具有明显差异表达的circRNA进行聚类分析。
1.2.2 细胞培养及转染
人乳腺上皮细胞系MCF-10A使用MEBM培养基,TNBC细胞(MDA-MB-231和BT-549)分别在含有10%FBS的DMEM培养基中培养,三株细胞均在37℃、CO2体积分数为5%的条件下培养,每间隔1~2 d更换新培养基。待细胞汇合度达到70%时,采用LipofectamineTM2000分别将si-NC和si-circ转染至MDA-MB-231细胞或BT-549细胞中,24、48 h后收获细胞。
1.2.3 实时荧光定量PCR(real-time fluorescent quantitative PCR,RTFQ-PCR)检测hsa_circ_0005320的表达
质粒转染24 h后,按照说明书用TRIzol法提取TNBC组织和癌旁组织、正常人乳腺上皮细胞(MCF-10A)和TNBC细胞(MDA-MB-231和BT-549)总RNA,琼脂糖凝胶电泳判断提取RNA的完整性,酶标仪检测其浓度和纯度。利用PrimeScript RT Reagent Kit将总RNA反转录为cDNA。利用美国Bio-Rad公司的RTFQ-PCR仪检测hsa_circ_0005320在TNBC组织和细胞中的相对表达水平。该研究所用特异性引物如下。GAPDH上游引物序列:5'-GAAGGTGAAGGTCGGAGTC-3',下游引物序列:5'-GAAGATGGTGATGGGATTTC-3';hsa_ci rc_00 05320上游引物序列:5'-TTG G AGAATTCAGAGCCTG-3',下游引物序列:5'-AAGATCTTTTCAAGGCCTCC-3'。以GAPDH为内参,cDNA为模板,在95 ℃预变性3 min;然后在95 ℃变性5 s,60 ℃退火30 s,72 ℃延伸30 s的条件下扩增40个循环。每个样品设置3个复孔,hsa_circ_0005320的相对表达量采用2-△△CT法表示。实验重复3次。
1.2.4 荧光原位杂交(fluorescencein situhybridization,FISH)实验检测hsa_circ_0005320的表达
将TNBC与癌旁组织经冰冻切片制备成8 μm厚度的组织切片。在0.2 mol/L HCl中室温处理15 min,0.25%胃蛋白酶37 ℃温育30 min,并用4%多聚甲醛固定20 min,然后加入杂交液55 ℃温育2 h;将探针与杂交液按1∶200的比例稀释,于PCR仪上85 ℃ 2 min变性,37 ℃2 min平衡后加入探针37℃避光温育过夜;加入DAPI复染20 min,抗荧光淬灭剂封片,置于荧光显微镜下拍照。hsa_circ_0005320探针序列为5'-AGATCTTTTCAAGGCCTCCTGGCT-3'。
1.2.5 EdU实验检测细胞增殖
接种至24孔板的各组细胞转染48 h后,4%多聚甲醛固定30 min,2 mg/mL甘氨酸脱色摇床温育5 min,0.5%TritonX-100脱色摇床温育10 min,然后根据Cell-LightTM EdU Apollo567In VitroKit试剂盒说明书对细胞进行Apollo染色及DNA染色,经抗荧光淬灭剂封片后置于荧光显微镜下拍照。
1.2.6 CCK-8实验检测细胞增殖
转染MDA-MB-231细胞或BT-549细胞24 h后,将各组细胞制备成100 μL的细胞悬液(密度为1×104/mL)接种到96孔板,置于温箱37 ℃、CO2体积分数为5%的条件下培养24、48、72、96 h后,每孔加入10 μL CCK-8溶液并混匀,然后将细胞放入温箱温育2 h,酶标仪测定450 nm处的吸光度(D)值,每组设置3个复孔。
1.2.7 流式细胞术检测细胞凋亡
MDA-MB-231细胞或BT-549细胞转染48 h后,胰酶消化细胞并用PBS洗3次,1 000 r/min离心5 min后弃上清,用Annexin结合液重悬细胞,每组再加入10 μL的Annexin V-PE室温避光温育15 min,使用流式细胞仪检测细胞凋亡。
1.2.8 Hoechst 33342染色实验检测细胞凋亡
接种至24孔板的各组细胞转染48 h后,用4%多聚甲醛固定1 h,经Hoechst 33342染色10 min,抗荧光淬灭剂封片,置于荧光显微镜下拍照。
1.2.9 Tunel实验检测细胞凋亡
24孔板中各组细胞转染至48 h后,4%多聚甲醛固定1 h,经0.1%TritonX-100冰上温育2 min,根据Tunel一步法测凋亡试剂盒配制Tunel检测液并于37 ℃下避光温育1 h,使用抗荧光淬灭剂封片,在荧光显微镜下观察细胞凋亡率并拍照。
1.2.10 流式细胞术检测细胞周期
转染MDA-MB-231细胞或BT-549细胞48 h后,PBS洗3次,1 000 r/min离心5 min后弃上清,70%乙醇轻柔重悬后4 ℃固定过夜。1 000 r/min离心5 min后去除乙醇,PBS洗3次,每组加入0.5 mL的PI溶液(50 μg/mL)室温避光温育30 min,使用流式细胞仪检测细胞周期。
1.2.11 蛋白质印迹法(Western blot)检测蛋白表达
收集6孔板中转染48 h后的MDA-MB-231或BT-549各组细胞,每组加入100 μL RIPA裂解液(PMSF∶RIPA=1∶100)后提取各组细胞总蛋白,每孔按30 μg蛋白进行上样。通过10%十二烷基硫酸钠聚丙烯酰胺凝胶电泳(sodium dodecyl sulfate polyacrylamide gel electrophoresis,SDSPAGE)恒压80 V电泳分离,再冰浴恒流200 mA转移到聚偏二氟乙烯(polyvinylidene fluoride,PVDF)膜上,在5%脱脂奶粉中常温摇床封闭2 h,然后4 ℃温育一抗(兔抗GAPDH 1∶5 000,STAT31∶1 000,P-STAT31∶1 000,羊抗LIF 1∶1 000)过夜,在37 ℃温育二抗(1∶5 000)2 h,经ECL化学发光显色,在Bio-Rad凝胶成像仪内拍照,并用Quantity One对图像进行分析。
1.3 统计学处理
采用SPSS 19.0和GraphPad Prism 6.0进行数据分析和作图,两样本均数比较采用t检验,多组均数比较采用方差分析,P<0.05为差异有统计学意义。
2 结果
2.1 hsa_circ_0005320在TNBC组织和细胞中高表达
通过RNA-Seq测序,在4对TNBC组织和癌旁组织中共检测到354个差异表达的circRNA(其中47个circRNA表达上调,307个circRNA表达下调)。应用聚类分析图分析部分差异表达的circRNA,其中hsa_circ_0005320在TNBC中上调较为显著(图1)。通过RTFQ-PCR检测20对TNBC组织和癌旁组织、正常人乳腺上皮细胞MCF-10A和TNBC细胞MDA-MB-231、BT-549中hsa_circ_0005320的表达,结果显示,hsa_circ_0005320在TNBC组织和细胞中均表达上调(图2A~B),该结果与测序结果一致,因此选择hsa_circ_0005320进行进一步的研究。通过FISH实验检测hsa_circ_0005320在TNBC组织和癌旁组织中的表达及定位,结果显示,hsa_circ_0005320在TNBC中高表达,且定位于细胞质(图2C)。

图1 RNA-seq分析显示TNBC组织和癌旁组织中部分差异表达的circRNA聚类图Fig.1 Heat map of differentially expressed circRNAs in TNBC tissues and para-cancerous tissues
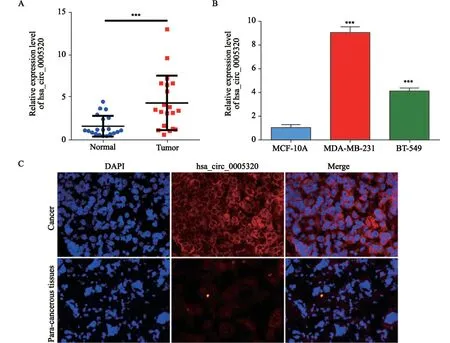
图2 hsa_circ_0005320在TNBC组织和细胞中的表达Fig.2 The relative expression of hsa_circ_0005320 in TNBC tissues and cells
2.2 hsa_circ_0005320干扰质粒效率验证
将si-NC和si-circ质粒分别转染MDA-MB-231和BT-549细胞24 h后,使用RTFQ-PCR检测细胞中hsa_circ_0005320的相对表达水平。结果表明,与si-NC组相比,si-circ组在MDA-MB-231细胞和BT-549细胞中hsa_circ_0005320的相对表达水平明显降低(图3)。
2.3 沉默hsa_circ_0005320抑制TNBC细胞的增殖
将si-NC和si-circ质粒分别转染MDA-MB-231和BT-549细胞后,通过EdU和CCK-8实验研究沉默hsa_circ_0005320对TNBC细胞增殖能力的影响。EdU实验结果发现,与si-NC组相比,si-circ组MDA-MB-231细胞和BT-549细胞的增殖能力明显降低(图4A~D)。CCK-8实验结果显示,连续观察48、72、96 h后,MDA-MB-231细胞和BT-549细胞的si-circ组细胞活性均显著低于si-NC组(图4E~F)。
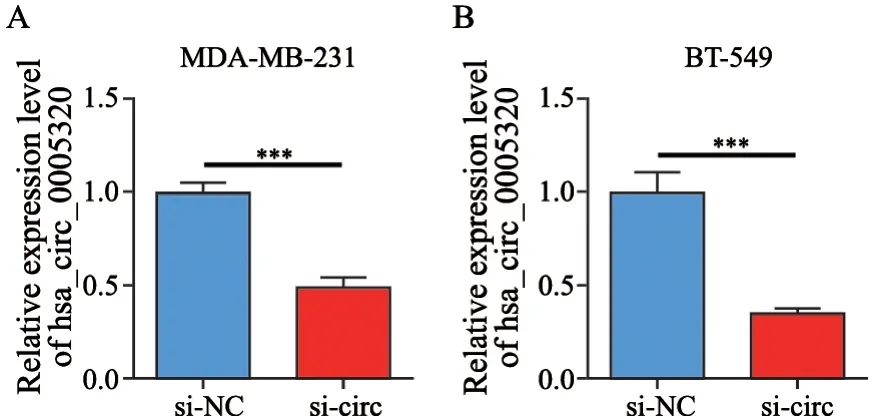
图3 hsa_circ_0005320干扰质粒效率验证Fig.3 Plasmid efficiency verification of hsa_circ_0005320 knockdown
2.4 沉默hsa_circ_0005320促进TNBC细胞的凋亡
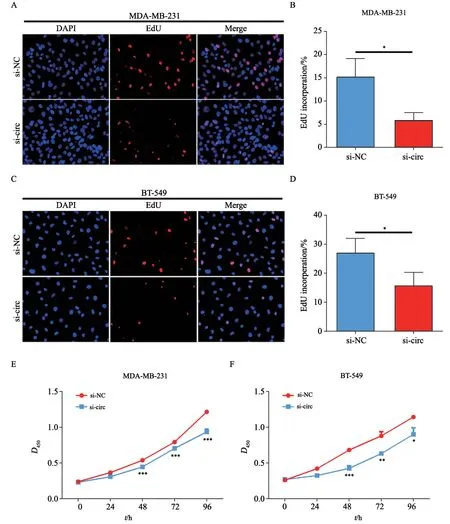
图4 沉默hsa_circ_0005320对TNBC细胞增殖能力的影响Fig.4 Effects of hsa_circ_0005320 knockdown on the proliferation of TNBC cells
MDA-MB-231细胞和BT-549细胞分别转染si-NC和si-circ质粒后,分别采用流式细胞术、Hoechst33342和Tunel实验检测沉默hsa_circ_0005320对TNBC细胞凋亡情况的影响。流式细胞术结果显示,与si-NC组相比,MDAMB-231细胞和BT-549细胞中si-circ组的凋亡率显著增加(图5A~B)。Hoechst33342实验表明,相比于si-NC组,si-circ组细胞的凋亡更明显,形态学特征表现为细胞核碎裂呈致密浓染,细胞质皱缩而发生高亮的荧光(图5C)。Tunel实验显示,si-circ组出现大量呈高亮荧光的凋亡细胞,而si-NC组的凋亡细胞较少(图5D)。
2.5 沉默hsa_circ_0005320对TNBC细胞周期的影响
将si-NC和si-circ质粒分别转染MDA-MB-231细胞和BT-549细胞后,应用流式细胞术分析沉默hsa_circ_0005320对TNBC细胞周期分布的影响。结果显示,两个处理组细胞绝大部分在G1期,S期细胞较少,G2期细胞最少。相较于si-NC组,si-circ组在G1期的细胞增多,在S期的细胞减少(图6A~C),表明沉默hsa_circ_0005320将细胞周期阻滞于G1期。
2.6 沉默hsa_circ_0005320调控LIF-STAT3通路相关蛋白的表达
收集转染si-NC和si-circ质粒48 h后的MDA-MB-231细胞和BT-549细胞提取总蛋白,Western blot检测LIF-STAT3通路相关蛋白的表达变化。结果显示,si-circ组相较于si-NC组,LIF和P-STAT3蛋白表达水平均明显下降(图7)。
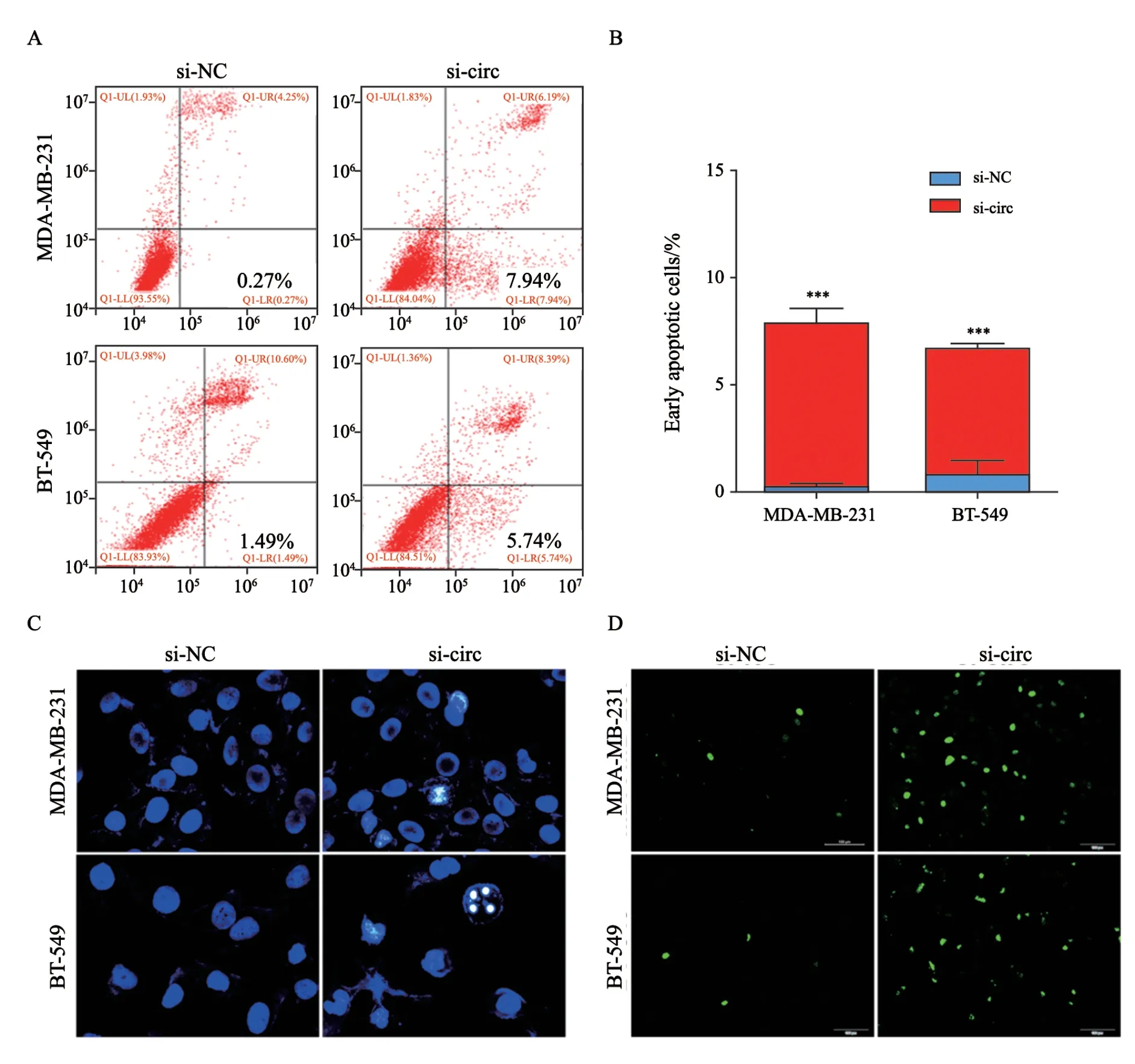
图5 沉默hsa_circ_0005320对TNBC细胞凋亡的影响Fig.5 Effects of hsa_circ_0005320 knockdown on apoptosis of TNBC cells
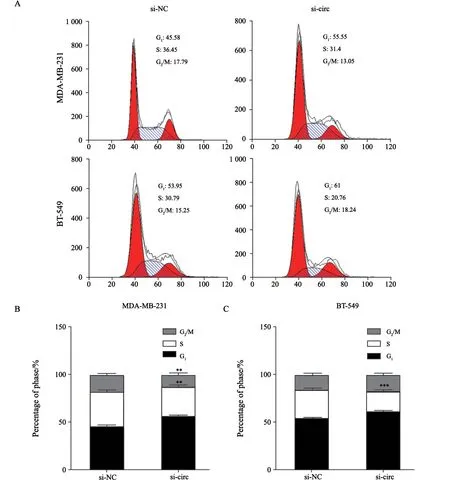
图6 沉默hsa_circ_0005320对TNBC细胞周期的影响Fig.6 Effects of hsa_circ_0005320 knockdown on cell cycle in TNBC
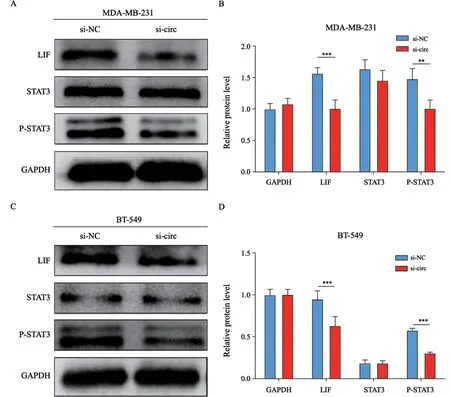
图7 沉默hsa_circ_0005320对TNBC细胞蛋白水平的影响Fig.7 Effects of hsa_circ_0005320 knockdown on protein levels of TNBC cells
3 讨 论
circRNA自首次在真核细胞的细胞质中被发现以来,一直被认为是错误剪接或内含子套索过程中形成的副产物。随着高通量测序技术和生物信息学分析的快速发展,大量circRNA在哺乳动物细胞中被发现,其环状结构由典型的剪接体介导或上游剪接体受体跟下游剪接体供体配对形成,在具有组织和发育阶段特异性特征的哺乳动物细胞中存在高度的稳定性和序列保守性,参与调控多种生物学进程[8-12]。circRNA可通过调控亲本基因的表达[13]、选择性剪接[14]、充当蛋白质复合物组装的支架[15]、作为miRNAs分子的海绵[6]以及RNA-蛋白相互作用[11]来调控基因表达。因此,circRNA的研究具有非常重要的意义。本研究中的hsa_circ_0005320来源于SEPT9基因的第2外显子,经剪接后形成长度为645 bp的环状转录本,已有文献报道其在癌症发生、发展过程中发挥着重要作用。Ju等[16]采用RNA微阵列技术发现了90个在转移性口腔黏膜黑色素瘤中差异表达的circRNA,通过RTFQ-PCR实验检测出hsa_circ_0005320在原发肿瘤和转移淋巴结中明显上调,结合生物信息学分析证实hsa_circ_0005320可以通过与特定的microRNAs结合作为竞争的内源性RNA,调控口腔黏膜黑色素瘤的发生和转移。
然而,hsa_circ_0005320在TNBC中的研究尚未见报道。本研究采用RNA-Seq技术分析4对TNBC组织及癌旁组织,获得差异表达的circRNA,利用RTFQ-PCR技术检测出hsa_circ_0005320在TNBC组织和细胞中表达水平均显著升高,进一步通过FISH实验证实hsa_circ_0005320在TNBC中呈高表达。为了进一步研究hsa_circ_0005320基因在TNBC中的作用,利用脂质体转染的方法将si-NC和si-circ质粒分别转染至MDA-MB-231和BT-549细胞中,RTFQ-PCR实验结果发现,si-circ组hsa_circ_0005320的表达水平显著降低。通过一系列功能实验发现沉默hsa_circ_0005320后,能够显著抑制TNBC细胞的增殖能力,促进细胞凋亡。最后通过Western blot实验证实hsa_circ_0005320可调节LIF-STAT3通路相关蛋白的表达,提示hsa_circ_0005320可能激活LIFSTAT3通路进而促进TNBC细胞的增殖。这些结果表明hsa_circ_0005320在TNBC的发生、发展过程中可能发挥重要作用,但其具体的分子作用机制仍有待进一步研究。
