花生酰基-CoA羧化酶基因ACC1的克隆与表达分析
2019-12-09刘浩鲁清李海芬洪彦彬陈小平梁炫强李少雄
刘浩 鲁清 李海芬 洪彦彬 陈小平 梁炫强 李少雄
摘要:ACC1基因编码酰基-CoA羧化酶,调节脂肪酸的从头合成。本研究克隆了花生ACC1基因,其DNA序列全长3 719 bp,编码区序列873 bp,蛋白质内含保守的酰基-CoA羧化结构域(124 ~ 288 aa)。利用Peanutbase数据库鉴定到15个ACC1同源基因;亚细胞定位发现ACC1主要定位于叶绿体。组织表达量分析表明该基因在种子与叶中的表达量最高,花与根瘤中的表达量较低。对不同发育期高、低油酸品种种子ACC1的表达量检测结果表明,高油酸能够上调ACC1的表达量。本研究为深入分析ACC1在脂肪酸合成通路中的分子功能提供了重要信息。
关键词:花生;脂肪酸;酰基-CoA羧化酶;克隆
中图分类号:S565.2:Q785文献标识号:A文章编号:1001-4942(2019)09-0021-07
Cloning and Expression Analysis of Acetyl-CoA
Carboxylase Gene ACC1 in Peanut
Liu Hao, Lu Qing, Li Haifen, Hong Yanbin, Chen Xiaoping, Liang Xuanqiang, Li Shaoxiong
(Crops Research Institute, Guangdong Academy of Agricultural Sciences/Guangdong Key Laboratory
of Crop Genetic Improvement, Guangzhou 510640, China)
Abstract ACC1 encodes the acetyl-CoA carboxylase, and regulates the de novo biosynthesis of fatty acid. In this study, ACC1 was cloned from peanut with DNA sequence as 3 719 bp and coding sequence as 873 bp, and its protein sequence contained conversed domain of cetyl-CoA carboxylase (124~288 aa). Fifteen homogeneous genes of ACC1 were identified by using the Peanutbase, and subcellular localization analysis indicated that ACC1 was mainly located in chloroplast. Tissue specific expression analysis suggested that the expression level of ACC1 was the highest in seeds and leaves, but lower in flowers and nodules. The expression level of ACC1 in the seeds of high- and normal-oleic acid peanut was examined, and the results indicated that high oleic acid up-regulated the expression level of ACC1 in peanut. This study could provide important information for deeply illustrating the molecular function of ACC1 in fatty acid synthesis pathway.
Keywords Peanut; Fatty acid; Acetyl-CoA carboxylase; Clone
花生(Arachis hypogaea)是我国重要的油料经济作物,种植面积超过5.5×106 hm 约占全球花生种植规模的1/5,占世界花生总产量的1/3,年产量超过1 600万吨[1, 2]。我国花生主要用于榨取食用油与鲜食加工,花生油因其风味独特、营养品质较高深受广大消费者喜爱。相比于大豆油与菜籽油,花生油不饱和脂肪酸(unsaturated fatty acids, UFA)含量較高,油酸(oleic acid, OA)与亚油酸(linoleic acid, LOA)约占总油脂的80%,且不含植物芥酸[3]。随着花生产业的快速发展,有关花生的应用研究取得了显著性突破:花生机械化进程加快、农机农艺融合推广范围逐渐扩大;优良品种育繁推效果显著;高油酸育种已达到国际领先水平。由于普通栽培种花生属于异源四倍体豆科作物[4],由二倍体的野生种A(A. duranensis)[5]与野生种B(A. ipaensis)[6]杂交形成,并经历了多倍化事件,因此,栽培种花生具有非常复杂的生物学特性,典型的表现为地上部开花而地下结果[7],种子发育需经历黑暗条件。相较于模式植物,花生的基础生物学研究进展较为缓慢,随着花生野生种与栽培种基因组的破译[8, 9],势必对花生功能基因组学研究注入新的驱动力。
植物油脂作为重要的食用油脂和工业原料,其需求量随着人口激增和工业发展不断增长,人类日常生活及饮食中有70%的油脂来自于植物油。鉴于植物油的重要性,深入了解油料作物脂肪酸代谢机理有助于提高油料作物的产量与改良植物油品质。长链脂肪酸的合成主要发生在植物细胞质体[10],16碳的棕榈酰(C16∶[KG-*2/3]0)与酰基载体蛋白(acyl carrier protein,ACP)结合生成最初的底物C16-ACP,经酮脂酰-ACP合成酶KAS Ⅱ(ketoacyl-ACP synthase)催化形成硬脂酰-ACP(C18∶[KG-*2/3]0-ACP)[11]。硬脂酸为18碳的饱和脂肪酸,经连续的脱饱和反应,18碳的长链脂肪酸不断引入双键(C=C)后生成不饱和的油酸、亚油酸和亚麻酸,其中定位于叶绿体的硬脂酰-ACP脱饱和酶(stearoyl-ACP desaturase,SAD)[12]与定位于内质网的脂肪酸脱饱和酶2(fatty acid desaturase 2,FAD2)、脂肪酸脱饱和酶3(FAD3)[13]是分别调控三种脂肪酸合成的关键限速酶。最后,不同类型的不饱和脂肪酸与甘油骨架形成花生油脂的主要成分三酰甘油(triacylglycerol,TAG),并存储于花生种子的子叶中。不同于长链脂肪酸的脱饱和过程(分别发生在质体与内质网),脂肪酸的从头合成主要发生在质体(黑暗条件)及叶绿体(光照条件)。光合产物丙酮酸经丙酮酸脱氢酶复合体(pyruvate dehydrogenase complex, PDCH)催化产生酰基辅酶A[14],随后乙酰辅酶A羧化酶(acetyl-CoA carboxylase, ACCase)[15]与单酰辅酶A酰基转移酶(malonyl-CoA: ACP malonyltransferase, MCMT)催化酰基辅酶A生成单酰基-ACP[16],以此为初始物经历7轮的碳链延伸生成棕榈酰-ACP(C16∶[KG-*2/3]0-ACP),每轮反应增加2个碳原子,其中酮酯酰-ACP合成酶 (KASⅠ)[17]、 酮酯酰-ACP还原酶(ketoacyl-ACP reductase, KAR)[18]、羟酰基-ACP脱氢酶(hydroxyacyl-ACP dehydrase, HAD)[19]、烯酰基-ACP还原酶(enoyl-ACP reductase, ER)[20]在该过程中起到非常重要的作用。
油脂合成过程中ACCase发挥着非常关键的作用,其催化酰基辅酶A发生羧化反应生成单酰辅酶A,是调节脂肪酸合成的初始步骤,同时,该基因亦参与其他的生理生化反应。拟南芥ACC1突变体叶片细胞不能正常分化,胚胎萌发以及根的生长均受到抑制,植株对低温反应敏感[15]。ACCase不仅调节脂肪酸的从头合成,而且影响长链脂肪酸的合成,并且调节植物叶片表面蜡质合成,参与多种激素的免疫抗性反应。油菜的高油酸突变体中,ACC1的转录表达会受到油酸过量积累的影响导致表达量降低[21]。前期研究发现在高油酸花生中,不论是转录水平还是蛋白质水平,FAD2突变导致的高油酸均会对ACC1的表达量产生影响,即生成产物反馈调节上游关键酶活性[22]。本研究对花生ACC1基因进行克隆,通过序列以及表达量分析初步明确了ACC1的基本功能,该研究为后续深入探究花生ACC1调控脂肪酸合成的分子功能奠定了基础。
1 材料与方法
1.1 试验材料
正常花生品种开农70与高油酸品种开农176种子,均由开封市农林科学院谷建中研究员提供, 并保存于国家油料改良中心南方花生分中心。2019年春季种植于广东省农业科学院白云基地,正常栽培, 于果针入土后的第20、30、40、50、60、70 d取发育的种子开展相关试验[23]。
1.2 总RNA提取及RT-PCR分析
分别取开花后的花生植株根、茎、叶、花、种子、根瘤、果针等部位,经液氮研磨后, 取0.1 g粉末, 利用Invitrogen的TRIZOL试剂盒提取总RNA[24]。取1 μg RNA, 利用FSK-100 cDNA反转录试剂盒(TOYOBO)合成第一条单链cDNA备用。采用ABI Stepone设备与天根SuperReal PreMix Plus(SYBR Green)试剂进行荧光定量PCR反应,总反应体积为20 μL。利用2–ΔΔCT计算差异基因表达量,每组荧光定量试验设3个重复。内参基因为adh3(F: 5′-GACGCTTGGCGAGATCAACA-3′,R: 5′-AACCGGACAACCACCACATG-3′),ACC1荧光定量检测引物为F:5′-ATGGACCTATACCTGCT-3′,R:5′-TAAGTTGTAGCTCAACG-3′。
1.3 亚细胞定位
将ACC1不含终止密码子的全长编码序列克隆到载体pNA580 (p35S-GFP)上, 将构建好的质粒转化大肠杆菌DH5α, 利用DNA提取试剂盒(Omega)提取大肠杆菌质粒, Nanodrop-1 000检测ACC1-GFP质粒浓度(约30 μg/μL)。将培养12 d的水稻黄化苗叶鞘切成0.5 mm细丝, 加入酶解液[含1.5% 纤维素酶(W/V)、0.75% 离析酶(W/V)、0.6 mol/L 甘露醇、10 mmol/L 吗啉乙磺酸、10 mmol/L 氯化钙、0.1%牛血清蛋白,pH 5.7]游离5 h, 用W5溶液(含154 mmol/L 氯化钠、125 mmol/L 氯化钙、5 mmol/L 氯化钾、2 mmol/L 吗啉乙磺酸,pH 5.7)洗涤原生质体2次, MMG溶液(含0.4 mol/L 甘露醇、15 mmol/L 氯化镁、4 mmol/L 吗啉乙磺酸,pH 5.7)重悬。加入10 μL ACC1-GFP质粒經40% PEG-4000介导共同转化水稻原生质体, 16 h后使用激光共聚焦显微镜Carl Zeiss LSM 780检测观察。ACC1-GFP载体构建引物为F:5′-GTTGTTGGATCCATGGCGTCTTGCACTATCCCTT-3′,R:5′-GTTGTTGGATCCTGGTACTATTACCAAAAGAGGCGT-3′。
1.4 生物信息学分析
ACC1序列信息参考花生基因组数据库(Peanutbase.org),基因与蛋白质结构采用绘制软件IBS,蛋白质进化分析采用MEGA软件,蛋白序列多重比对采用CLC sequence viewer 7,蛋白质保守结构域预测采用NCBI数据库中的BlastP,统计分析软件为GraphPad Prism 8。
2 结果与分析
2.1 ACC1基因与蛋白质结构
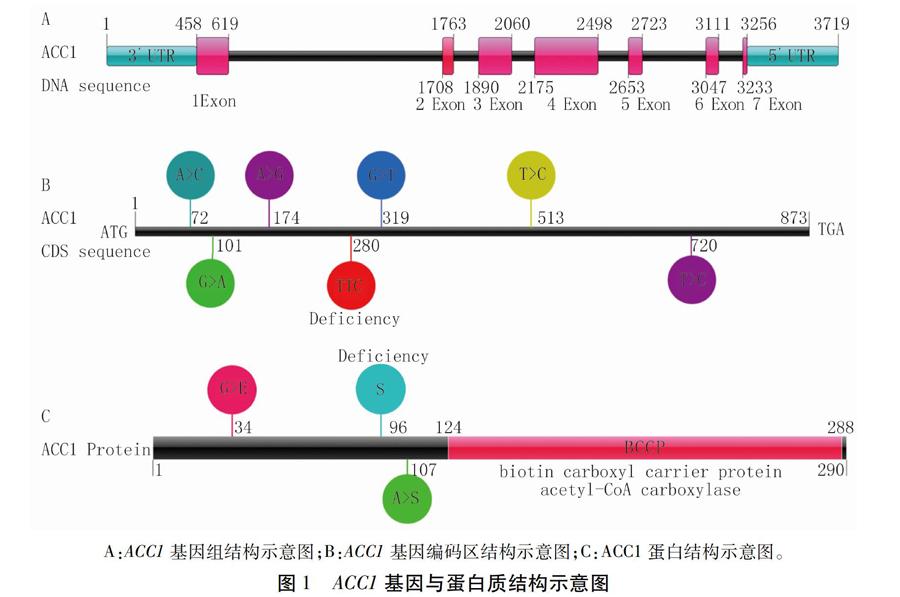
花生ACC1(arahy.KBA53Y)基因组序列全长3 719 bp,含有7个外显子与6个内含子(图1A)。编码区序列长873 bp,编码蛋白含有290个氨基酸(amino acid, aa)。相比于公布的花生基因组测序品种Tifrunner,开农70与开农176内ACC1编码区的280 bp处编码苯丙氨酸的密码子TTC缺失(图1B)。蛋白质序列分析表明由于编码区第72位的碱基由A突变为C以及第319位的碱基由G突变为T,导致氨基酸序列第34位的甘氨酸(G)突变为谷氨酸(E),第107位的丙氨酸(A)突变为L-丝氨酸(S),并且由于编码区280 bp处碱基缺失导致第96位的L-丝氨酸(S)缺失。蛋白质保守结构域预测结果表明ACC1蛋白中含有保守的BCCP结构域(124 ~ 288 aa)(图1C),该结构对ACC1发挥酰基羧化功能具有重要作用。
2.2 花生ACC1基因进化分析
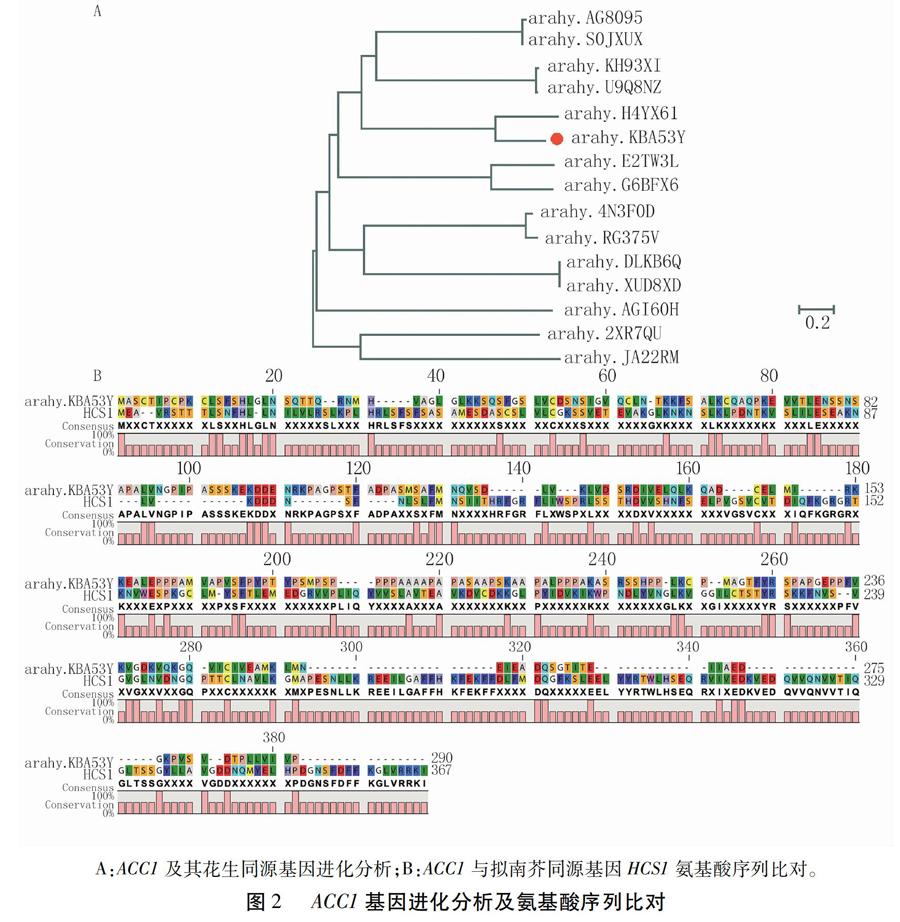
利用Peanutbase数据库,检索到15个ACC1同源基因,利用蛋白序列进行进化分析,发现ACC1(arahy.KBA53Y)与arahy.H4YX61属于不同亚基因组内的同一基因(图2A)。arahy.AGI60H仅有一个基因存在于亚基因组内,推测该基因可能在进化过程中发生了正向选择,其余ACC1同源基因均存在2种不同的亚基因组形式。ACC1在拟南芥中的同源基因为HCS1(AT2G25710),氨基酸序列比对发现,在1 ~ 82 aa与153 ~ 236 aa区间内两者的氨基酸序列同源性最高(图2B)。
2.3 ACC1亚细胞定位分析
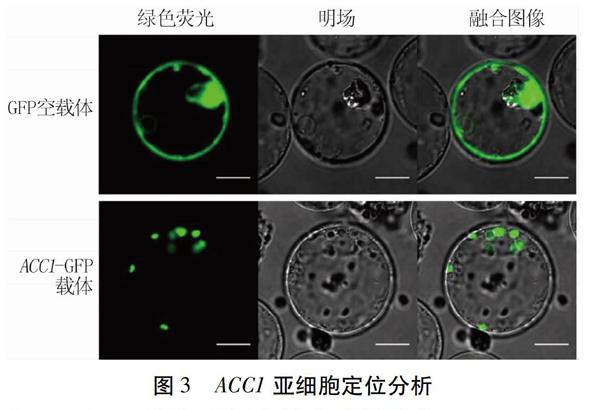
对ACC1-GFP的亚细胞定位进行活体检测显示, GFP空载体转化的水稻细胞液泡内未检测到荧光信号,细胞质内荧光强度较高;相比于对照组,ACC1- GFP融合蛋白释放的绿色荧光主要位于叶绿体部位, 表明ACC1主要定位在叶绿体(图3)。
2.4 ACC1在各组织中的表达量分析
以正常品种开农70为材料,以根部的样品为对照,利用荧光定量检测ACC1基因在不同组织中的表达量。结果表明ACC1在种子与叶中的表达量最高,其次为果针,茎与根部的表达量差异不大,根瘤中ACC1的表达量显著低于根中,而花中的表达量极显著低于根中(图4)。组织表达量分析表明ACC1虽能够在不同组织中检测到,但是存在明显的组织表达差异。
2.5 ACC1在高、低油酸品种内的表达量
高油酸品种开农176内ACC1(arahy.KBA53Y)及其等位基因arahy. H4YX61的表达量在种子发育早期低于正常品种开农70,种子发育中期表达量逐渐升高,发育后期表达量达到最大值(图5A、B)。荧光定量检测结果显示,在种子发育的第50 d,高油酸品种开农176 ACC1表达量最高(图5C),总体变化趋势与转录组测序结果相同。该结果表明高油酸(C18∶[KG-*2/3]1)积累会反馈促进上游基因ACC1的表达量。
3 讨论与结论
花生是我国重要的油料经济作物,相对于模式植物拟南芥,花生脂肪酸合成的分子机制并不清楚。花生属异源四倍体作物[25],基因组约2.7 GB,且含有两套亚基因组,其特殊的基因组结构导致同一基因可能存在多个拷贝数,而且大量的同源基因之间的冗余关系很难确定,为花生油脂基因功能研究带来了诸多难题。本研究利用花生基因组数据库克隆了花生酰基羧化酶基因ACC1,发现在花生基因组中ACC1具有15个同源基因(图2A),但是有关该类家族基因的功能尚不清楚。ACC1的拟南芥同源基因HCS(HOLOCARBOXYLASE SYNTHETASE )含有两个同源基因,HCS1[26]调解将生物素(biotin)添加到酰基-CoA羧化酶BCCP亚基上的反应,该基因信使RNA的5′UTR区间存在多种剪切模式,导致蛋白可定位在细胞质、线粒体以及质体中,调解蛋白翻译后的生物素修饰反应;但是HCS2[27, 28]过表达与干涉不会增加叶绿体内羧化酶的活性,可見HCS基因之间存在功能性的冗余。利用水稻原生质体进行亚细胞定位,发现ACC1蛋白定位于叶绿体(图3),组织表达量显示该基因在种子与叶中的表达量最高(图4),ACC1可能主要参与光合反应与脂肪酸合成。
鉴于油酸对人体健康较为有益,前期利用高油酸品种开农176种子发育的6个阶段进行转录组测序,发现开农176 ACC1的表达量高于正常品种开农70[29]。荧光定量验证也表明ACC1的实际表达量与转录组测序结果相符,该结果表明油酸过量积累对脂肪酸的从头合成通路产生影响(图5C)。ACC1表达量在高油酸种子内呈现上调表达趋势,但是导致该现象的分子机制仍有待深入研究。开农176产生高油酸的机理主要是因为FAD2[29]发生突变抑制了油酸向亚油酸转化,FAD2蛋白定位于内质网[30],油酸主要的合成场所也为内质网,通过形成油脂囊泡存储于细胞质内。但是,ACC1定位于质体,油酸过量积累是通过什么信号通路跨细胞器调节ACC1的表达仍不清楚;并且转录调节涉及大量的核内转录因子,ACC1的表达量升高肯定会受到核内转录因子的调控,即内质网-细胞核-质体间的未知信号可能是导致油酸调控ACC1表达的关键因子,然而揭示该机制仍需要深入研究。综上所述,本研究对ACC1的克隆与表达分析研究为后续深入理解ACC1在花生脂肪酸合成过程中的作用奠定了基础,解析ACC1功能的生物学意义对揭示高油酸反馈调节油脂合成基因亦具有重要意义。
参 考 文 献:
[1]李淞淋, 曹永跃. 世界花生和花生油生产、贸易发展动态及结构特征[J].世界农业, 2018(11): 113-119, 253.
[2]鲁清, 李少雄, 陈小平, 等. 我国南方产区花生育种现状、存在问题及育种建议[J].中国油料作物学报, 2017, 39(4): 556-566.
[3]Lu Q, Liu H, Hong Y, et al. Consensus map integration and QTL meta-analysis narrowed a locus for yield traits to 0.7 cM and refined a region for late leaf spot resistance traits to 0.38 cM on linkage group A05 in peanut (Arachis hypogaea L.)[J].BMC Genomics, 2018, 19(1): 887.
[4]Chen X, Lu Q, Liu H, et al. Sequencing of cultivated peanut, Arachis hypogaea, yields insights into genome evolution and oil improvement[J].Mol. Plant, 2019, 12(7): 920-934.
[5]Chen X, Li H, Pandey M K, et al. Draft genome of the peanut A-genome progenitor (Arachis duranensis) provides insights into geocarpy, oil biosynthesis, and allergens[J].Proc. Natl. Acad. Sci. U. S. A., 2016, 113(24): 6785-6790.
[6]Lu Q, Li H, Hong Y, et al. Genome sequencing and analysis of the peanut B-genome progenitor (Arachis ipaensis)[J].Front. Plant Sci., 2018, 9: 604.
[7]Chen X, Yang Q, Li H, et al. Transcriptome-wide sequencing provides insights into geocarpy in peanut (Arachis hypogaea L.)[J].Plant Biotechnol. J., 2016, 14(5): 1215-1224.
[8]Bertioli D J, Jenkins J, Clevenger J, et al. The genome sequence of segmental allotetraploid peanut Arachis hypogaea[J].Nat. Genet., 2019, 51(5): 877-884.
[9]Zhuang W, Chen H, Yang M, et al. The genome of cultivated peanut provides insight into legume karyotypes, polyploid evolution and crop domestication[J].Nat. Genet., 2019, 51(5): 865-876.
[10]Beld J, Lee D J, Burkart M D. Fatty acid biosynthesis revisited: structure elucidation and metabolic engineering[J].Mol. Biosyst., 2015, 11(1): 38-59.
[11]Wu G Z, Xue H W. Arabidopsis beta-ketoacyl-[acyl carrier protein] synthaseⅠis crucial for fatty acid synthesis and plays a role in chloroplast division and embryo development[J].Plant Cell, 2010, 22(11): 3726-3744.
[12]Kachroo A, Shanklin J, Whittle E, et al. The Arabidopsis stearoyl-acyl carrier protein-desaturase family and the contribution of leaf isoforms to oleic acid synthesis[J].Plant Mol. Biol., 2007, 63(2): 257-271.
[13]Dar A A, Choudhury A R, Kancharla P K, et al. The FAD2 gene in plants: occurrence, regulation, and role[J].Front. Plant Sci., 2017, 8: 1789.
[14]Ito J, Batth T S, Petzold C J, et al. Analysis of the Arabidopsis cytosolic proteome highlights subcellular partitioning of central plant metabolism[J].J. Proteome Res., 2011, 10(4):1571-1582.
[15]Lü S, Zhao H, Parsons E P, et al. The glossyhead1 allele of ACC1 reveals a principal role for multidomain acetyl-coenzyme A carboxylase in the biosynthesis of cuticular waxes by Arabidopsis[J].Plant Physiol., 2011, 157(3): 1079-1092.
[16]Jung S H, Kim R J, Kim K J, et al. Plastidial and mitochondrial malonyl CoA-ACP malonyltransferase is essential for cell division and its overexpression increases storage oil content[J].Plant Cell Physiol., 2019, 60(6): 1239-1249.
[17]Todd J, Post-Beittenmiller D, Jaworski J G. KCS[STBX]1 encodes a fatty acid elongase 3-ketoacyl-CoA synthase affecting wax biosynthesis in Arabidopsis thaliana[J].Plant J., 1999, 17(2):119-130.
[18]Venkatesan R, Sah-Teli S K, Awoniyi L O, et al. Insights into mitochondrial fatty acid synthesis from the structure of heterotetrameric 3-ketoacyl-ACP reductase/3R-hydroxyacyl-CoA dehydrogenase[J].Nat. Commun., 2014, 5: 4805.
