不同体力活动水平大学生的生物运动工作记忆加工特征研究
2019-10-08娄莹周成林陆颖之
娄莹 周成林 陆颖之
摘 要:以生物运动光点视频为刺激方式,探讨不同体力活动水平大学生生物运动工作记忆加工的差异性特征。方法:通过64导脑电设备记录18名高体力活动水平大学生(高体力活动组)和19名低体力活动水平大学生(低体力活动组)在生物运动工作记忆任务中的脑电(EEG)变化情况,在这一任务中的正确率和反应时作为行为评价指标,以alpha频段能量、theta频段能量和theta频段能量/alpha频段能量作为脑电指标。结果:高体力活动组的行为指标优于低体力活动组,正确率组间差异显著。高体力活动组在alpha频段能量上显著高于低体力活动组,并且在额区和中央区满足高记忆负荷下的alpha频段能量大于低记忆负荷。theta频段能量和theta频段能量/alpha频段能量均存在组别与脑区的交互作用,且2组被试均满足大脑额区>大脑中央区>大脑顶区>大脑枕区这一规律。结论:在进行生物运动工作记忆加工过程中,高体力活动水平的大学生有着更高的大脑唤醒水平和更好的行为表现。
关键词:生物运动;体力活动;工作记忆
中图分类号:G 804.8 学科代码:040302 文献标识码:A
Abstract:Objective: Point light displays were used as the stimulating approach to explore the differences between different levels of physical activity in biological motion working memory of university students. Method: The electroencephalogram(EEG) data in biological motion working memory task were recorded in 19 participants with low physical activity and 18 participants high physical activity. Behavioral data of the participants were recorded including correct rate and response time were used as evaluation indicators of cognitive behavior. In terms of EEG, alpha(8-13 Hz), theta(4-7 Hz) and theta/alpha were used as indicators for EEG changes. Results: The behavior of the high physical activity group was better than that of the low physical activity group, and there was a significant difference in the correct rate between the groups. The alpha energy value of the high physical activity group was significantly higher than that of the low physical activity group, and the alpha energy value under the high memory load condition was significantly higher than that of the low memory load condition in the frontal and central regions. Both theta energy value and theta/alpha ratio had interaction between groups and brain regions, and both groups met the rule of frontal area > central area > parietal area > occipital area. Conclusion: Individuals in the high physical activity group have greater levels of brain activation and better cognitive behavior performance than those in the low physical activity group.
Keywords:biological motion; physical activity; working memory
在日常生活中,如何正確地认识和理解环境中生物的运动和行为,是个体与他人进行交流和互动的重要前提。而对生物运动信息的加工是人们与环境及他人进行互动的重要内容之一,通过对生物运动的感知和加工,人们能够在动态的自然环境中,推断生物运动的意图和目的,进而形成互动[1]。近年来,研究者从行为层面对生物运动信息的加工特征进行了探讨,发现了大量可能影响生物运动信息加工的因素[2],而除了认知风格等内部因素外,外部因素(如体力活动等)是否也有可能影响个体对生物运动信息的认知加工呢?
随着全民健身活动的开展,研究者越来越多地对体力活动促进认知加工的作用机制进行探索[3-4]。较高水平的体力活动有利于个体认知加工能力(工作记忆、注意转换、抑制控制等)的发展和保持。工作记忆作为执行功能的重要组成部分,是人们完成运算、推理、语言理解等高级认知活动的基础[5]。研究发现,高水平体力活动能有效延缓老年人工作记忆的衰退[6-7],也能促进学龄前儿童工作记忆能力的发展[8]。目前,尚未有研究对体力活动与生物运动工作记忆之间的关系进行探讨。
与一般的视觉客体工作记忆不同,生物运动工作记忆有其相对独立的加工和存储机制,例如其在工作记忆中的复述需镜像神经元的参与,同时,工作记忆加工生物运动信息相关的过程也有别于一般视觉客体[9]。对生物运动信息进行加工的能力被认为是衡量个体社会认知水平的关键指标,例如研究发现,生物运动工作记忆的负荷可以预测个体的共情水平,这是视空间及视觉客体工作记忆无法实现的[10]。体力活动对视空间工作记忆的促进作用得到了大量研究证实,较高水平的体力活动和参与较多种类的体力活动都能够有效提升人们运动知觉的敏感性,而运动知觉敏感性是提高个体生物运动知觉的关键因素[11]。尽管高水平体力活动对生物运动知觉有积极的促进作用,但这些结果并不适合直接推论为高水平体力活动对生物运动工作记忆产生的影响;因此,本研究将针对生物运动工作记忆,探索不同的体力活动水平对其产生的影响。
目前,在有关生物运动知觉和工作记忆加工的神经机制的研究中,脑电(EEG)技术及与工作记忆相关的评价指标被广泛地使用。theta频段(4~7 Hz)的能量反映了工作记忆的信息编码过程与表现过程[12-13],并与大脑认知负荷呈正相关关系[14]。Neymotin等[15]研究视觉工作记忆的EEG信号时发现,theta频段的能量随认知负荷的增加而提高,而alpha频段(8~13 Hz)的能量被认为是与大脑皮质的激活或觉醒有关[16-17],它与抑制性的加工呈正相关关系[18]。为了减少个体差异及不同认知任务带来的影响,近年来,研究人员开始采用theta频段能量与alpha频段能量的比率评估工作记忆负荷对认知加工的影响[13,19],并且发现,theta频段能量/alpha频段能量会随着工作记忆难度的增大而提升[20]。因此,本研究中分别以theta频段能量、alpha频段能量及theta频段能量/alpha频段能量作为评判指标,探讨不同生物运动工作记忆负荷对大脑认知加工的影响。
综上所述,体力活动可以有效改善个体的认知水平,但这种认知水平的改善是否也会体现在生物运动工作记忆的加工过程中,目前尚不确定。因此,本研究将探索不同体力活动水平成年人生物运动工作记忆的加工特征。选取不同体力活动水平的大学生,使用“光点视频”(PLD)记录和比较其在不同工作记忆负荷下的大脑活动情况,揭示不同水平体力活动与生物运动工作记忆之间的关系。本研究假设:高水平體力活动个体在较高生物运动工作记忆负荷下,消耗的认知资源更少,效率更高,但体力活动水平不影响个体生物运动工作记忆的负荷。
1 研究对象与方法
1.1 研究对象
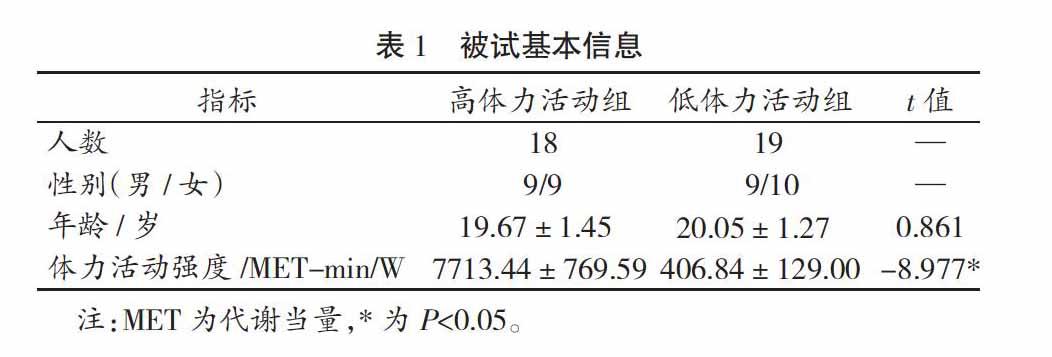
研究共招募某高校大学生54人,根据国际体力活动调查问卷(IPAQ),以高强度体力活动≥3 d/W,且每周总体力活动≥3 500 MET-min/W的标准筛选出18人,组成为高体力活动组。其中,男性9人,年龄为(20.05±1.27)岁。同时以有无规律性的高强度体力活动且每周总体力活动<600 MET-min/W的标准筛选出19人,组成低体力活动组[21]。其中,男性9人,年龄为(19.67±1.45)岁。被试基本信息见表1,所有被试视力正常或矫正视力正常,无色盲。被试均自愿参与本研究,并签署知情同意书。
1.2 实验设计
本实验为两因素混合实验设计,组别(高体力活动组和低体力活动组)为被试间变量,工作记忆负荷(低等、中等、高等)为被试内变量。因变量包括行为反应(反应时间、反应正确率)和大脑活动情况(alpha频段能量、theta频段能量及theta频段能量/alpha频段能量)。
1.3 实验仪器及设备
本实验设备包括1台安装运行程序的笔记本电脑和1台用于刺激呈现的笔记本电脑,其分辨率为1 024×768,刷新频率为100 Hz。被试的脑电采集使用德国Brain Products公司的记录系统,采样率为500 Hz。64个电极点按照国际10-20标准分布于脑电帽,佩戴于被试头部完成脑电采集。
1.4 实验材料
本实验以经典的PLD[22]为识记资料,在来自于 Vanire和Verfaillie的数据库[23]中,选取18个时长为1 s的PLD,每1种PLD皆由13个光点构成。实验中刺激均在电脑显示器上呈现,屏幕背景为黑色,被试眼睛距离电脑屏幕大约60 cm。
1.5 实验过程
实验采用生物运动工作记忆的变化觉察任务(the change detection task),实验流程如图1所示,本实验在经典的变化觉察范式基础上添加了同步的数字复述任务,以防止被试使用语音编码策略[24]。呈现3个随机数字,要求被试大声复述,时长为500 ms,随后呈现一个注视点,呈现时长为1 000 ms,提醒被试将进行PLD记忆任务。在记忆项部分:屏幕上会出现2~4个PLD,呈现的时长分别为2 000、3 000或4 000 ms。2 000 ms的注视点后,呈现随机PLD的检测项。随后出现1个红色问号,要求被试判断检测项的PLD是否在记忆项中出现过,若出现过,按J键,反之,则按F键。按键后,出现1 000 ms时长的注视点,最后呈现1个数字,要求被试判断该数字是否在数字复述任务中出现过,按X键判断为“是”,按Z键判断为“否”,2个判断均需在1 000 ms内完成。
实验共3组,每组30个试次。3组任务难度由低到高,分别为2个PLD、3个PLD和4个PLD。所有任务呈现与控制均通过Matlab2014a及Psychtoolbox工具包实现。正式实验前,被试进行30个试次的练习(练习试次为2个PLD),确保被试完全了解实验流程后正式开始实验,被试在每组任务之间有3 min休息时间,整个实验共90 min。
1.6 数据采集与分析
1.6.1 行为数据采集与分析
行为数据由Matlab2014a软件记录采集,分析指标为被试判断光点视频的反应时和正确率及数字任务正确率;通过IBM SPSS Statistics 19软件,使用重复测量方差分析对2组数据进行处理。
1.6.2 EEG数据采集与分析
脑电数据采集将FCz电极点作为参考电极,AFz电极点作为接地电极在线采集。分别放置右眼眶外侧水平眼电(HEOG)、左眼眶正下方垂直眼电(VEOG),并确保头皮与电极之间的阻抗小于10 kΩ。根据被试情况调整显示高度,并在数据采集过程中,要求被试双脚平放,并尽量减少肩部和头部活动,保持眼睛与显示器中心处于同一高度。
在线采集脑电数据后,采用Brain Production Analyzer 2.1软件进行离线分析。首先进行重参考,选择双耳乳突(TP9和TP10)为新参考电位,并将参考电极FCz点还原为记录电极。以低通70 Hz,高通0.3 Hz,斜度24 dB/oct为标准进行带通滤波,之后通过独立成分分析进行半自动去眼电伪迹,将各组的EEG数据以1 s为单位进行分段,分段后通过快速傅立叶变换将其从时域信息转为频域信息。在对数据进行基线校正后,进行叠加,并计算得到alpha频段能量、theta频段能量和theta频段能量/alpha频段能量。
根据已有研究及脑地形图,分别选取大脑额区(F1、F2、Fz)、大脑中央区(C1、C2、Cz)、大脑顶区(P1、P2、Pz)及大脑枕区(O1、O2、Oz)4个脑区,对各脑区的电极点进行平均以计算各个脑区的alpha频段能量、theta频段能量及theta频段能量/alpha频段能量。进行2(组别)×3(2PLD、3PLD、4PLD)×4(F区、C区、P区、O区)的重复测量方差分析。对不满足球形检验的统计量采用Greenhouse-Geisser法矫正自由度和P值,事后比较采用Bonferroni法进行校正,本研究中的多重比较P值均为校正值,P<0.05为显著,η2为效果量。
为进一步探讨在不同记忆负荷下,不同体力活动水平的个体行为表现和大脑活动的差异,将各记忆负荷下,各组的行为指标(正确率、反应时)和大脑活动指标(4个脑区的alpha频段能量、theta频段能量及theta频段能量/alpha频段能量)进行皮尔逊相关分析。
2 结果
2.1 行为结果分析
数字复述任务的正确率统计结果显示,2组正确率分别为92%和97%,根据先前的研究[25],认为数字任务的正确率在90%以上时,被试对数字任务进行了有效的加工。
生物运动工作记忆任務的行为结果如图2所示,正确率组别主效应显著(F(1,35)=8.563,P=0.006,η2=0.197),高体力活动组正确率显著高于低体力活动组(P=0.006),认知任务主效应显著(F(2,70)=18.916,P<0.001,η2=0.351),事后检验发现低等记忆负荷下的正确率显著高于中等记忆负荷(P=0.023)和高等记忆负荷(P<0.001),而中等记忆负荷正确率显著高于高等记忆负荷(P=0.003)。而认知任务与组别的交互作用不显著(F(2,70)=0.550,P=0.579,η2=0.15)。
反应时结果发现,认知任务主效应显著(F(2,70)=7.664,P=0.001,η2=0.180),事后检验发现高等记忆负荷下的反应时高于低等记忆负荷下的反应时(P=0.004),而中等记忆负荷下的反应时显著高于低等记忆负荷下的反应时(P=0.034)。但组别主效应(F(1,35)=3.233,P=0.081,η2=0.085)及任务与组别交互作用(F(2,70)=0.376,P=0.688,η2=0.11。)均不显著。
2.2 EEG结果分析
2.2.1 不同脑区的alpha频段能量分析
alpha频段能量的结果表明(如图3A和3B所示),组别主效应显著(F(1,35)=6.744,P=0.014,η2=0.162),事后检验发现高体力活动组被试的alpha频段能量显著高于低体力活动组(P=0.014)。脑区主效应显著(F (3,33)=9.225,P<0.001,η2=0.456),任务与脑区交互作用显著(F (6,30)=3.411,P=0.011,η2=0.406)。任务与脑区alpha频段能量的交互作用显示,通过简单效应分析发现,不同认知负荷对alpha频段能量的诱发差异主要发生在大脑额区,即高等工作记忆负荷显著大于低等工作记忆负荷诱发的alpha频段能量(P=0.018),大脑中央区高等工作记忆负荷诱发的alpha频段能量显著大于低等工作记忆负荷诱发的alpha频段能量(P=0.018)。经相关性检验发现,高体力活动组在中强度工作记忆负荷下,大脑额区的alpha频段能量与反应时呈正相关关系(r=0.507,P=0.032)。
另外,任务主效应(F (2,34)=0.841,P=0. 440,η2=0.047)、任务与组别的交互作用(F (2,34)=0.254,P=0.778,η2=0.015)、组别与脑区的交互作用(F (3,33)=1.230,P=0.314,η2=0. 101)及任务、脑区与组别的交互作用(F (6,30)=0.534,P=0.778,η2=0.096)均不显著。
2.2.2 不同脑区的theta频段能量比较
theta频段能量结果表明(如图3C和3D所示),组别主效应不显著(F (1,35)=1.118,P=0.298,η2=0.031),工作记忆负荷主效应不显著(F (2,34)=0.284,P=0.782,η2=0.014),组别与脑区的交互作用显著(F (3,33)=3.162,P=0.037,η2=0.223),工作记忆负荷与组别的交互作用(F (2,34)=0.542,P=0.587,η2=0.031)、工作记忆负荷与脑区交互作用(F (6,30)=1.723,P=0.150,η2=0.256)及工作记忆负荷、脑区与组别的交互作用(F (6,30)=1.426,P=0.238,η2=0.222)均不显著。脑区主效应显著(F (3,33)=97.834,P<0.001,η2=0.899),通过事后检验发现,大脑额区theta频段能量显著高于大脑中央区(P<0.001)、大脑顶区(P<0.001)和大脑枕区(P<0.001);大脑中央区theta频段能量显著高于大脑顶区(P<0.001)和大脑枕区(P<0.001);大脑顶区theta频段能量显著高于大脑枕区(P<0.001)。组别与脑区的交互作用显著(如前述),通过简单效应分析发现低体力活动组和高体力活动组均满足额区theta频段能量显著高于大脑中央区(P<0.001)、大脑顶区(P<0.001)和大脑枕区(P<0.001);大脑中央区theta频段能量显著高于大脑顶区(P<0.001)和大脑枕区(P<0.001),大脑顶区theta频段能量显著高于大脑枕区(P<0.001)。
2.2.3 不同脑区的theta频段能量/alpha频段能量分析
theta频段能量/alpha频段能量的结果表明,工作记忆负荷主效应显著(F (2,34)=3.303,P=0.049,η2=0.163)。通过事后检验发现,低等工作记忆负荷高于高等工作记忆负荷(P =0.043),脑区主效应显著(F(3,33)=97.978,P<0.001,η2=0.899)。通过事后检验发现,大脑额区显著高于大脑中央区(P<0.001)、大脑顶区(P<0.001)和大脑枕区(P<0.001);大脑中央区显著高于大脑顶区(P<0.001)和大脑枕区(P<0.001);大脑顶区显著高于大脑枕区(P<0.001)。工作记忆负荷与脑区交互作用显著(F (6,30)=6.120,P<0.001,η2=0.550),如图3E所示。通过简单效应分析发现,3个实验条件均满足大脑额区显著高于大脑中央区(P<0.001)、大脑顶区(P<0.001)和大脑枕区(P<0.001);大脑中央区显著高于大脑顶区(P<0.001)和大脑枕区(P<0.001);大脑顶区显著高于大脑枕区(P<0.001)。同时,在各个脑区,不同工作记忆负荷诱发的theta频段能量/alpha频段能量的不同仅出现在大脑额区,即低等工作记忆负荷高于中等工作记忆负荷(P=0.024)和高等工作记忆负荷(P=0.006)。组别与脑区的交互作用显著(F (3,33)=4.057,P=0. 015,η2=0.269),如图3F所示。通过简单效应分析发现,低体力活动组和高体力活动组均满足大脑额区显著高于大脑中央区(P<0.001)、大脑顶区(P<0.001)和大脑枕区(P<0.001);大脑中央区显著高于大脑顶区(P<0.001)和大脑枕区(P<0.001);大脑顶区显著高于大脑枕区(P<0.001)。同时,在大脑额区,高体力活动组theta频段能量/alpha频段能量显著低于低体力活动组(P=0.026)。
此外,组别主效应(F(1,35)=2.952,P=0.095,η2=0.078)、工作记忆负荷与组别的交互作用(F(2,34)=0.095,P=0.909,η2=0.006)及工作记忆负荷、脑区与组别的交互作用(F (6,30)=1.219,P=0.324,η2=0.196)均不显著。
3 討论
本研究通过对不同工作记忆负荷下,2组大学生认知行为绩效和大脑活动特征的比较发现,在行为层面,高体力活动组的生物运动工作记忆正确率优于低体力活动组;同时,随着工作记忆负荷的增大,被试的行为表现会下降。在电生理层面,组别优势主要表现在alpha频段,高体力活动组比低体力活动组诱发更多的alpha频段活动,工作记忆负荷与脑区存在交互作用,并且在额区和中央区均出现了高等记忆负荷下alpha频段能量大于低等记忆负荷下的结果,特别是高体力活动组在中等记忆负荷下,其额区所诱发的alpha频段能量与反应时间成正相关;theta频段能量和theta频段能量/alpha频段能量均出现了组别与脑区的交互作用,且2组被试均满足大脑额区>大脑中央区>大脑顶区>大脑枕区这一规律,而工作记忆负荷效应则发生在theta频段能量/alpha频段能量这一指标上,低等记忆负荷下该指标最高,而高等记忆负荷下,该指标最低并且两者具有显著差异。上述结果说明了不同体力活动水平个体在生物运动工作记忆时所诱发的大脑活动和加工策略存在差异。
3.1 不同体力活动水平大学生生物运动工作记忆加工的行为特征
高体力活动组被试的数字复述正确率高于低体力活动组被试。由此推测,较高水平的体力活动可以对大学生生物运动工作记忆产生积极影响,这与已有研究结果一致,例如:有研究发现,高水平的体力活动对儿童的认知水平和学业成绩都有显著的促进作用[26];在老年人群中,体力活动水平越高,工作记忆的表现越好[27]。同时,本研究结果也显示,生物运动工作记忆负荷对个体工作记忆的表现具有显著影响,且不受体力活动水平的调节。对于2组被试,其工作记忆的正确率随着工作记忆负荷的增加而下降,工作记忆的反应时随着工作记忆负荷增加而延长。
虽然已有大量研究证实了体力活动对视空间工作记忆的记忆负荷、加工速度有积极作用,但本研究发现,对于生物运动信息的工作记忆加工过程,体力活动的促进作用仅体现在正确率上,对加工速度及记忆负荷的影响并不明显。这可能与生物运动工作记忆的加工方式有关。与一般视空间信息不同,对生物运动信息的工作记忆编码包括了时间序列上的加工,是一种细节信息的整合,其编码和复核比单一的视空间信息需要更多的认知资源,而这一过程,受知觉者加工信息(如认知风格)和知觉对象的基本特征(如光点数量、持续时间)影响。在本研究中,随着工作记忆负荷的增大,被试进行信息加工的持续时间也在增多,这可能是导致体力活动促进作用仅体现在加工正确率上的原因。
3.2 不同体力活动水平大学生生物运动工作记忆加工的大脑认知负荷特征
本研究通过记录大脑活动情况,比较和分析不同体力活动水平大学生在加工不同负荷工作记忆时大脑的激活特征。从结果来看,高体力活动组和低体力活动组被试的alpha频段能量存在显著的组间差异。高体力活动组被试的alpha频段能量值显著高于低体力活动组,并且在大脑额区、大脑中央区、大脑顶区和大脑枕区4个脑区均得到体现。已有研究发现,在认知过程中alpha频段能量越高,大脑的唤醒水平越低[15],认知加工的效率越高。本研究认为,被试在生物运动工作记忆加工的过程中,高体力活动组被试的alpha频段能量较高,说明其大脑的唤醒水平也较高,因此,高体力活动组被试在进行生物运动工作记忆加工时的神经效率更高。另外,记忆负荷对认知产生影响主要发生在额区位置。额区在工作记忆的过程中发挥着重要的作用。已有研究认为,额区损伤者执行空间工作记忆[28]和视空间工作记忆[29]任务的行为结果显著低于正常人。本研究的结果显示,在大脑额区高体力活动组被试完成生物运动工作记忆任务的水平显著高于低体力活动组,因此,高体力活动组被试在进行生物运动工作记忆时的大脑唤醒效率更高,而大脑额区在信息加工过程中的唤醒程度受工作记忆负荷的调节最为明显。
从theta频段能量结果来看,组别与脑区存在显著的交互作用,且高低体力活动组被试4个脑区的激活水平均满足大脑额区>大脑中央区>大脑顶区>大脑枕区的规律,但在theta频段并没有发现2组之间的体力活动水平的差异。因此,本研究中不同体力活动水平和不同工作记忆负荷下,大学生在进行生物运动信息工作记忆加工时调用的认知资源并未表现出差别。在分析theta频段能量/alpha频段能量指标时,发现了工作记忆负荷的调节,低等记忆负荷的theta频段能量/alpha频段能量指标值显著高于高等记忆负荷下该指标值。在已有的研究中,把theta频段能量/alpha频段能量作为评价认知负荷的敏感指标,这一指标将工作记忆与认知资源的调用和个体的大脑皮层唤醒水平进行了标准化,比单一的theta频段能量或alpha频段能量对工作记忆负荷的评价更加精准[30-31]。但已有研究显示,theta频段能量/alpha频段能量指标值会随着认知任务难度的提升而增大,但在本研究中,工作记忆负荷增加时,theta频段能量/alpha频段能量指标值反而会下降。这可能与本研究中知觉对象呈现的时间长短有关,虽然工作记忆负荷增大,并且不一定要提高认知任务难度,持续时间也会随之增多。
3.3 不足与展望
虽然本研究试图研究体力活动与生物运动工作记忆之间的规律,并得到了一些初步的结论,但仍存在局限。本研究中虽然以调节工作记忆负荷作为认知任务难度的变量,对认知任务难度的其他方面没有进行控制,这可能影响了研究结果。进一步的研究可调整光点数量或持续时间,调节认知任务的难度,通过增加认知资源的调用负荷探讨体力活动对生物运动的工作记忆加工策略的影响。
4 结论
高水平体力活动的大学生在认知行为表现上具有对生物运动工作记忆加工优势,表现出更高的再认正确率,同时,这种工作记忆加工优势在大脑活动的频率特征上主要表现在alpha频段的变化,高体力活动水平大学生的alpha频段激活效果显著优于低体力活动水平的大学生。说明其在对生物运动工作记忆加工的过程中,有着更高的神经加工效率和大脑唤醒水平。
参考文献:
[1] THOMPSON J, PARASURAMAN R. Attention, biological motion, and action recognition[J]. Neuroimage, 2012, 59(1): 4.
[2] WOOD J N. A core knowledge architecture of visual working memory: correction to wood(2011)[J]. J Exp Psychol Hum Percept Perform, 2011, 37(2): 357.
[3] PADILLA C, PéREZ L, ANDRéS P. Chronic exercise keeps working memory and inhibitory capacities fit[J]. Frontiers in Behavioral Neuroscience, 2014, 8(8): 49.
[4] CONN V S, HAFDAHL A R, MEHR D R. Interventions to increase physical activity among healthy adults: meta-analysis of outcomes[J]. American Journal of Public Health, 2011, 101(4): 751.
[5] BADDELEY A. Working memory[J]. Psychology of Learning & Motivation, 1992, 255(5044): 556.
[6] 郭瑋.不同运动锻炼方式老年人的视空间工作记忆的特征[D].上海:上海体育学院, 2017.
[7] 徐畅,周成林,马阳.生活方式对延缓老年人认知功能衰退的研究[J].体育科学,2014, 34(5):35.
[8] 文椈.学龄前儿童睡眠及体力活动对认知能力的交互作用[D].上海:上海体育学院,2017.
[9] DING X, ZHAO Y, WU F, et al. Binding biological motion and visual features in working memory[J]. J Exp Psychol Hum Percept Perform, 2015, 41(3): 850.
[10] PAVLOVA M A. Biological motion processing as a hallmark of social cognition[J]. Cerebral Cortex, 2012, 22(5): 981.
[11] HIRIS E, KREBECK A, EDMONDS J, et al. What learning to see arbitrary motion tells us about biological motion perception[J]. Journal of Experimental Psychology Human Perception & Performan, 2005, 31(5): 1096.
[12] SAUSENG P, GRIESMAYR B, FREUNBERGER R, et al. Control mechanisms in working memory: a possible function of eeg theta oscillations[J]. Neurosci Biobehav Rev, 2010, 34(7): 1015.
[13] NASSEF A. On-line monitoring and adaptive control of psychophysiological markers relating to humans under stress[D]. Sheffield:University of Sheffield, 2009.
[14] GEVINS A, SMITH M E, LE J, et al. High resolution evoked potential imaging of the cortical dynamics of human working memory[J]. Electroencephalography & clinical neurophysiology, 1996, 98(4): 327.
[15] NEYMOTIN S A, JACOBS K M, FENTON A A, et al. Synaptic information transfer in computer models of neocortical columns[J]. Journal of Computational Neuroscience, 2011, 30(1): 69.
[16] GOLDMAN R I, STERN J M, JR E J, et al. Simultaneous eeg and fmri of the alpha rhythm[J]. Neuroreport, 2002, 13(18): 2487.
[17] SHAGASS C. Electrophysiological studies of psychiatric problems[J]. Rev Can Biol, 1972(31): 77.
[18] KLIMESCH W. Eeg alpha and theta oscillations reflect cognitive and memory performance: a review and analysis[J]. Brain Res Brain Res Rev, 1999, 29(2-3): 169.
[19] HOCKEY G R, NICKEL P, ROBERTS A C, et al. Sensitivity of candidate markers of psychophysiological strain to cyclical changes in manual control load during simulated process control[J].Applied Ergonomics, 2009, 40(6): 1011.
[20] KYLE J, LO L, RODOLPHE G, et al. Changes in mental workload and motor performance throughout multiple practice sessions under various levels of task difficulty[J]. Journal of Neuroscience, 2018, 19(17): 2.
[21] 王正倫,孙飙,戴剑松.大学生体质与体力活动的相关分析和研究[J].武汉体育学院学报,2006,40(12):67.
[22] JOHANSSON G. Visual perception of biological motion and a model for its analysis[J]. Perception & Psychophysics, 1973, 14(2): 201.
[23] VANRIE J, VERFAILLIE K. Perception of biological motion: a stimulus set of human point-light actions[J]. Behavior Research Methods Instruments & Computers, 2004, 36(4): 625.
[24] CURBY K M, GAUTHIER I. A visual short-term memory advantage for faces[J]. Psychonomic Bulletin & Review, 2007, 74(4): 620.
[25] XIQIAN L, JIAN H, YUJI Y, et al. Holding biological motion in working memory: an fmri study[J].Cognitive Neuroscience Journal, 2016, 10(1): 198.
[26] DONNELLY J E, HILLMAN C H, CASTELLI D, et al. Physical activity, fitness, cognitive function, and academic achievement in children: a systematic review[J]. Med Sci Sports Exerc, 2016, 48(6): 1197.
[27] CHANG Y K, HUANG C J, CHEN K F, et al. Physical activity and working memory in healthy older adults: an erp study[J]. Psychophysiology, 2013, 50(11): 1174.
[28] PETRIDES M, MILNER B. Deficits on subject-ordered tasks after frontal- and temporal-lobe lesions in man[J]. Neuropsychologia, 1982, 20(3): 249.
[29] OWEN A M, DOWNES J J, SAHAKIAN B J, et al. Planning and spatial working memory following frontal lobe lesions in man[J]. Neuropsychologia, 1990, 28(10): 1021.
[30] HOLM A, LUKANDER K, KORPELA J, et al. Estimating brain load from the EEG[J]. Scientific World Journal, 2009(9): 639.
[31] GENTILI R J, RIETSCHEL J C, JAQUESS K J, et al. Brain biomarkers based assessment of cognitive workload in pilots under various task demands[C]// Chicago: Engineering in Medicine & Biology Society, 36th Annual International Conference of the IEEE, 2014.
