花生VAMP基因家族全基因组鉴定及表达分析
2019-12-09陆晓东张浩苑翠玲孙全喜闫彩霞赵小波王娟李春娟郑奕雄单世华
陆晓东 张浩 苑翠玲 孙全喜 闫彩霞 赵小波 王娟 李春娟 郑奕雄 单世华
摘要:植物囊泡结合膜蛋白(VAMP)是定位在囊泡上的运输蛋白,在植物发育以及响应生物和非生物胁迫中发挥重要作用。本研究对花生VAMP基因进行了全基因组鉴定与分析,并对它们在22个组织中的表达模式进行了分析。结果表明,二倍体野生种花生Arachis duranensis基因组有18个VAMP基因,Arachisi paensis基因组有21个,栽培种有41个,剔除假基因后花生VAMP基因家族有62个成员;表达模式分析表明AdVAMP17与AiVAMP1在大部分组织中均有表达,AdVAMP18与AiVAMP21在雌蕊中表达量最高,可能有特异性表达。本研究为进一步分析VAMP家族成员,并深入探讨其在花生中的生物学功能和进化模式奠定了基础。
关键词:花生;囊泡结合膜蛋白(VAMP)基因家族;囊泡运输;表达模式
中图分类号:S565.2:Q781文献标识号:A文章编号:1001-4942(2019)09-0042-08
Genome-Wide Identification and Expression Analysis of Peanut VAMP Gene Family
Lu Xiaodong1, Zhang Hao2, Yuan Cuiling2, Sun Quanxi2, Yan Caixia2, Zhao Xiaobo2, Wang Juan2, Li Chunjuan2,Zheng Yixiong1, Shan Shihua2
(1. College of Agriculture and Biology, Zhongkai University of Agriculture and Engineering, Guangzhou 510225, China;
2. Shandong Peanut Research Institute, Qingdao 266100, China)
Abstract Vesicle associated membrane protein (VAMP) is a transport protein located on vesicles of plants, which plays an important role in plant development and response to biotic and abiotic stresses. In this study, the genome-wide of peanut VAMP gene was identified and analyzed, and the expression patterns in 22 tissues were analyzed. The results showed that there were 62 members in peanut VAMP gene family, 18 of which were the Arachis duranensis genomes, 21 of which were the A. ipaensis genomes and 41 of which were the cultivars. Expression pattern analysis showed that AdVAMP17[STBZ] and AiVAMP1 were expressed in most tissues, and AdVAMP[STBX]18[STBZ] and AiVAMP[STBX]21[STBZ] had the highest expression level in pistils, which might have specific expression. These results laid a foundation for further analysis of the VAMP family members and discussing their biological functions and evolutionary patterns in peanut.
Keywords Peanut; Vesicle associated membrane protein (VAMP) gene family; Vesicle trafficking; Expression pattern
花生是我國重要的油料作物,在我国国民经济中占有重要地位,其生长过程遭受的各种生物与非生物胁迫对花生产量和品质造成了极大的影响[1]。培育抗逆品种是提高花生产量与品质的重要手段,抗逆基因的挖掘利用是培育抗逆品种的前提,花生全基因组测序工作的完成为挖掘抗逆基因奠定了基础[2]。
VAMP基因在植物发育以及响应生物和非生物胁迫中发挥重要作用[3]。Levine等[4]研究发现了拟南芥AtVAMP7蛋白超量表达能阻断BAX诱导酵母产生的程序性细胞凋亡(programmed cell death,PCD),从而改善细胞膜的组分与膜的完整性,阻止细胞凋亡,对膜起修复作用。薛媛[5]研究证明了拟南芥VAMP711参与了植物激素脱落酸(ABA)介导的质膜H+-ATPase活性调节机制,在干旱胁迫下,VAMP711基因通过与质膜H+-ATPase蛋白AHA1/AHA2互作,抑制质膜H+-ATPase的活性,促进气孔的关闭从而减少植物体内水分的流失,应对干旱胁迫。Sugano等[6]研究OsVAMP714基因敲除和过表达水稻植株并进行抗病性试验,证明了OsVAMP714参与了稻瘟病抗性且在水稻稻瘟病抗性中具有潜在的特异性,并对水稻植株生长发育有重要作用。李华一[7]的研究证明TaVAMP714基因参与了小麦条锈菌的感病反应,且ABA对TaVAMP714基因的诱导明显,干旱和盐胁迫都能诱导TaVAMP714上调表达。Kim等[8]的研究发现拟南芥VAMP721/722基因是将白粉病蛋白RPW82携带至植物-真菌界面这一激活抗性反应分泌机制的关键组分,能用于细胞外周的侵入前防御和EHM的侵入性防御,对致病性白粉病真菌起防御作用。方静[9]研究发现热处理诱导CitVAMP71基因表达量与翻译水平上升,椪柑果实通过增厚细胞壁响应热胁迫,进而提高了细胞的支撑作用,利于果实加强对外界病原菌侵染及其他生物胁迫的物理屏障。Sun等[10]研究发现GsVAMP72基因的过表达不仅能通过改变离子含量和下调应激反应基因的表达而降低大豆耐盐性,还能增加植物对ABA敏感性并改变ABA响应基因的表达水平。
目前,关于VAMP基因家族在花生中的研究尚未见报道。本研究利用生物信息学方法从全基因组水平对花生VAMP基因家族进行系统挖掘,并对该基因家族的基因结构、染色体定位、组织表达模式等进行系统分析,以期为后续研究此类基因在花生中的生物学功能奠定基础。
1 材料与方法
1.1 序列收集
在TAIR(TAIR release 10, http://www.arabidopsis.org/)数据库下载拟南芥VAMP家族蛋白全长序列[11],Pfam数据库(http://pfam.Xfam.org/)下载VAMP家族蛋白结构域序列,将两类序列在花生基因数据库Peanutbase(https://www.peanutbase.org/)和NCBI数据库(http://www.ncbi.nlm.nih.gov/)中进行Blastp检索(E值≤10-15),获得花生VAMP家族类似序列,人工去除冗余序列,作为VAMP家族的候选序列并编好基因名称。结合Pfam数据库中VAMP结构域保守序列和SMART在线分析软件(http://smart.embl-eidelberg.de/)对候选序列进行VAMP结构域分析,剔除不含有该结构域的蛋白质序列,得到花生VAMP基因家族全体成员序列。
1.2 系统发育进化树构建与染色体定位分析
根据获得的拟南芥与花生VAMP家族蛋白全长序列数据信息,将其氨基酸序列在MEGA 7.0上进行序列比对,采用非加权组平均法(unweighted pair-group method with arithmetic means, UPGMA)对比后获得的序列构建系统发育进化树,采用Bootstraping法对进化树进行评估与分组。校验参数: p-distance重复1 000次。
利用ExPASy-ProtParam(http://web.expasy.org/protparam/)对所得花生VAMP基因家族蛋白序列的分子量及等电点等理化性质进行分析。
从花生基因组中的gff3文件中提取VAMP基因家族的染色体起始和结束位置,利用MapInspect软件进行染色体物理分布图的绘制,获得VAMP成员的染色体物理定位信息。
1.3 花生VAMP家族基因结构及保守基序分析
由Gene Structure Display Server 2.0 (http://gsds.cbi.pku.edu.cn/)在线网站提供基因结构预测的可视化工具[12]。将去除假基因的花生VAMP基因家族的CDS序列及基因组序列在GSDS程序中得到其基因结构示意图,在基因结构图上标注特定区域,得到VAMP基因家族基因结构图。
利用MEME 4.12(http://meme-suite.org/tools/meme)[13]在线网站对VAMP家族基因的保守基序进行分析。
1.4 花生VAMP基因表达量分析
利用Heml软件分析花生VAMP基因在花生22个组织中的表达情况。
2 结果与分析
2.1 花生VAMP基因家族鉴定与理化性质分析
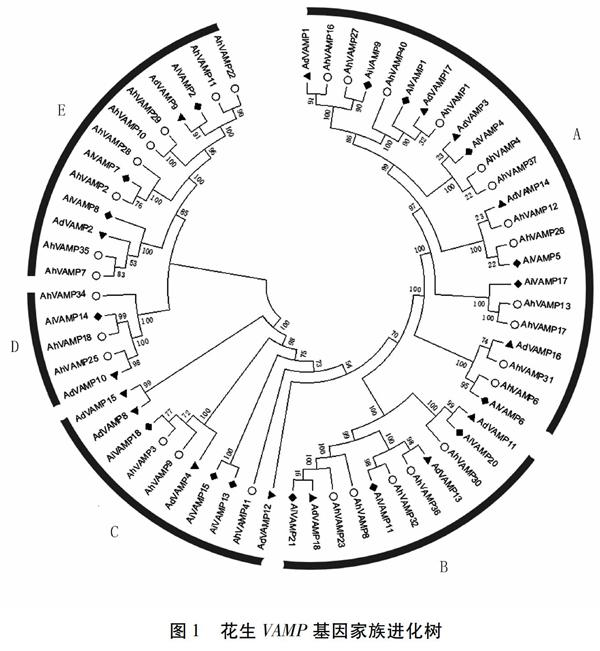
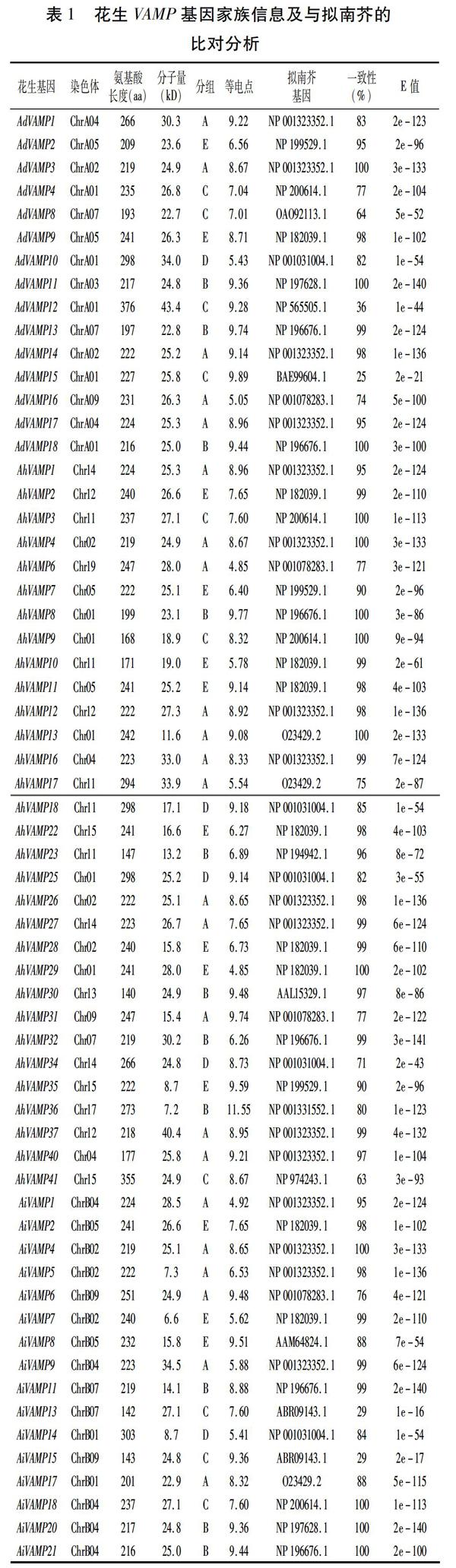
从两个二倍体野生种花生A.duranensis和A.ipaensis基因组中分别得到18个和21个VAMP基因,在普通花生栽培种中得到41个VAMP基因家族成员,分别命名为AdVAMP1~18、AiVAMP1~21、AhVAMP1~41(表1)。利用Pfam、SMART等软件检验这80个基因家族成员,发现其中有18个成员具有不完整的结构域,可能为假基因,剔除这18个成员,将拥有完整结构域的62个花生VAMP基因家族成员构建系统进化树,根据其拓扑结构,将花生VAMP基因家族分为5组,分别命名为group A、B、C、D、E。
在進化树中我们发现group A拥有最多的花生VAMP基因家族成员,为23个;其次是group E(13个)、group B(11个)、group C(10个)、group D(5个)(图1)。由进化树分析可知,花生VAMP蛋白家族形成两大分支,共有24对旁系同源基因,其中有5对旁系同源对bootstrap值小于70,其他19对旁系同源对bootstrap值在70以上,其中有15对旁系同源对bootstrap值大于90,因此我们认为花生VAMP蛋白在进化上亲缘关系较为接近。
理化性质分析发现,VAMP基因家族成员的氨基酸长度差异较小,其长度在140(AhVAMP30)~376 aa(AdVAMP12)之间,且大部分基因家族成员氨基酸长度在200~300 aa之间;花生VAMP蛋白分子量在6.6 kD(AiVAMP7)与43.4 kD(AdVAMP12)之间;等电点分布在4.85(AhVAMP6、AhVAMP29)与11.55(AhVAMP36)之间,有17个VAMP蛋白偏酸性,等电点小于7.0,其他45个VAMP蛋白呈中性或偏碱性。为了探究花生VAMP家族基因的生物学功能,将其与模式植物拟南芥进行了比对分析(表1)。结果表明,花生VAMP基因家族与拟南芥对应的基因之间有较好的同源关系(E-value<3.00e-141)。该结果为研究花生VAMP基因家族的功能提供了一定的借鉴。
2.2 花生VAMP基因家族的染色体分布
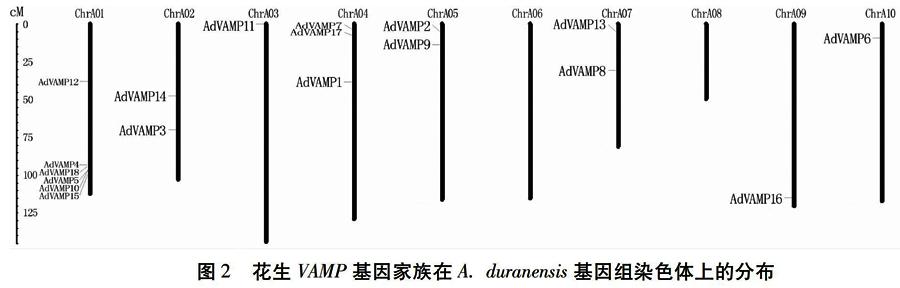
染色体定位分析有助于更直观地了解花生VAMP基因家族在染色体中的分布情况,由图2~图4[JP]可知VAMP基因家族成员在染色体上并没有均匀分布。在A.duranensis基因组染色体A06与A08上并无分布,而在染色体A01上分布最多,高达6个VAMP基因(图2);在A.ipaensis基因组染色体B10上无分布,而在染色体B01及B02上均分布有4个成员(图3);在栽培种染色体06、染色体08、染色体10、染色体16、染色体20上无分布,而在染色体01与染色体11上均分布有6个成员(图4)。由此可见,花生VAMP基因家族成员在染色体上的位置分布并不均匀,且对于花生VAMP基因家族40条染色体的观察发现多数基因家族成员分布在染色体的两端,而靠近着丝粒部分的基因较少。
2.3 VAMP家族基因结构及保守性分析
使用MEME软件对花生62个VAMP基因家族成员所编码的蛋白进行保守基序分析,获得6个motif的信息,分别命名为motif 1~motif 6。预测结果如图5所示,有23个基因成员均含有motif 1、motif 2、motif 4、motif 5,有16个基因家族成员仅含有motif 3和motif 6两个结构域,motif 1、motif 2、motif 4在VAMP基因家族中分别存在36、38、35个,由此,我们认为VAMP基因家族有较为保守的结构。此外,我们观察到group A与group B大部分基因编码的蛋白均含有motif 1、motif 2与motif 4,而group C基因成员编码的蛋白则含有motif 1、motif 2、motif 4与motif 5。这说明group A与group B功能可能较为类似,group C与group A及group B亲缘关系较近,基因可能有较为类似的功能。而group D与group E大部分基因编码的蛋白仅含有motif 3与motif 6,说明这两组基因家族成员与group A、group B、group C亲缘关系较远,基因编码蛋白功能可能差异较大。我们还分析了62个花生VAMP基因家族的结构,发现5组基因家族成员之间结构差异较大,而组内的结构相对接近(图5)。大部分基因家族成员都由外显子(exon)、非编码区(UTR)和内含子(intron)组成,仅有AiVAMP11、AiVAMP20、AhVAMP3、AhVAMP8、AhVAMP9、AhVAMP23这6个基因家族成员仅由内含子和外显子组成。我们注意到,在VAMP基因家族中,5组基因家族的内含子外显子结构分化严重,变化较大,其中group C的平均长度在五组里面最短,group A平均长度最长。
2.4 VAMP基因家族表达模式分析
基因表达模式分析有利于我们研究VAMP基因家族的功能。利用苗期叶片等22个组织的转录组数据,对花生A.duranensis与A.ipaensis基因组共31个成员在花生22个组织中的表达模式进行分析,由图6可知,大部分VAMP基因家族成员在22个组织中有不同程度的表达,且在不同组织或器官中,不同基因家族成员的表达模式存在明显差异。在本聚类图中,用蓝、绿、黄、红深度表示基因表达的强弱情况,颜色越接近红色表达情况越强,黑色则表示该成员在此组织中未见表达。AdVAMP17与AiVAMP1表达量最高,整体呈红色,说明这两个蛋白在花生22个组织中均有表达,且表达量是所有VAMP基因家族最高的;而AdVAMP18、AiVAMP21、AdVAMP14、AiVAMP5、AiVAMP13、AiVAMP15与AdVAMP8共7个基因家族成员在多数组织中未见表达或表达量较低,但在雌蕊(pistils)中,AdVAMP18与AiVAMP21两个基因成员的表达量是所有VAMP基因家族成员中最高的,表明在雌蕊中AdVAMP18与AiVAMP21可能有特异性表达。
3 讨论与结论
花生是我国重要的油料作物,生长过程中遇到的胁迫问题复杂而多样,挖掘和利用抗逆基因具有重要意义。众多学者的研究表明VAMP蛋白介导的囊泡运输在植物对抗逆境胁迫中发挥重要作用,因此对于VAMP基因家族的研究不容忽视。近年来,关于植物VAMP基因家族克隆及功能方面的研究集中在拟南芥、水稻、麦类等植物,主要参与抗病、干旱、盐胁迫等逆境反应[4-10,14,15]。因此,我们预测花生VAMP基因家族可能参与抗逆反应。
本研究基于花生全基因组和22个组织转录组测序数据,首次对花生VAMP基因家族进行了系统发育树、保守域、保守基序、染色体定位、22个组织表达模式的分析,共鉴定到62个花生VAMP蛋白,并根据进化树与基因结构将其分为5组。花生VAMP基因家族的保守基序非常明显地将其分为了两组,我们推测拥有motif 1、motif 2、motif 4的A、B、C组基因功能可能较为接近,拥有motif 3和motif 6的D、E组基因与前者可能有不一样的功能。近年来关于植物功能基因家族的研究表明,在水稻、拟南芥、陆地棉等植物中,拥有相同保守基序或功能域的氨基酸序列在系统发育树相同的组内是高度保守的,这些同一组内含有相同基序的蛋白或具有相似功能[16]。
基因的表达模式分析可以为未知基因功能的预测做一定的贡献。我们分析了花生VAMP基因家族在22个组织中的表达模式,发现AdVAMP18和AiVAMP21在雌蕊中表达量最高,雄蕊次之,而这两个基因在其他组织中几乎不表达或者表达量很低,而AdVAMP14和AiVAMP5也在雌蕊中有较高表达,因此我们推测AdVAMP18、AiVAMP21、AdVAMP14和AiVAMP5在花及种子形成过程中可能发挥着重要作用。而AdVAMP8基因在花生种子中不表达,暗示其启动子可能是个非种子表达启动子,可以用于花生遗传转化,使转化基因在花生种子中不表达,避免人们对转基因的恐慌。同时,我们发现在花生VAMP基因家族染色体定位中,A.duranensis基因组的AdVAMP16基因与A.ipaensis基因组的AiVAMP6均在09号染色体,四倍体栽培种花生的AhVAMP31、AhVAMP6在09号及19号染色体上具有相似的位置,结合表达聚类图中AdVAMP16基因与AiVAMP6基因在花生22个组织中相似的表达模式及在基因结构分析中相似的内含子、外显子结构,我们认为这四个基因可能为同源基因,具有相同的功能。同理,A.duranensis基因组的AdVAMP18基因与A.ipaensis基因组的AiVAMP18基因均在1号染色体,四倍体栽培种花生的AhVAMP8、AhVAMP18等在1号染色体与11号染色体上也具有类似位置,也可能為具有相同功能的同源基因。具体基因功能的探索还需要进一步试验验证。
本研究首次系统分析了VAMP蛋白在花生中的情况,包括染色体定位、基因结构、保守结构域、表达模式等,为下一步深入研究此类蛋白提供了参考。
参 考 文 献:
[1]Sharma S, Pandey M K, Sudini H K, et al. Harnessing genetic diversity of wild Arachis species for genetic enhancement of cultivated peanut[J].Crop Science, 2017, 57(3): 1121-1131.
[2]Zhuang W, Chen H, Yang M, et al. The genome of cultivated peanut provides insight into legume karyotypes, polyploid evolution and crop domestication[J].Nature Genetics, 2019, 51(5): 865.
[3]Fujii H, Zhu J K. Osmotic stress signaling via protein kinases[J].Cellular and Molecular Life Sciences, 2012, 69(19):3165-3173.
[4]Levine A, Belenghi B, Damari-Weisler H, et al.Vesicle-associated membrane protein of Arabidopsis suppresses Bax-induced apoptosis in yeast downstream of oxidative burst[J].Journal of Biological Chemistry, 2001, 276(49):46284-46289.
[5]薛媛. 拟南芥SNARE蛋白VAMP711通过调控质膜H+-ATPase响应干旱胁迫的机制研究[D].北京:中国农业大学,2019.
[6]Sugano S, Hayashi N, Kawagoe Y, et al. Rice OsVAMP714, a membrane-trafficking protein localized to the chloroplast and vacuolar membrane, is involved in resistance to rice blast disease[J].Plant Molecular Biology, 2016, 91(1/2):81-95.
[7]李华一. 小麦SYP71和VAMP714基因在小麦与条锈菌互作中的功能分析[D].杨凌:西北农林科技大学,2014.
[8]Kim H, OConnell R, Maekawa-Yoshikawa M, et al. The powdery mildew resistance protein RPW8.2 is carried on VAMP721/722 vesicles to the extrahaustorial membrane of haustorial complexes[J].The Plant Journal, 2014, 79(5):835-847.
[9]方静. 热处理诱导椪柑果实抗逆相关蛋白VAMP表达的初步研究[D].武汉:华中农业大学,2011.
[10]Sun X, Ji W, Ding X, et al. GsVAMP72, a novel Glycine soja R-SNARE protein, is involved in regulating plant salt tolerance and ABA sensitivity[J].Plant Cell Tissue and Organ Culture, 2013, 113(2):199-215.
[11]Poole R L. The TAIR database[M].Plant Bioinformatics, 2005.
[12]郭安源, 朱其慧, 陈新, 等. GSDS:基因结构显示系统[J].遗传, 2007, 29(8):1023-1026.
[13] Bailey T L, Johnson J, Grant C E, et al. The MEME suite[J].Nucleic Acids Research, 2015, 43(W1):W39.
[14]豆献英. 稻瘟病菌Mgvam7基因的克隆与功能分析[D].南京:南京农业大学,2008.
[15]裴洪翠. 麦类作物抗锈病相关蛋白互作网络的研究[D].泰安:山东农业大学,2014.
[16] Tang Y, Qin S, Guo Y, et al. Genome-wide analysis of the AP2/ERF gene family in physic nut and overexpression of the JcERF011 gene in rice increased its sensitivity to salinity stress[J].PLoS ONE, 2016, 11(3): e0150879.
收稿日期:2019-07-24
基金项目:泰山学者特聘专家(ts201712080);山东省农业良种工程项目(2017LZN033,2017LZGC003);山东省现代农业产业技术体系(SDAIT-04-02);国家自然科学基金青年科学基金项目(31601336);广东省农业农村厅科技项目(2019KJ136);山东省重点研发计划项目(2016GGH4518);青岛市科技惠民计划项目(17-3-3-49-nsh)
作者简介:陆晓东(1994—),女,在读硕士研究生,研究方向为花生遗传育种。E-mail:xdongdonglu@163.com
通讯作者:郑奕雄(1963—),男,教授,主要从事花生育种与栽培研究。E-mail: gdsscqs@163.com
单世华(1971—),男,研究员,主要從事花生种质创新及利用研究。E-mail:shansh1971@163.com
