CFTR氯离子通道与囊性纤维化病
2019-10-23王利平
王利平
(内蒙古乌兰察布市凉城县第一中学 乌兰察布 013750)
人教版高中生物学教材有3次提到了囊性纤维化及其病因,介绍都较为简略。为什么氯离子转运异常会导致支气管中黏液增多?为什么汗液中氯离子和钠离子的浓度会升高?1987年,世界著名分子遗传学家徐立之(Lap-Chee Tsui)等[1]将囊性纤维化(CF)致病基因定位于人类的第7号染色体长臂。1989年,Riordan等[2]在寻找CF疾病基因时意外发现了囊性纤维化跨膜转导调节因子(CFTR),并且首先克隆了CFTR基因。1991年,CFTR被确定为氯离子选择性通道,从而开始了对CFTR作为氯离子通道的系统研究。
1 CFTR的结构
CFTR氯离子通道是ABC转运蛋白超家族中的一员。CFTR基因位于人类的第7号常染色体长臂(7q31.2)上,全长250 000 bp,包含27个外显子,成熟的mRNA长度为6129个碱基,其中4443为可编码序列,成熟的CFTR糖蛋白全长1480个氨基酸残基。CFTR包含5个结构域(图1),2个跨膜结构域MSD1和MSD2、 2个核苷酸结合结构域NBD1和NBD2、 1个调节结构域R。每个MSD含有6个跨膜的α螺旋,这6个螺旋组成了氯离子通道。每个NBD都含有保守的Walker A、 WalkerB和LSGGQ基序。Walker A和Walker B基序能与ATP结合,并通过ATP结合、水解调节氯离子通道的门控性。调节结构域R含有蛋白激酶A(PKA)和蛋白激酶C(PKC)所共有的磷酸化位点以及许多带电的氨基酸,当R被PKA磷酸化时,可以激活CFTR。
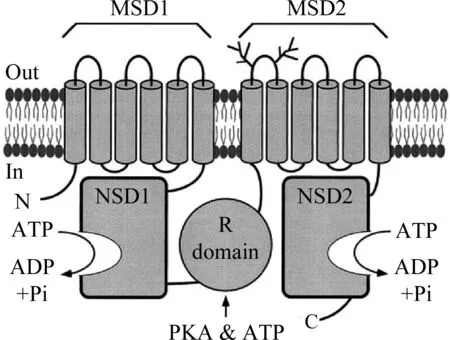
图1 CFTR氯离子通道的结构模型[3]
2 CFTR的调节机制
CFTR是一种cAMP/PKA依赖的上皮氯离子通道。允许氯离子顺电化学梯度跨膜转运。与其他ABC转运蛋白不同的是CFTR允许氯离子双向转运,而不是定向转运。当胞内氯离子浓度升髙,激活cAMP依赖的蛋白激酶最终可使通道打开,胞外的氯离子浓度升高也可以促进通道打开。MSD1和MSD2共同构成了对氯离子具有选择性的通道。MSD的部分氨基酸构成了对氯离子的选择性运输,如带有正电荷的K95、 R134、 R334、 K335、 R347和R1030在物种间具有高度保守性,它们的突变会影响到通道对氯离子的通透性。CFTR氯离子通道的门控性主要通过两个NBD来调节。NBD1和NBD2组成头尾相对的二聚体,在接触面上形成2个ATP结合位点,其中一个是由NBD1的Walker A基序与NBD2的LSGGQ基序形成的(位点1),另一个是由NBD2的Walker A基序与NBD1的LSGGQ基序形成的(位点2)。
CFTR氯离子通道开放的基本步骤是: R结构域必须先由cAMP介导的PKA磷酸化;ATP与两个NBD结合,改变CFTR立体构象,NBD1和NBD2形成紧密结合的二聚体,使CFTR氯离子通道开放,氯离子外流;结合于NBD2的ATP水解,ATP的水解和水解产物Pi的丧失使相对稳定的开路状态变得不稳定。随后,紧密结合的二聚体界面被破坏,通道关闭。
R结构域通过磷酸化和调节NBD对ATP的结合和水解能力,调控通道的门控。R结构域含有大量可被磷酸化的位点(如丝氨酸、苏氨酸等),它们可以被PKA或PKC磷酸化,同时这些位点也受到磷酸酶的调节,从而实现可逆磷酸化调节。当R结构域中的丝氨酸被磷酸化后,可以有效地促进两个NBD结构域的二聚体化,同时还增加了它们对ATP的结合和水解能力。CFTR氯离子通道的关闭则与R结构域的去磷酸化相关。
3 CFTR与囊性纤维化
CFTR广泛分布于多种组织器官,如肺、肝、胰腺、呼吸道、肠道、胆道、生殖道、心脏和血管平滑肌等。CFTR对电解质和水分的分泌和吸收发挥着非常重要的作用。
CFTR基因突变是引起囊性纤维化(CF)临床症状的分子基础。目前已报道超过1900种基因突变。常见的类型有氨基酸删除(ΔF508、ΔI507等)、氨基酸代换(错义突变)、翻译提前终止(无义突变)、插入或删除1到2个碱基而引起的移码突变,此外还有内含子碱基突变影响外显子拼接而翻译出缺陷的蛋白质。上述突变最常见的是CFTR第508位苯丙氨酸缺失(ΔF508-CFTR),引起蛋白质异常折叠,内质网相关的泛素连接酶复合体可以有效地识别ΔF508-CFTR,从而将其降解。因此ΔF508-CFTR无法到达细胞膜形成氯离子通道,即使少量到达细胞膜也无法有效打开通道,从而造成氯离子运输严重障碍。这种突变引起的后果最为严重,约占所有突变的70%,是CFTR基因突变的主要类型。多种与人类CF症状或发病机理类似的转ΔF508-CFTR基因模型和ΔF508-CFTR基因敲除模型已成功建立(如猪、雪貂、小鼠和细胞系等),以用作CF诊断、治疗和药物研发。
3.1 CF患者肺部感染及炎症 呼吸道表面附有黏液纤毛清除系统,主要由纤毛细胞、黏液层、浆液层、杯状细胞与浆液细胞组成,其中黏液层和浆液层合称为呼吸道表面液体(ASL)。ASL是抵抗病原体侵入机体的第一道防线,当有害因子吸入呼吸道时,黏附于黏液层,由上皮纤毛细胞的纤毛在浆液层中作周期性摆动,将黏附于黏液层的有害因子和异物排出呼吸道。ASL中的抗菌肽、抗氧化因子、防御素和溶菌酶等免疫因子,不仅有抗菌、抗损伤作用,而且介导炎症反应,是呼吸道免疫防御的重要环节。CF患者的一个典型特征就是呼吸道上覆黏稠、增厚的黏液,黏液不能被黏液纤毛清除系统清除,最终导致包括铜绿假单胞菌在内的少数机会性细菌对气道的定植和感染,同时机体产生特异性的抗体,抗体和细菌结合后使大量的中性粒细胞聚集于病变部位,释放大量的蛋白酶,引起免疫介导的病变部位的炎症反应,但失衡的免疫调节无法清除外在感染,反而通过多种机制进一步损伤气管结构,最终发展为不可逆的支气管扩张。在呼吸道CFTR主要表达于呼吸道上皮细胞和黏膜下腺体细胞的顶膜侧。呼吸道上皮的CFTR和上皮钠通道(ENaC)可通过控制水、盐跨膜转运来调节ASL的成分、厚度及黏液——纤毛摆动的速率。生理情况下,CFTR氯离子通道激活后,氯离子与ATP排出到呼吸道管腔,ATP又可激活外向整流氯离子通道(ORCC)和钙激活氯离子通道(CaCCs),氯离子通过CFTR、 ORCC与CaCCs等离子通道流出细胞,并且为水分子的流出提供了渗透动力。CF患者异常的CFTR不能正常分泌氯离子和ATP,上述氯离子通道受阻,氯离子在细胞内积累。在CFTR缺陷的情况下,ENaC通道被上调,呼吸道上皮ENaC功能增强,通过ENaC吸收钠离子增加,黏液中过多的钠离子转移进细胞,破坏了细胞的电解质平衡,水分从黏液中转移进细胞和血液中去,使黏液丢失水分而变得稠厚。
CFTR基因突变导致上皮氯离子分泌减少,钠离子吸收增加。然而,这些改变的离子转运过程与CF肺病之间的机制还不清楚。有关CF肺病的发病机制还有高盐假说: CF患者ASL中氯化钠的浓度高于正常人,高浓度的氯化钠使ASL中的抗菌物质失活,从而损伤了呼吸道的自我防御体系[4]。必修1教材《分子与细胞》73页提到的“1996年研究人员发现,肺部细胞外侧Na﹢的聚积会使肺易受细菌伤害”与高盐假说一致。
3.2 CF患者汗液中氯化钠浓度升高 正常情况下,汗腺分泌汗液后经由导管排泄到皮肤表面,汗液中的部分离子(如氯离子、钠离子)可由导管重吸收。人体汗液中氯化钠的浓度是由汗腺分泌和导管重吸收氯离子、钠离子的平衡决定的。汗腺主要通过两条途径分泌氯离子: 一种是CFTR,另一种是CaCCs。即使在没有CFTR的情况下,氯离子也会通过CaCCs分泌到汗液中。在汗腺导管上,CFTR和ENaC位于导管细胞的顶膜,ENaC的主要功能是重吸收汗液中的钠离子,而CFTR负责重吸收汗液中的氯离子。由于氯离子内流所造成的电化学梯度,钠离子被动地通过ENaC吸收,这减少了水和盐的损失[5]。在汗腺导管上,ENaC的活性依赖于CFTR,并随CFTR活性的增加而增强。CFTR缺陷会导致ENaC无法被激活[6, 7]。CF患者,汗腺导管细胞上CFTR功能异常,ENaC也不能被激活。汗液中氯离子重吸收明显减少,钠离子不能经ENaC内流,汗液中氯离子、钠离子浓度明显升高。
