中国茶叶生物化学研究40年
2019-10-21张梁陈琪宛晓春李大祥
张梁,陈琪,宛晓春,李大祥
安徽农业大学茶树生物学与资源利用国家重点实验室,230036
茶叶生物化学是以茶叶(茶树)为研究对象,通过现代生物化学、分子生物学、有机化学等技术手段发现、认识并掌握茶叶中特有次生代谢产物合成、降解、代谢基本原理的一门科学。茶叶生物化学研究范畴涵盖了茶叶生物体内的化学成分特点、变化规律及机制机理等一系列内容。茶树属于山茶科山茶属植物,含有特殊的次生代谢产物,如咖啡碱、茶氨酸和儿茶素等。这类物质对于茶叶品质形成至关重要,而且也是茶树生物化学研究的主要对象。
我国在1980年重新组织出版了《茶叶生物化学》一书,作为高等院校茶学相关专业的教材使用。改革开放促使现代生物化学研究水平的发展,茶叶香气的研究在气相色谱等仪器发展的基础上得到快速进步。上世纪80年代,我国开始对不同茶类的香气物质进行了分析研究,并且组织各种规模的学术研讨会,有力地推动了茶叶生物化学的交流和发展。同时,我国商业部开始组织专家对茶叶的标准进行制定,规范了茶叶的生产和品质控制。
20世纪80—90年代,国内涌现出一批研究茶叶特征成分生物合成和转化机理的学者,其研究的范围涵盖了茶树的栽培育种、制茶过程中特征成分的结构转化和机制等各个方面。这些研究逐步形成了茶学研究的主流方向,例如茶树生理学、茶叶加工化学、茶叶成分的生物合成等。另外,随着分析化学技术手段的进步,大量新型的分析方法被应用于茶叶品质成分的研究,气相色谱、液相色谱、红外光谱、紫外-可见光光谱等技术极大地丰富了茶叶生物化学研究手段,提高了茶叶生化的研究水平。
上世纪90年代至本世纪初,茶叶生物化学的发展又进入了一个新阶段。在这个阶段,除了国内学者团体的不断发展壮大,一些国外研究学者也开始茶叶生物化学的研究。随着我国科研投入的增加,国内形成了一批专门的茶叶研究机构,例如中国农业科学院茶叶研究所、安徽农业大学茶树生物学与资源利用国家重点实验室等。
随着茶树基因组的破解,以及现代生化研究方法和技术手段的发展,近年来茶叶生物化学的研究日新月异,大量高水平的研究成果不断涌现。迄今,Camellia sinensis(小叶种)和Camellia assamica(大叶种)茶树的基因组都已经绘制完成,对于其中某些标志性次生代谢产物的代谢通路关键基因也逐步清晰[1-2]。
为了能够系统地综述茶叶生物化学的最新发展前沿,笔者按照茶叶生物化学的研究对象,在前人综述论文的基础上[3-4],概述了近40年来咖啡碱、儿茶素和茶氨酸的生物合成,以及茶叶加工化学、品质化学的相关研究成果。
一、茶树主要成分的次生代谢
1.咖啡碱的生物合成
咖啡碱是一类嘌呤类生物碱,是茶叶中非常重要的一个次生代谢产物,咖啡碱含量占干重的2.50%~4.50%[5]。除了咖啡碱之外,茶叶中还含有一定量的可可碱,以及极其微量的茶碱[6]。茶树体内除种子外,其他部位均含有咖啡碱。其中,叶中含量较高,茎梗次之,花、果最少。咖啡碱呈苦味,且有较强的兴奋性,一般认为它在茶树体内扮演生物防御的作用,能够保护幼嫩组织免受害虫的伤害[7]。在茶树叶片生长过程中,咖啡碱在幼嫩叶片中含量最高,随叶片老化其含量逐渐降低。咖啡碱含量也受季节影响,夏茶中咖啡碱含量常比春茶和秋茶高。
茶树(Camellia sinensis)的咖啡碱含量高于可可碱,但是可可茶(Camellia ptilophylla)中则几乎不含有咖啡碱,却含有非常丰富的可可碱,含量可以达到6.5%左右[8]。由此可见,这些嘌呤类生物碱在山茶科的植物分布中存在着比较显著的种属差异性。我国的低咖啡碱茶树资源非常丰富,例如大坝大树茶(0.07%)、金厂大树茶(0.06%)、盐津牛寨茶(<1.0%)和厚轴茶(<1.0%)等[9]。
目前,咖啡碱的生物合成途径比较清晰,其中关键的酶也都已经基本解析,但是仍有一些合成通路的酶尚不明确。咖啡碱(1,3,7-三甲基黄嘌呤)的生物合成是以黄嘌呤核苷(XR)为底物,通过三步甲基化、一步脱核苷酸化的核心途径来实现的[10]。茶树中咖啡碱的生物合成途径主要是多个N-甲基转移酶参与甲基化而将黄嘌呤核苷转化为咖啡碱的过程,具体转化过程为:黄嘌呤核苷→7-甲基黄嘌呤核苷→7-甲基黄嘌呤→可可碱(3,7-二甲基黄嘌呤)→咖啡碱(图1),该途径也是其他含咖啡碱植物中咖啡碱的主要生物合成途径。一般所谓的咖啡碱合成酶(TCS),主要是催化最后两步的甲基化(从7-甲基黄嘌呤→3,7-二甲基黄嘌呤→1,3,7-三甲基黄嘌呤)。另外,咖啡碱的合成还有一条次要途径,通过7-甲基黄嘌呤→1,7-二甲基黄嘌呤→咖啡碱。
咖啡碱合成过程中的甲基供体是S-腺苷-L-甲硫氨酸(SAM),通过N-甲基转移酶类(NMTs)催化黄嘌呤三步甲基化,最终生物合成咖啡碱。这3种甲基化转移酶分别是黄嘌呤核苷N-甲基转移酶(7-NMT)、7-甲基黄嘌呤N-甲基转移酶(3-NMT)和3,7-甲基黄嘌呤转移酶(1-NMT),其中3-NMT的活性最高,是7-NMT及1-NMT活性总和的10倍以上。由于3-NMT和1-NMT具有几乎相同的性质,人们把这两种酶看作为同一种酶,也就是目前已经解析的TCS。

图1 咖啡碱生物合成途径
TCS基因 (CsCS1,GenBank:AB031280) 在茶树中被克隆之后,研究发现其合成酶基因存在突变。以可可茶为例,其咖啡碱合成酶(CpCS)由于个别氨基酸的突变失去了常规茶咖啡碱合成酶(TCS1)的正常功能,使可可碱(3,7-二甲基黄嘌呤)不能催化成为咖啡碱,从而导致可可碱的含量较高。茶树NMT的独立和快速的进化机制导致了TCS1具有丰富的等位变异[11]。目前,已经从茶树中发现了6种TCS1的等位基因,其中TCS1a是主要基因,其他基因存在于一些野生种茶树里面,例如不含咖啡碱的红芽茶。也正是由于TCS1序列变异多样,导致TCS1酶活性多样,从而形成了我国茶树资源中嘌呤生物碱具有不同的分布模式。Zhu等[12]利用山茶属植物咖啡碱含量不同的特点,选择了Camellia crassicolumna等低咖啡碱植物作为对照,通过转录组学等手段发现了咖啡碱降解为可可碱的途径。
次黄嘌呤核苷酸是腺嘌呤核苷酸、鸟嘌呤核苷酸及咖啡碱合成的前体物质,茶树中咖啡碱的合成可以分为核心途径和供体途径,其中合成黄嘌呤核苷有4种途径,黄嘌呤核苷酸可以继续合成咖啡碱,次黄嘌呤核苷酸脱氢酶参与了其中3种供体途径。
次黄嘌呤核苷酸脱氢酶(IMPDH)和S-腺苷甲硫氨酸合成酶(SAM)也是咖啡碱合成过程中两个关键酶,IMPDH基因在叶内表达量高于根和茎[13]。茶树次黄嘌呤核苷酸脱氢酶催化次黄嘌呤核苷酸合成黄嘌呤核苷酸,其cDNA全长序列被克隆,命名为TIDH,对其在不同组织器官中的表达也有初步研究[14]。研究发现,可以通过抑制次黄嘌呤核苷酸脱氢酶活性增加次黄嘌呤核苷酸的量来培育低咖啡碱茶树。
茶树新梢嫩叶中TCS表达量较高,与咖啡碱含量变化一致,同时检测到较强的一甲基转移酶活性,表明嫩叶中咖啡碱生物合成主要受到基因水平上的调控和底物水平控制。
茶树咖啡碱分解代谢途径通常是通过7-N-脱甲基酶介导脱去7位甲基而成为茶碱,茶碱再脱甲基成3-甲基黄嘌呤和黄嘌呤,最后经嘌呤代谢途径分解成CO2、NH3和尿素。目前,咖啡碱在茶树中的末段的生物合成途径基本清晰,但是前端的某些关键酶的基因还不能确定。7-甲基黄嘌呤核苷合成酶是催化黄嘌呤核苷成为7-甲级黄嘌呤的关键酶,但是目前这个关键酶还未被确证,其基因也不明确;是否存在3-甲基黄嘌呤直接合成可可碱的途径也尚未确证。
2.茶氨酸的生物合成
茶氨酸(Theanine)是茶叶中特有的一类非蛋白质氨基酸,化学名为5-N-乙基-γ-谷氨酰胺或γ-谷氨酰-L-乙胺。迄今为止,已经在很多山茶科植物中检出了茶氨酸,不过它在Camellia sinensis和Camellia assamica中含量较高。目前,一般认为茶树体内乙胺和谷氨酸在茶氨酸合成酶作用下生成茶氨酸[15]。
茶氨酸是茶树主体氨基酸,其存在于除果实以外的茶树各个器官,嫩叶中含量最高,其次分别是根皮、吸收根、老叶和茎等。在茶树新梢萌发前,供给茶树的氨态氮主要以茶氨酸、谷氨酰胺和精氨酸为主,这些氮源主要贮藏在根部和叶部;随着茶树的萌发,这些化合物转移到新梢,尤以茶氨酸浓度最高。
茶氨酸合成代谢途径基因包括直接参与合成的茶氨酸合成酶基因和茶氨酸水解酶基因,以及控制主要前体物乙胺来源的丙氨酸脱羧酶基因等。茶氨酸合成酶(TS)即L-谷氨酸-乙胺连接酶,是茶氨酸合成的关键酶,催化谷氨酸和乙胺合成茶氨酸。
研究表明,茶叶中的茶氨酸合成酶和谷氨酰胺合成酶(GS)具有高度的基因同源性,TS1基因与GS3基因有99%相同,而TS2基因与GS1基因有97%相同,因此推测两个基因可能源于GS家族,因为氨基酸突变导致酶学功能产生差异,使得TS具有催化谷氨酸转乙胺基合成茶氨酸功能,而普通植物中的GS只有催化谷氨酸转氨基合成谷氨酰胺的能力[16]。茶氨酸合成酶TS在ATP存在的条件下,能以L-谷氨酸和乙胺为底物催化合成茶氨酸,而作为茶氨酸组成部分的乙胺则是茶树新梢儿茶素间苯三酚核的直接前体,茶氨酸代谢通路的水解产物乙胺还参与了儿茶素的生物合成。
茶氨酸合成酶(TS)是茶氨酸合成代谢的关键酶,在ATP、Mg2+、K+存在的条件下,茶籽苗匀浆能够催化谷氨酸和乙胺合成茶氨酸。目前,宛晓春课题组从茶树基因组中找到了5条GS基因序列,分别命名为CsTSI、CsGSII-1a、CsGSII-1b、CsGSII-1c 和 CsGSII-2a[1]。其中 CsTSI具备体外合成茶氨酸的能力,与较为古老的藻类、细菌中的I型GS同源性较高,因此分类为CsTSI。 CsGSII-1a、 CsGSII-1b、 CsGSII-1c 和 CSGSII-2a均为II型GS。转基因植物研究表明,CsTSI具有双功能酶特性,即可催化谷氨酸合成谷氨酰胺,又可合成茶氨酸,而其酶学特性切换主要受底物乙胺的调控。
TS1基因在茶树芽头和根部表达几乎相当,但是TS2在茶树芽头的表达要高于根部。Liu等人研究了茶籽苗各部位茶氨酸合成酶基因的表达差异,结果表明TS1在新梢中表达量高于其他部位,根部相对较低[17]。Deng等[18]研究了咖啡碱和茶氨酸合成的特点,发现咖啡碱只在叶片和茎部合成,而茶氨酸在根部合成。
Cheng等[15]认为乙胺是茶氨酸生物合成过程中的关键前体,因为茶氨酸合成所需要的谷氨酸在很多植物中都存在,但是乙胺主要存在于山茶科的植物中,尤其是在Camellia sinensis中。乙胺作为茶氨酸的合成前体物质,在茶树根部由丙氨酸脱羧酶将丙氨酸脱去羧基生成。最近,Bai等[19]从茶树中克隆了1条新的丝氨酸脱羧酶(SDC)的基因,该基因具有很强的催化丙氨酸脱羧的作用,它在茶树根部的表达高于叶片,该基因被命名为丙氨酸脱羧酶(AlaDC)。谷氨酰胺-α-酮戊二酸氨基转移酶(GOGAT,又称谷氨酸合酶),能将GS催化生成的谷氨酰胺催化生成谷氨酸。此外,谷氨酸脱氢酶(GDH)催化α-酮戊二酸发生还原氨基化反应,生成谷氨酸。茶氨酸转运至叶部后在茶氨酸水解酶作用下降解为谷氨酸和乙胺。
茶氨酸合成酶需要K+和磷酸盐维持它的活性。在茶叶采摘后的前10 h,此水解酶活力增加,随后逐渐下降,而谷氨酰胺酶活力不断下降,采摘48 h后几乎失去活性。秋冬增施含氮基肥、采前遮阴、控制日采摘时间、喷施含茶氨酸前体叶面肥等措施可以提高鲜叶氨基酸(茶氨酸)含量。Liu等[20]关注了茶叶采摘之后在不同温度和光照条件下(遮阴)茶氨酸含量变化以及相关基因的表达差异。高温条件下,茶氨酸的含量显著降低,通过关联分析发现GOGATs在处理过程中变化较大,但是茶氨酸合成酶相关表达变化不大,加热条件下CsNADH-GOGAT表达下调,且茶氨酸含量降低。CsFd-GOGAT(铁氧还原蛋白-谷氨酰胺谷氨酸合成酶)表达水平与茶氨酸含量呈现负相关。尽管以上两个酶基因不是茶氨酸合成的关键基因,但是也属于其前体物质谷氨酸的关键合成基因[20],因此也可以通过谷氨酸的合成来调控茶氨酸的含量。
一些研究还发现气候,如气温等因素也会影响茶氨酸的生物合成。高温条件下,尤其是夏秋茶中茶氨酸的含量显著低于春茶,其主要原因可能是高温导致GOGAT等茶氨酸合成酶的活性增加,降低了茶氨酸前体的合成,从而降低茶氨酸含量[21]。
随着茶叶中茶氨酸合成途径及相关基因研究的深入,已经有一些基因工程和发酵工程技术应用于茶氨酸的生物发酵。通过构建具有茶氨酸合成能力的基因工程菌来发酵生产茶氨酸是其体外生物合成的一条有效途径[22]。
3.儿茶素的生物合成
儿茶素类化合物(Catechins)是2-苯基苯并吡喃的衍生物,属于类黄酮化合物(Flavonoids)中的黄烷-3-醇类(Flavan-3-ols)。类黄酮分子中的A环是由3个乙酸分子头尾相接而成的,而B环与C环上的碳原子则来自于由莽草酸途径合成的苯丙氨酸。苯丙氨酸经过苯丙酸盐途径形成查尔酮后,再进入各种不同的类黄酮合成途径,形成不同的黄酮类物质。这些黄酮类物质的生物合成途径具有部分的同源性。
茶树中主要多酚类物质——儿茶素类合成是一个复杂的网络途径,涉及莽草酸途径、苯丙烷类代谢途径和类黄酮合成途径,并形成多种产物(图2)。叶片中的儿茶素类化合物特别是酯型儿茶素含量远远高于根中,但后者的原花青素(聚合的儿茶素)含量是叶片的数倍,说明叶中的儿茶素类化合物后期的合成是以没食子酰基化反应为主,而根中是以原花青素聚合反应为主[23]。
植物体中类黄酮化合物的B环羟基数目受黄酮-3'-羟化酶(F3'H)和黄酮-3',5'-羟化酶(F3'5'H)基因的调控,顺式和反式儿茶素含量受花色素还原酶(ANR)和无色花色素还原酶(LAR)基因的控制,但是酯型儿茶素的没食子酰基化机理尚不完全清楚。Niemetz等[24]系统研究了水解单宁合成的相关酶类,推测1-O-没食子酰-β-葡萄糖(βG)是单宁合成有效的酰基供体和受体。利用体外酶学手段,揭示了反式儿茶素C和GC的合成模式,发现在DFR和LAR催化下,二氢黄酮醇形成无色花青素即花白素,然后形成反式儿茶素C和GC。儿茶素的合成过程中,花青素合成酶(CsANS)是一个重要的关键酶。前期研究发现光照是CsANS基因编码的一个重要调节因素。
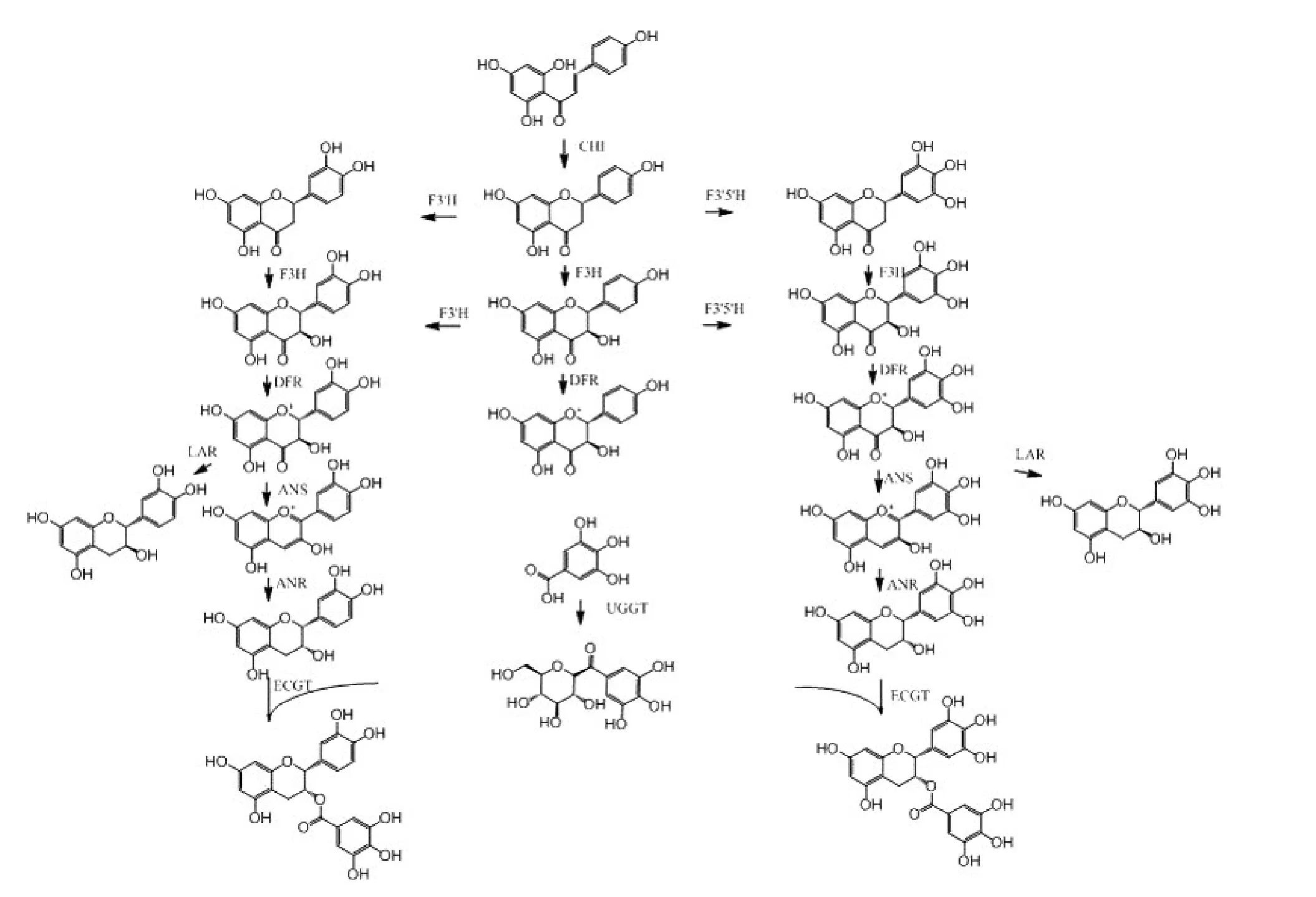
图2 儿茶素生物合成途径
Liu等[25]研究发现1-O-没食子酰基-β-D-葡萄糖转移酶(UGGT)和O-没食子酰基转移酶(ECGT)是酯型儿茶素合成过程两个关键酶。这两个酶分别以尿苷二磷酸葡萄糖和1-O-没食子酰基-β-D-葡萄糖为底物。没食子酰基化过程与水解单宁合成途径具有相似性,即βG是它们合成的酰基供体。
茶树中的黄酮醇类物质占干重的3%~4%,多以糖苷形式存在,在茶提取液的纸层析谱上就能鉴别出20多种,大部分是上述3种基本黄酮醇的各种糖苷。黄酮与黄酮醇的差别只是少1个3-羟基取代,在植物中它大多以黄酮糖苷的形式存在。
Liu等[26]利用分析化学和转录组学的手段研究春季和秋季茶树儿茶素生物合成的差异。秋茶中总儿茶素的含量显著高于春茶,其中以EGC最为突出。苯丙氨酸解氨酶(PAL)、黄酮-3-羟化酶(F3H)、黄酮-3',5'-羟化酶(F3'5'H)、二氢黄酮醇4-还原酶(DFR)和花青素合成酶(ANS)的基因表达与儿茶素的含量密切相关。Zhang等[27]也研究了黄酮代谢通路中相关基因表达与儿茶素含量的关系,结果发现查尔酮合酶1、查尔酮合酶3、花青素还原酶1、花青素还原酶2和无色花青素还原酶(LAR)的表达与儿茶素含量呈正相关,其中花青素还原酶(ANR)和无色花青素还原酶的表达与酯型儿茶素(EGCG、ECG)含量呈正相关。
近期有研究报道茶树叶片的不同位置会影响其光合作用和呼吸。叶片的成熟过程中,呼吸速率以及总氮含量会持续下降,但是淀粉的含量却会随着成熟度的增加而增加。成熟叶片中叶绿素a和b的含量都显著增加,另外,氨基酸和茶多酚的含量显著降低[28]。还有一些学者通过基因共表达网络的分析来探讨茶树3种特征性成分的相互调节关系[29],通过研究发现3类次生代谢产物的相关基因相互影响,并且相关性分析发现F3'5'H、FLS和βG不仅与EGCG的代谢相关,还与咖啡碱代谢相关。Jin等[30]最近从贵州野生茶树中发现了F3'5'H突变等位基因,其丢失了14个碱基对,从而导致F3'5'HmRNA表达水平很低,使得野生茶树中三羟基儿茶素含量较低。Tai等人[31]还比较了茶树和油茶(Camellia oleifera)在转录组水平的差异。
Yang等[32]还研究了不同氮形态对于茶叶中主要次生代谢产物合成的影响。结果发现施铵态氮(NH4+)对于茶树次生代谢产物差异基因表达的影响大于施硝态氮(NO3-)或者同时施NH4+和NO3-。长时间给予NO3-会减少黄酮类的生物合成,但是会增强咖啡碱和茶氨酸的生物合成。Fan等[33]也比较了NH4+和NO3-两种氮肥对茶树次生代谢的影响,发现Cs-miR156是调节儿茶素次生代谢途径中关键基因的一个调节基因。NO3-会增加芽头中儿茶素的含量,并且PAL、CHS、CHI和DFR的相关基因也呈现出高表达,Cs-miR156的高表达则受到NH4+的影响。Sun等[34]也试图通过生物信息学的手段挖掘与儿茶素合成基因相关的miRNAs,研究结果发现miR529d和miR156g-3p分别是CHI和F3H两种基因表达的负调控因子。
Chen等[35]还专门研究了茶树与儿茶素异构化相关基因的表达。特殊茶树品种Y510含有较高含量的GCG和C,但是其EGC和EC含量却很低。RNA序列分析发现两个影响儿茶素异构化的关键基因,花青素还原酶基因(CsANR1、CsANR2)和花青素合成酶基因。CsANS在拟南芥突变体tds4-2的过表达会导致EC含量显著增加。Li等[36]也通过比较Camellia ptilophylla和Camel-lia sinensis儿茶素合成相关基因转录水平的差异,发现Camellia ptilophyllaa的CpANS2与Camellia sinensis相似度仅有80%左右,推测其可能失去了合成cis-儿茶素的能力,从而导致白毛茶中表型儿茶素含量较低。
甲基化儿茶素是另一类具有显著生理活性的儿茶素衍生物。一般认为,甲基化儿茶素也是通过黄酮类途径合成,咖啡酰基-CoA-3-O-甲基转移酶是与甲基化儿茶素生物合成的关键酶[37]。研究发现CsWRKY31和CsWRKY48转录因子与甲基化儿茶素的合成有关,这两个转录因子的表达会抑制CsLAR、CsDFR、CCoAOMT基因的表达[38]。此外,Zhang等人[39]也发现ANL2、WRKY44和AtMYB113等转录因子也与儿茶素的合成相关。
二、加工工序对茶叶品质化学的影响
1.萎凋
陈静等[40]研究了白茶萎凋过程中茶多酚和儿茶素组分含量的变化,以及这些变化与其合成途径中关键酶基因表达的变化情况。结果发现萎凋32 h后,非酯型儿茶素EC、GC、EGC和酯型儿茶素ECG和EGCG的含量达到最高值,且这种含量变化与儿茶素合成途径中关键酶PAL、C4H、F3H、F3'H、DFR、LAR、ANR基因的表达基本一致。因此提出在白茶萎凋中,适当缩短萎凋时间,可以提高白茶品质。
刘亚峰等[41]利用通径分析研究了萎凋槽和萎凋室中不同萎凋程度对红茶化学成分与感官品质之间的关系。在以空调和除湿机控温控湿的萎凋室中,萎凋至水分含量58%~60%之间时,红茶品质最好。茶多酚、游离氨基酸、茶黄素双没食子酸酯(TFDG)、EC、EGCG和ECG与感官品质呈负相关,而茶黄素(TF1)、茶黄素-3-单没食子酸酯(TF-3-G)和茶黄素-3'-单没食子酸酯(TF-3'-G)与感官品质呈正相关,且TF-3'-G是影响红茶品质的最主要成分。
庞月兰等[42]研究了萎凋程度对红条茶品质的影响,设置了56.00%~59.99%、60.00%~63.99%和64.00%~68.00%3个萎凋程度处理,发现凌云白毫茶红条茶萎凋时间为17 h,萎凋叶含水量至60.00%~63.99%时,成品茶香气浓郁,汤色红亮,滋味浓厚,叶底红亮,茶黄素和茶红素含量均较高。
2.杀青
吴本刚等[43]以镇江金山翠芽茶鲜叶为原料,以蒸汽杀青为对照,研究了红外杀青对茶叶品质和理化成分的影响,确立了最优的杀青干燥工艺条件为:红外辐照距离20 cm杀青150 s,经揉捻做形后,热风干燥温度70℃、干燥40 min,制成品中维生素C和茶多酚保留量较高,外形色泽紧实翠绿,茶香明显。同时建立了红外杀青过程中PPO钝化动力学模型和干燥过程中水分干燥动力学模型,可为杀青干燥过程预测提供理论参考。吴雅丽[44]综述了杀青技术对改善夏秋茶品质的研究进展,指出蒸汽杀青有利于茶多酚物质和蛋白质的水解,氨基酸含量增加,有利于减少夏秋茶品质的苦涩味。
3.发酵
发酵是红茶加工中的关键过程,也是形成红茶特有品质的关键工序。Tan等[45]对工夫红茶发酵过程中(0~14 h,2 h间隔取样)的非挥发性成分进行代谢组分分析表明,儿茶素、儿茶素二聚体、黄酮醇糖苷、氨基酸、酚酸、生物碱和核苷酸等成分发生了显著变化;随着发酵时间的延长,儿茶素类、香豆素、原花青素B1和B2显著下降,咖啡碱较稳定,茶氨酸略降,而茶氨酸葡萄糖苷、杨梅素C糖苷、腺苷酸、香豆酰奎宁酸显著上升。
4.干燥
郭桂义等[46]以低档信阳毛尖毛茶为原料,研究了不同烘焙温度和时间对品质的影响。研究发现烘焙能够去除茶叶粗老气和青草气,有利于提高茶叶香气,降低茶汤苦涩味,在100℃下烘焙15 min或30 min信阳毛尖茶的品质最优,茶叶中的氨基酸、咖啡碱等物质含量较高。
陈义等[47]以不同嫩度的信阳夏茶红茶为原料,研究了50℃的炭火温度下,不同烘焙时间对茶叶感官品质的影响。研究发现炭火低温长焙能够改变信阳夏茶红茶的感官品质,提高茶叶香气,且水浸出物、茶多酚和咖啡碱含量均有不同程度的降低,从而降低了茶叶的涩味。其中一芽一叶和一芽二叶原料茶分别慢焙10.5 h,品质最佳;而一芽二三叶原料茶叶则慢焙14 h品质最佳。
张丽等[48]以水仙、肉桂2个品种的武夷岩茶毛茶为原料,经不同程度(130~150℃,2~4 h)焙火处理后。结果表明,随着焙火程度的增加,醇类含量呈降低趋势,酯类和酮类含量呈增加趋势,其中具花果香的脱氢芳樟醇、己酸叶醇酯、己酸己酯等主要香气物质含量呈先增后减的变化趋势,具烘烤香或焦糖香的香气物质(如1-乙基-1H-吡咯)呈增加趋势,苯乙腈、2,5-二甲基吡嗪、2-乙基-5-甲基吡嗪和2-乙酰基呋喃等整体呈先增后减的变化趋势。
三、展望
茶叶生物化学的研究是茶产业的基础,它将传统农业领域的茶产业提升到了涵盖第一、第二和第三产业的现代化产业。茶叶生物化学的基础研究为茶叶的品种选育、加工提升以及精深产品延伸提供了坚实的理论依据。然而,尽管近年来茶学研究的最新科技进展不断涌现,但是也存在一些研究内容雷同、研究方式类似的问题,尤其是近年来茶叶生物化学研究过程中积累的大量研究数据缺乏有效的沟通与共享。
以茶叶加工化学的研究为例,加工过程中茶叶中主要物质的变化规律受到许多因素的影响,例如基础材料(鲜叶)的成分差异,成分之间的相互作用,加工条件和参数等因素的影响,这类加工化学的研究往往都具有一定的个体性,而难以代表不同茶类,甚至同类茶叶的规律。因此,以茶叶主要内含物质为基础,建立标准化的组合化学物,并以其为基础模拟茶叶加工过程中热、湿度、pH值、酶促反应、单一/组合微生物发酵等条件,探究茶叶成分的变化规律,相互作用特点,建立可重复、可参考的茶叶化学转化模式,从而揭示茶叶加工过程中的生化变化特点。此外,国内多个科研院所都分别建立了茶叶相关数据,涵盖了茶叶研究的各个方面,存在着一定的重复建设和资源浪费。因此,我们也亟待建立区域间协作,甚至是国际范围内的茶叶生物(基因)和化学数据库,供全球茶叶研究者和公众使用。
