凤丹牡丹PoFAD7基因的克隆及表达分析
2019-11-20李超琼王雪芹刘红占王会芳朱英男田辉辉
李超琼,王雪芹,刘红占,王会芳,朱英男,田辉辉
(1.周口师范学院 生命科学与农学学院,河南 周口 466001; 2.周口师范学院 新闻与传媒学院,河南 周口 466001)
牡丹隶属于芍药科芍药属牡丹组,为多年生小灌木,具有很高的观赏和药用价值。自2011年牡丹籽油被批准为新资源食品,油用牡丹作为一种新型木本食用油料植物引起广泛关注。目前,国家公布的油用牡丹品种为凤丹牡丹和紫斑牡丹,其中凤丹牡丹以生长适应性强、种子产量高等显著特点在全国大面积种植[1-3]。牡丹籽油含有丰富的不饱和脂肪酸,主要为α-亚麻酸、油酸、亚油酸等,总不饱和脂肪酸含量可达93.23%[4-5]。α-亚麻酸属于ω-3 系列不饱和脂肪酸,为人体不能自主合成的必需脂肪酸,是二十二碳六稀酸(DHA)和二十碳五稀酸(EPA)的前体,具有降血脂、降胆固醇、促进大脑发育和预防心脑血管疾病等作用[1, 6-8]。
ω-3脂肪酸脱氢酶是高等植物脂肪酸生物合成途径中的关键酶,将亚油酸催化合成亚麻酸,是α-亚麻酸合成的关键酶[9]。在高等植物中,ω-3脂肪酸脱氢酶可分为定位于内质网中的FAD3和定位于质体的FAD7和FAD8[10-11]。目前,FAD7基因在拟南芥、大豆、油菜、烟草、麻疯树、光皮树等多种植物中得到克隆,且发现FAD7基因能响应多种环境因素,如低温、创伤等[9, 12-14]。研究发现,超量表达拟南芥FAD7基因的转基因烟草α-亚麻酸含量增加,亚油酸含量减少,且转基因植株抗寒能力也得到提高,在1℃低温下仍能正常生长[15]。另外,超量表达FAD7基因的转基因番茄抗寒性也有所增加[16]。由此可见,FAD7基因不仅影响植物的脂肪酸组成,还能改良植物对低温的适应能力。
本研究从凤丹牡丹中克隆得到α-亚麻酸生物合成的关键基因PoFAD7,并进行生物信息学分析及其在不同种子发育时期的表达模式研究,为进一步研究其在油用牡丹不饱和脂肪酸合成过程中的调控机理奠定基础。
1 材料与方法
1.1 试验材料
凤丹牡丹种植于周口师范学院试验田,2016年4—8月采集凤丹牡丹不同发育时期的种子,样本经液氮速冻后于-80℃冰箱保存待用。
多糖多酚总RNA提取试剂盒购自北京天根,Trans 2K plus II DNA Marker、TransStar taq酶和感受态细胞购自北京全式金生物技术有限公司,RACE试剂盒、T/A克隆试剂盒、反转录试剂盒(PrimeScript® RT Reagent Kit with gDNA Eraser)购自大连宝生物工程有限公司。
1.2 试验方法
1.2.1PoFAD7基因的克隆
以凤丹牡丹种子不同发育期转录组测序数据(SRA accession number: SRP051810)为基础,找到一条含有部分PoFAD7基因的EST序列,根据该序列设计特异引物(见表1)进行3′-RACE。按照多糖多酚总RNA提取试剂盒操作步骤提取总RNA,检测总RNA完整性和浓度,并按照RACE试剂盒操作步骤反转录合成cDNA。利用已设计好的引物进行RT-PCR扩增获得PoFAD7基因3′端基因序列片段,与转录组数据中的EST序列进行拼接获得PoFAD7基因完整的开放阅读框。以PoFAD7基因序列为模板,设计含有起始密码子和终止密码子的特异引物,扩增获得PoFAD7基因的开放阅读框全长序列。

表1 引物序列
1.2.2PoFAD7基因cDNA序列生物信息学分析
使用在线工具ProtParam(http://web.expasy.org/protparam/)进行蛋白质亲水性预测和理化性质分析。使用在线工具TMHMM (http://www.cbs.dtu.dk/services/TMHMM/)分析PoFAD7 蛋白质序列跨膜结构域。从GenBank下载其他物种FAD7蛋白氨基酸序列,使用DNAMAN软件进行序列相似性比对。应用软件MEGA5.0邻位连接法构建PoFAD7 蛋白的系统进化树。
1.2.3 凤丹牡丹种子亚油酸和α-亚麻酸含量测定
自凤丹牡丹授粉后每隔10 d采集不同发育时期的种子,在45℃烘箱中烘至恒重。采用索氏提取法提取凤丹牡丹籽油,所得牡丹籽油经甲酯化处理,以气相色谱-质谱联用仪(Agilent 5977A Series GC/MSD System)分离和鉴定脂肪酸组分,测定干燥种子中亚油酸和α-亚麻酸的含量。具体步骤参照秦亚龙等[17]的方法。
1.2.4PoFAD7基因定量表达分析
提取凤丹牡丹不同发育时期的种子总RNA并反转录为cDNA为模板,并以获得的PoFAD7基因序列为模板设计特异性引物,以UBQ基因为内参基因进行荧光定量PCR试验检测PoFAD7基因的表达情况。荧光定量PCR按照SYBR Premix Ex TaqTMⅡ(TaKaRa)试剂盒说明在仪器 CFX96 Real-Time PCR Detection System (Bio-Rad 公司)上进行。扩增反应程序为:95℃ 3 min,95℃ 15 s,58℃ 15 s,72℃ 15 s,扩增40个循环。试验所用引物见表1。
2 结果与讨论
2.1 PoFAD7基因克隆
根据凤丹牡丹种子转录组数据获得951 bp的PoFAD7基因片段,依据所获得的片段设计特异性引物,以凤丹牡丹种子cDNA为模板进行3′-RACE,获得3′末端序列长度约为800 bp(见图1)。对所获得的基因片段进行拼接,获得全长cDNA序列为1 666 bp。根据所拼接的序列设计包含起始密码子和终止密码子的特异性引物进行PCR,获得包含一个完整的开放阅读框(1 353 bp)的基因片段(见图1),编码450个氨基酸(见图2)。将该片段与拼接的PoFAD7基因序列进行比对分析,显示两序列一致,表明PoFAD7基因拼接正确。
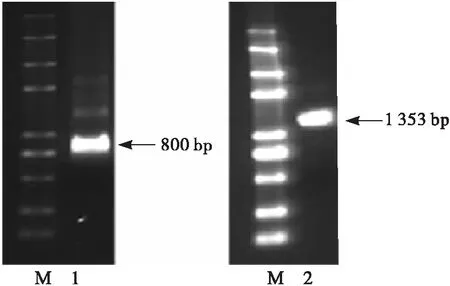
注:M. Trans 2K plus Ⅱ;1. 3′-RACE; 2.凤丹牡丹PoFAD7基因开放阅读框PCR产物。
图1 凤丹牡丹PoFAD7基因克隆

注:方框表示起始密码子ATG;“*”表示终止密码子。
2.2 PoFAD7蛋白理化性质及氨基酸序列比对
ProtParam分析结果显示,PoFAD7蛋白分子式为C2 359H3 543N643O631S14,理论等电点为9.04,相对分子质量为51.4 kDa,理论半衰期为30 h,不稳定系数为31.75,属于稳定蛋白,且其亲水性平均系数为-0.285。应用TMHMM在线工具分析该蛋白具有2个明显的跨膜结构域,分别位于131~153、285~307位氨基酸(见图3),表明PoFAD7蛋白为跨膜蛋白。
将PoFAD7蛋白的氨基酸序列与其他物种中已克隆的FAD7基因的氨基酸序列进行比对分析,发现该蛋白与已报道的FAD7蛋白超家族的保守结构域具有高度一致性。氨基酸序列中有3 个保守组氨酸中心位点(HDCGH、HRTHH和HVIHH)(见图4),这是维持膜结合类脂肪酸去饱和酶活性所必备的结构域[18]。另外,PoFAD7的氨基酸序列与芍药FAD7氨基酸序列相似性最高,相似度达95%。
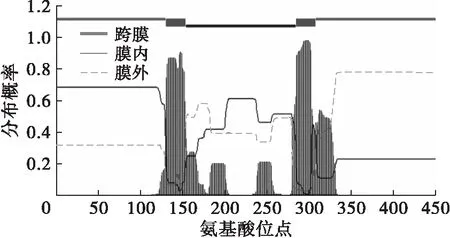
图3 凤丹牡丹PoFAD7蛋白氨基酸序列跨膜结构域预测

注:PoFAD7为凤丹牡丹FAD7;NtFAD7为烟草FAD7;GmFAD7为大豆FAD7;AtFAD7为拟南芥FAD7;BnFAD7为油菜FAD7;RcFAD7为蓖麻FAD7;JcFAD7为麻疯树FAD7;PlFAD7为芍药FAD7。组氨酸保守基序用方框标注。
图4 凤丹牡丹PoFAD7与其他物种FAD7蛋白氨基酸序列比对分析
2.3 PoFAD7蛋白的系统进化
利用MEGA5.0软件,采用邻位连接法构建PoFAD7 蛋白的系统进化树,结果见图5。
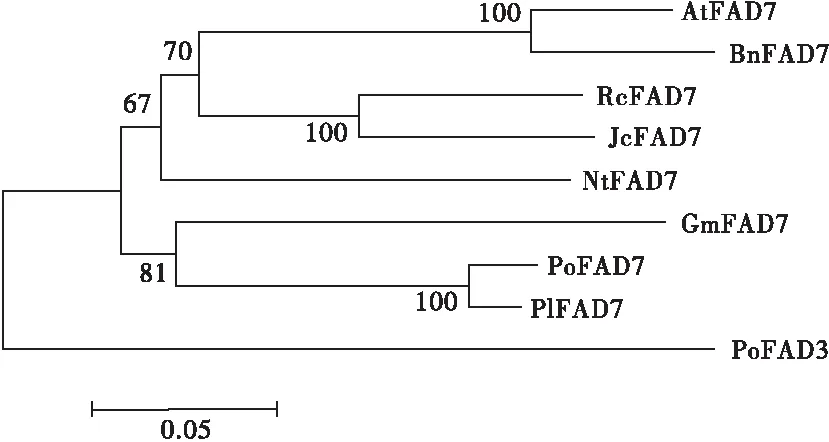
注:FAD7和FAD3蛋白来源为拟南芥AtFAD7(NP_187727);油菜BnFAD7(ACS16270);蓖麻RcFAD7(AAA73511);麻疯树JcFAD7(ABG49414);烟草NtFAD7(AIA22325);大豆GmFAD7(AEE25911);芍药PlFAD7(AKE44631);凤丹牡丹PoFAD3(ARJ54829)。
图5 凤丹牡丹PoFAD7与其他植物FAD7的系统进化树
由图5可知,PoFAD7与芍药聚为一类,亲缘关系最近,这也与凤丹牡丹和芍药同属于芍药属植物相吻合。另外,在进化树中发现PoFAD7与已克隆的凤丹牡丹PoFAD3蛋白[19]聚为不同分支,表明两者虽均属于ω-3脂肪酸脱氢酶,但功能可能不尽相同。
2.4 不同发育时期凤丹牡丹种子的亚油酸和α-亚麻酸含量
提取凤丹牡丹授粉后10、20、30、40、50、60、70、80、90、100、110、120 d的不同发育时期凤丹牡丹籽油并进行亚油酸和α-亚麻酸含量分析,结果见图6。由图6可知,从授粉30 d后凤丹牡丹种子亚油酸和α-亚麻酸含量开始急剧增加直至授粉后80 d。在种子发育后期凤丹牡丹种子α-亚麻酸含量在波动中平稳上升,而亚油酸含量大体平稳略有波动。

图6 凤丹牡丹不同发育阶段种子亚油酸和α-亚麻酸的含量
2.5 荧光定量检测PoFAD7基因表达量
利用qRT-PCR方法和PoFAD7基因的特异引物检测其在凤丹牡丹授粉后不同种子发育阶段的表达情况,结果见图7。
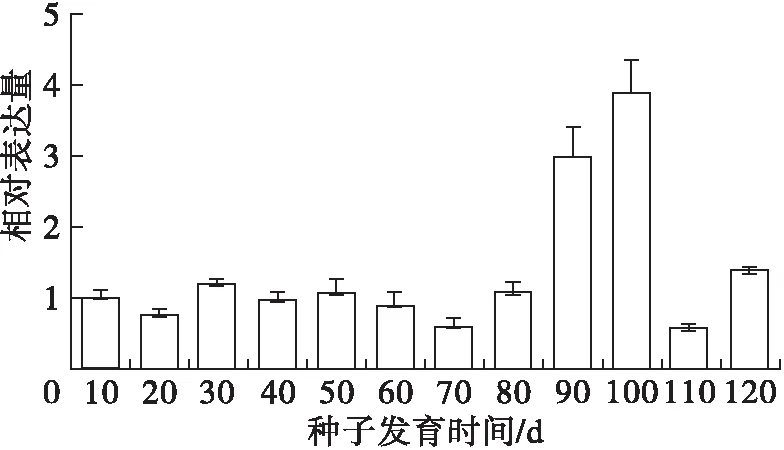
图7 凤丹牡丹PoFAD7在种子不同发育阶段的相对表达量
由图7可知,PoFAD7基因在种子发育的前80 d表达比较平稳,但在种子成熟后期的90 d急剧升高并在100 d达到最高后又下降。这与牡丹籽油中α-亚麻酸主要在授粉后30~80 d急剧累积的规律并不相符,说明PoFAD7可能并不是凤丹牡丹籽油α-亚麻酸合成的主效基因。同时此种表达模式与已报道的光皮树SwFAD7在果实中的双峰表达模式也不一致[14]。但牡丹籽油中亚油酸含量在授粉90 d和100 d时含量略有下降,这表明PoFAD7在催化亚油酸合成α-亚麻酸的反应中可能也发挥了一定的作用。
3 结 论
ω-3 脂肪酸脱氢酶是多不饱和脂肪酸亚麻酸生物合成途径的关键酶。本研究利用凤丹牡丹种子转录组数据,采用RACE和RT-PCR方法克隆获得了凤丹牡丹PoFAD7基因序列(GenBank登录号:MK205360),其包含完整的开放阅读框,推测其编码的蛋白质含有450个氨基酸。生物信息学分析表明PoFAD7蛋白具有2个跨膜结构域,含有3个保守的组氨酸基序,且系统进化树分析显示凤丹牡丹与芍药亲缘关系较近。基因表达分析表明,PoFAD7在授粉后100 d的种子中具有较高的相对表达量。结合凤丹牡丹种子中亚油酸和α-亚麻酸累积规律,推测PoFAD7在催化亚油酸合成α-亚麻酸的反应中发挥一定的功能,但可能并不是α-亚麻酸合成的主效基因。
