喀喇昆仑-帕米尔地区牦牛mtDNA Cytb遗传多样性及系统发育分析
2019-10-14韩建林王玉涛
李 静, 王 莉, 道 敏, 胡 平, 肖 逸, 韩建林, 王玉涛*
1.喀什大学生命与地理科学学院, 新疆维吾尔自治区叶尔羌绿洲生态与生物资源研究高校重点实验室, 新疆 喀什 844000;2.新疆维吾尔自治区塔什库尔干县畜牧兽医局, 新疆 塔什库尔干县 845250
牦牛(Bosgrunniens),属哺乳纲(Mammalia),偶蹄目(Artiodactyla),反刍亚目(Ruminantia),牛科(Bovidae)[1],主要分布在中国青藏高原以及与其相邻近的亚高原地区[2~4],中国牦牛是世界牦牛数量最多[5~7]、品种资源最丰富的国家[8,9],中国牛品种志收录的地方牦牛品种有5个[10],现国家鉴定的牦牛品种(类群)18个,西藏藏族自治区主要的牦牛品种为帕里牦牛、嘉黎牦牛和斯布牦牛,青海省主要分布环湖牦牛和高原牦牛,四川省主要牦牛品种为九龙牦牛和麦洼牦牛,甘肃省主要牦牛品种为天祝牦牛和甘南牦牛,云南省牦牛品种为中甸牦牛,新疆维吾尔自治区已鉴定的牦牛品种为巴州牦牛,2014年后又陆续鉴定出四川省木里牦牛、金川牦牛、西藏藏族自治区类乌齐牦牛、青海省雪多牦牛、四川省昌台牦牛;培育品种两个:包括青海省大通牦牛[11]和阿什旦牦牛[12~14],而以上品种资源均未涵盖分布于新疆喀喇昆仑-帕米尔地区的牦牛群体,是地方牦牛品种资源认定分布上未涉及的种群。
牦牛几乎是集各种家畜产品优势[15]、性能于一身的多功能动物[16~18],是我国高寒地区的主要畜种和重要的生产资料[9,19,21]。对牦牛遗传多样性的研究有助于了解牦牛的分类地位及其品种的遗传多样性,本文对牦牛地方种群现状进行了分子遗传学分析,对牦牛资源的合理保护与科学利用具有重要意义。
在研究动物遗传多样性分子标记中,哺乳动物线粒体细胞色素b(Cytb)基因是线粒体DNA中重要的蛋白编码基因,它包含从种内到种间乃至到科间的进化遗传信息[21],因而被广泛地用来进行脊椎动物种上和种下系统进化研究[22],被认为是解决系统发育问题最可信的线粒体DNA标记之一。常国斌[23]对20头巴州牦牛进行研究分析发现,巴州牦牛核苷酸多样性相对贫乏,单倍型多样性相对较丰富,推测巴州牦牛可能存在不同的母系起源。王兰萍等[24]在分析巴州牦牛时发现,一部分巴州牦牛含野牦牛血统,推测可能由于这部分牦牛与野牦牛有共同祖先,在漫长进化过程中仍保留相似序列。涂世英等[25]研究中甸牦牛发现,中甸牦牛核苷酸多样性较贫乏,而单倍型多样性较丰富,推测中甸牦牛可能存在不同的母系起源,系统发育树结果表明中甸牦牛与野牦牛亲缘关系较近,其次是美洲野牛,推测中甸牦牛在发展的过程中与野牦牛发生了基因交流,作者支持牦牛属于牛亚科牦牛属的观点。胡丹[26]对新疆塔什库尔干县牦牛10个个体的mtDNA D-loop序列和mtDNACytb进行分析,表明塔什库尔干县牦牛具有丰富的遗传多样性,支持牦牛和野牦牛划分为牛亚科牦牛属的观点,但该研究采集样本少且地理覆盖率低。以上研究均未对分布在喀喇昆仑-帕米尔地区的牦牛群体进行深入研究,因此该地区牦牛遗传多样性、遗传分化及系统地位尚不清楚。
牦牛在牛亚科中的分类地位一直是牛亚科动物遗传发育领域的热点问题,牦牛是隶属于牛亚科中独立的牦牛属,还是与北美野牛隶属于野牛属,或是作为牛亚科牛属牦牛亚属仍存在热议,到目前为止仍没有确定的结论。姬秋梅[27]、李齐发[28]构建系统发育树发现,牦牛与野牦牛、美洲野牛亲缘关系较近,与牛属关系较远,李齐发[28]推测家牦牛与野牦牛的分化时间约为55万年前。杨万远[29]对野牦牛分析发现,野牦牛具有丰富的遗传多样性,支持将牦牛划分为牛亚科中一个独立的牦牛属,李齐发[28]也认为家牦牛和野牦牛应划分为牛亚科独立的牦牛属。但常国斌[23]、钱建新[30]基于水牛属的系统发育地位,认为牦牛应归为牛属中牦牛亚属更合理。耿荣庆[31]牛亚科动物亲缘关系的研究中发现,大额牛与瘤牛之间可能有共同的母系起源;家牛与准野牛属亲缘关系相近、其次是牦牛属,认为牛亚科动物应划分为4个属,包括家牛属、准野牛属、牦牛属和亚洲水牛属。
喀喇昆仑-帕米尔地区海拔高、常年积雪、气候干旱、地势险峻,这里形成了天然的地理隔离带[32,33],从而造成牦牛之间基因交流少,群体退化较为严重,物种资源保护与利用亟待解决。为进一步明确喀喇昆仑-帕米尔地区牦牛遗传多样性水平、遗传分化和系统进化地位,本文以分布在该区域的牦牛为研究对象,选择mtDNACytb遗传标记,利用PCR直接测序和生物信息学方法,研究喀喇昆仑-帕米尔地区牦牛mtDNACytb序列特征和遗传多样性水平,选择GenBank中已提交的野牦牛及不同品种家牦牛序列,利用最大似然法构建系统进化树,这将为该区域牦牛遗传资源保护和利用提供理论依据;利用牛亚科代表物种序列进行系统发育分析,研究牦牛在牛亚科中的分类地位。
1 材料与方法
1.1 实验材料
本实验采集覆盖疆喀喇昆仑-帕米尔地区5个地方(塔什库尔干县、乌恰县、阿克陶县、阿合奇县、叶城县)牦牛群体血样或肌肉组织样本共计110个(表1)。血液样本采取颈静脉采血,ACD抗凝剂抗凝,-80℃超低温保存;肌肉组织样品取屠宰厂牦牛背最长肌,编号后冷冻保存,运输途中保持低温,储存于喀什大学叶尔羌绿洲生态与生物资源研究高校重点实验室-80℃冰箱,取部分样品在100%乙醇中固定,分样保存。
1.2 基因组DNA提取
血液和肌肉组织DNA提取方法参考经典的苯酚-氯仿抽提法,在此基础上,由于血液、肌肉组织中蛋白质含量较多,本实验中对饱和酚、氯仿-异戊醇处理步骤适当重复及处理时间适当增加。
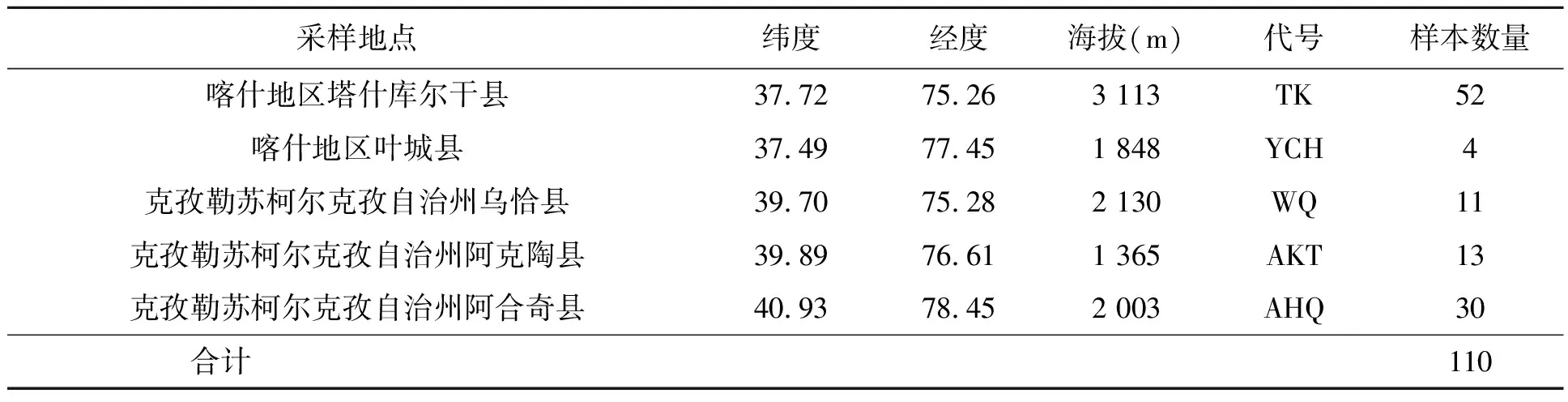
表1 牦牛样本采集的样品信息Table 1 Sample information collected in this study.
1.3 引物设计和PCR扩增
引物设计与合成:以GenBank数据库中已提交的家牦牛的mtDNA序列(GenBank登陆号:GQ464270.1、JQ692071.1)为参考序列,利用Primer Premier 5.0软件进行引物设计,引物序列为:YAK mtDNAcytb-F:5′-GTTCCGTAGCCATAGCCG-3′, YAK mtDNAcytb-R: 5′-TTGAGTCTTAGGGAGGTT-3′。PCR目的产物的大小约为1 520 bp,引物由苏州金唯智生物科技有限公司合成。
PCR反应体系:总体积50 μL,其中2×TaqPCR Mix 26 μL,上、下游引物(10 pmol/μL)各1 μL,DNA模板(100 ng/μL) 2 μL,ddH2O 20 μL。扩增反应条件:95℃预变性5 min;94℃变性30 s,51.5℃退火30 s,72℃延伸80 s,40个循环;72℃延伸10 min,4℃保存。PCR产物经2%的琼脂糖凝胶电泳检测,扩增产物送苏州金唯智生物科技有限公司测序。
1.4 序列分析
通过Chromasversion 2.6.6原始序列峰图数据进行人工校对。利用Clustal X软件对所测定的mtDNACytb序列进行同源序列比对,用MEGA 5.2、MEGA 6.06统计序列碱基组成分析,用DNASP 5.10.01软件进行单倍型的推导,并开展遗传多样性的分析;最后将获得的单倍型数据用MEGA 6.06软件进行系统发育分析,采用软件NETWORK 4.1.0.9进行单倍型的网络结构和频率分布分析。从GenBank数据库下载已提交的野牦牛及中国不同品种牦牛mtDNACytb的同源序列共82条,这些序列的来源信息见表2,同时下载42条牛亚科代表物种及绵羊、犀牛序列,序列的来源信息见表3,将GenBank数据库中下载的序列进行比对剪切为1 400 bp,便于后续数据分析。
2 结果与分析
2.1 PCR扩增
对110个喀喇昆仑-帕米尔地区牦牛mtDNACytb区进行扩增,用2.0%的琼脂糖凝胶电泳进行检测(图1)。扩增产物条带质量较好,条带清晰、明亮、单一,无非特异性扩增产物,符合后续的PCR产物测序要求。
2.2 牦牛mtDNA Cytb序列分析
2.2.1牦牛mtDNACytb核苷酸组成 测序结果与参照序列GQ464270.1进行比对、人工矫正后,将测序获得的1 150 bp序列与参照序列GQ464270.1、JQ692071.1进行对比和校对后,最后获得长约1 140 bp的核苷酸序列用于进一步分析。

表2 从GenBank 下载的牦牛mtDNA Cytb 序列信息Table 2 Information of mtDNA Cytb sequences in yak retrieved from GenBank.

表3 从GenBank 下载的牛亚科代表物种mtDNA Cytb 序列信息Table 3 Information of mtDNA Cytb sequences of bovine subfamily representative species from GenBank.
图1 mtDNA Cytb全长扩增结果Fig.1 Results of full length amplification of mitochondrial mtDNA Cytb.
利用Clustal X 软件对所测定的mtDNACytb序列进行同源序列比对后,通过MEGA5.2对mtDNACytb全序列长度和碱基组成进行分析(表4)。由表中可以看出这6个牦牛群体mtDNACytb中T、C、A和G 4种碱基的平均比例分别为26.2%(26.1%~26.2%)、29.0%(28.0%~29.1%)、31.7%和13.1%,在不同种群之间,mtDNACytb序列碱基差异不明显。

表4 5个牦牛群体mtDNA Cytb序列(1 140 bp)的碱基组成Table 4 Nucleotide composition in the mtDNA Cytb sequences (1 140bp) of five populations of yak
2.2.2喀喇昆仑-帕米尔地区牦牛mtDNACytb多态位点分析 用DNASP 5.10.01软件对喀喇昆仑-帕米尔地区5个牦牛种群的110个样品的mtDNACytb全序列分析共检测出多态位点10个,界定出单倍型5个(表5,图2)。其中H1、H4为特有单倍型,单倍型H1为阿合奇县牦牛特有单倍型包含4个个体;单倍型H4为乌恰县牦牛特有单倍型仅有2个个体;单倍型H2、单倍型H5为优势单倍型,5个牦牛群体均包含单倍型H2,共35个个体,单倍型频率为31.82%;5个牦牛群体也均含有单倍型H5,共65个个体,单倍型频率为59.09%;单倍型H3为3个牦牛群体共享单倍型,共有6个个体含1个阿克陶县牦牛、1个乌恰县牦牛、4个塔什库尔干县牦牛。
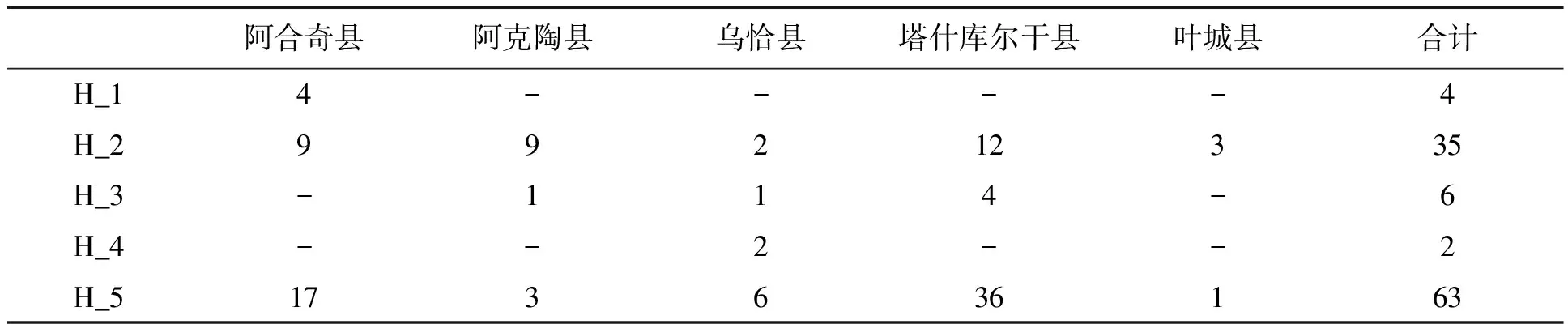
表5 5个牦牛群体中5个线粒体mtDNA Cytb单倍型(1 140 bp )在各群体中的分布Table 5 Distribution of five mitochondrial mtDNA Cytb haplotypes (1 140 bp) in five yak populations.
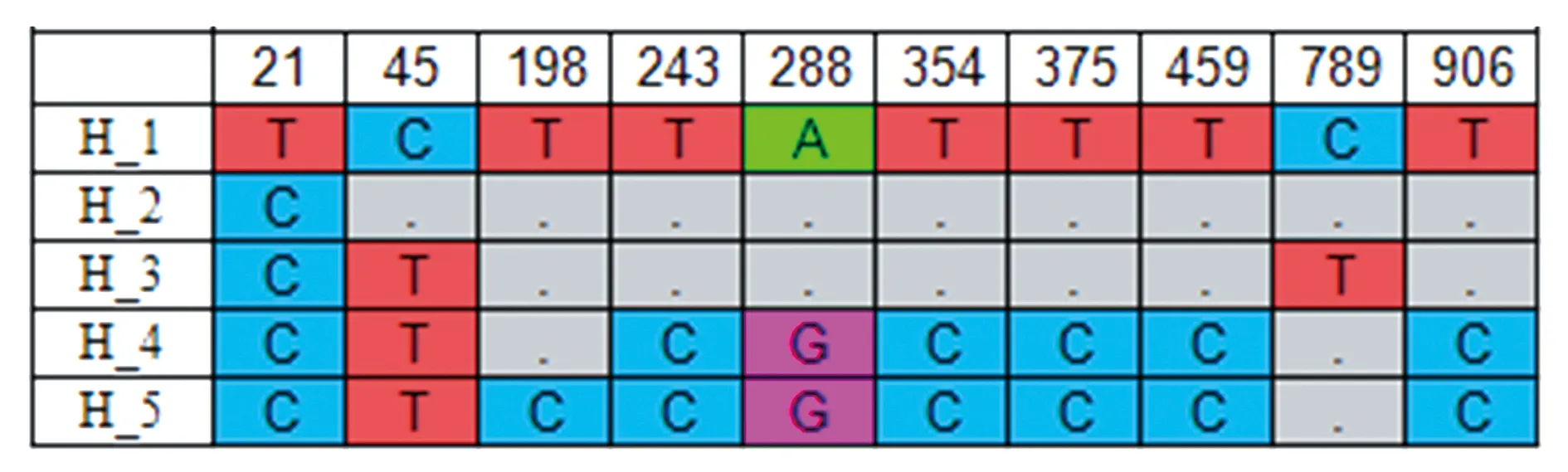
图2 5个牦牛群体5个线粒体mtDNA Cytb单倍型(1 140 bp )中10个多态位点的分布Fig.2 Distribution of 10 polymorphic loci in 5 mitochondrial mtDNA Cytb haplotypes (1 140 bp) in 5 yak populations.
2.2.3喀喇昆仑-帕米尔地区mtDNACytb核苷酸多样性和单倍型多样性分析 用DNASP5.10.01软件对喀喇昆仑-帕米尔地区牦牛5个牦牛群体的110个个体的单倍型多样性和核苷酸多样性进行系统分析(表6),发现总的平均单倍型多样性(Hd)为0.566±0.001 23,平均核苷酸多样性(π)为0.002 95,平均核苷酸差异性(K)为3.363。
2.2.4基于mtDNACytb序列分析牦牛在牛亚科中的分类地位 为了揭示喀喇昆仑-帕米尔地区牦牛群体与中国地方牦牛品种的差异情况和遗传

表6 5个牦牛群体mtDNA Cytb 序列遗传多样性相关参数的分析结果Table 6 Results of genetic diversity parameters of mtDNA Cytb in the five populations of yak.
关系,为喀喇昆仑-帕米尔地区牦牛的品种鉴定提供理论基础,我们进一步对提交到GenBank数据库中的不同地区牦牛品种的mtDNACytb序列进行比对分析和聚类分析(图3,表7,图4)。
通过DNASP 5.10.01软件对喀喇昆仑-帕米尔地区牦牛与野牦牛及中国10个牦牛品种和西藏地区牦牛mtDNACytb(1140 bp)共82条序列检测出35个多态位点,共获得24个单倍型(表7,图4)。单倍型H2、H3、H4、H5为优势单倍型,其中单倍型H2含有97个个体,占个体总数的50%;每个家牦牛品种中均含有1个特有单倍型,野牦牛含有7个特有单倍型;基于各种群间的遗传距离构建系统进化树显示喀喇昆仑-帕米尔地区牦牛与野牦牛亲缘关系较近,与巴州牦牛亲缘关系较远(图3)。
2.2.5基于mtDNACytb序列分析牦牛在牛亚科中的分类地位 为了明确牦牛在牛亚科中的分类地位,我们进一步对提交到GenBank数据库中的牦牛mtDNACytb序列进行了比对,建树分析牦牛的分类地位,将110个喀喇昆仑-帕米尔地区牦牛和下载的82条牦牛序列进行单倍型分析后,将牦牛形成的24个单倍型与牛亚科代表物种42个序列构建系统发育树(图5),结果表明:牦牛首先和美洲野牛聚类,形成的分支与欧洲野牛聚类,再与印度野牛、大额牛、林牛、爪哇野牛及普通牛与原牛、瘤牛形成的分支聚为一类,最后和水牛属相聚。
3 讨论
本研究测定mtDNACytb基因全长为1 140 bp,牦牛mtDNACytb基因不存在长度变异;T、C、A和G碱基的平均含量分别为26.2%、29.0%、31.7%、13.1%,其中A+T碱基含量(57.9%)大于C+G碱基的含量(42.1%),表现出了碱基的偏倚性;共发现10个多态位点:均为转换,与胡丹[26]研究10头塔什库尔干县牦牛分析共得到3个SNP 位点变异类型结果一致,而与涂世英[25]分析中甸牦牛mtDNACytb基因发现3个多态位点及杨万远[29]对 6头野牦牛 mtDNACytb基因研究发现 13 处变异位点的变异类型(转换、颠换)存在差异;表明喀喇昆仑-帕米尔地区牦牛mtDNACytb基因SNP 位点较丰富,不同牦牛品种的SNP位点变异类型存在差异。

表7 12个牦牛群体24个mtDNA Cytb 单倍型(1 140 bp )在各群体中的分布Table 7 Distribution of 24 mtDNA Cytb haplotypes (1 140 bp) from 12 yak populations in each population.

图4 12个牦牛群体24个线粒体mtDNA Cytb单倍型(1 140 bp )中35个多态位点的分布Fig.4 Distribution of 35 polymorphic loci in 24 mitochondrial mtDNA Cytb haplotypes (1 140 bp) from 12 yak populations.

图5 牛亚科代表物种间mtDNA Cytb序列系统进化发育聚类图Fig.5 Clustering gram of development and evolution between species of bovine subfamily based on mtDNA Cytb sequence.
本研究喀喇昆仑-帕米尔地区110个牦牛中共界定出5个单倍型,平均单倍型多样性(Hd)为0.566±0.001 23,平均核苷酸多样性(π)为0.002 95,平均核苷酸差异性(K)为3.363。与涂世英[25]研究的中甸牦牛、常洪等[24]分析的巴州牦牛、胡丹[26]塔什库尔干县牦牛mtDNACytb的研究结果相比,平均单倍型多样性(Hd)、平均核苷酸多样性(π)均较高;但与姬秋梅[27]研究测定的西藏牦牛mtDNACytb基因相比平均单倍型多样性(Hd)、平均核苷酸多样性(π)均较低。说明西藏牦牛遗传多样性最为丰富,喀喇昆仑-帕米尔地区牦牛遗传多样性较为丰富,而中甸牦牛、巴州牦牛遗传多样性较弱;西藏牦牛丰富的遗传多样性与西藏是牦牛起源的观点相符;喀喇昆仑-帕米尔地区牦牛遗传多样性较为丰富,但基于喀喇昆仑-帕米尔地区为天然的地理隔离带,推测可能是由于最初迁徙至喀喇昆仑-帕米尔地区的牦牛含不同起源有关。
本研究中牦牛与美洲野牛亲缘关系较近,与普通牛、原牛及瘤牛亲缘关系较远。前期基于mtDNA D-loop区序列研究,郭松长[34]认为牦牛应与草原野牛、美洲野牛合为一个属,而不是独立为牦牛属;李齐发[1]、钟金城[35]支持将牦牛、野牦牛划分为牛亚科中牦牛属的观点;基于野牦牛线粒体全基因组研究,钟金城[36]认为牦牛属应为牛亚科中的一个独立属;基于mtDNACytb区序列研究,李齐发[26]也认为家牦牛和野牦牛应划分为牛亚科独立的牦牛属;常国斌[23]、钱建新[30]基于水牛属的系统发育地位,认为牦牛应归为牛属中牦牛亚属更合理。本研究认为牦牛应属于牛亚科中独立的牦牛亚属。
本研究首次以喀喇昆仑-帕米尔地区牦牛为研究对象,对喀喇昆仑-帕米尔地区牦牛110个个体的mtDNACytb基因进行了测序,序列长均为1 140 bp,形成5种单倍型,含10个多态位点,核苷酸变异类型仅为转换,平均单倍型多样性(Hd)、平均核苷酸多样性(π)较高,其群体变异程度高,遗传多样性较为丰富,系统进化树表明与野牦牛亲缘关系较近,与其他地方品种牦牛亲缘关系较远。
喀喇昆仑-帕米尔地区牦牛遗传背景独特,具有作为我国优良地方牦牛品种的遗传资源的潜质,应加大对喀喇昆仑-帕米尔地区牦牛品种优良资源的保护。
