外来种克氏原螯虾(Procambarus clarkii)对3种沉水植物的牧食研究
2019-09-24袁野张尹哲吕超超马徐发夏成星
袁野, 张尹哲, 吕超超,2,3, 马徐发,2,3, 夏成星,2,3,*
外来种克氏原螯虾()对3种沉水植物的牧食研究
袁野1, 张尹哲1, 吕超超1,2,3, 马徐发1,2,3, 夏成星1,2,3,*
1. 华中农业大学水产学院, 农业部淡水生物繁育重点实验室, 武汉 430070 2. 淡水水产健康养殖湖北省协同创新中心, 武汉 430070 3. 池塘健康养殖湖北省工程实验室, 武汉 430070
为探讨外来种克氏原螯虾()对沉水植物牧食的偏好性和牧食强度, 采用投喂实验的方式, 研究了3种沉水植物刺苦草()、轮叶黑藻()和伊乐藻()的适口性。结果表明, 克氏原螯虾对轮叶黑藻的取食速率最大(7.24±0.24 mg·d-1), 其次是苦草(3.70±1.14 mg·d-1), 伊乐藻最小(0.60±0.12 mg·d-1), 说明伊乐藻的适口性最差, 轮叶黑藻的适口性最好, 苦草的适口性居中。3种沉水植物的纤维素、多酚含量和氮含量都没有显著性差异, 而伊乐藻却具有更高的碳(C)含量和更高的碳氮比(C:N)。总体来说, 3种沉水植物的物理结构、碳(C)含量和碳氮比(C:N)在外来种克氏原螯虾对沉水植物的牧食中具有重要的作用。
克氏原螯虾; 沉水植物; 牧食; 适口性; 碳氮比
0 前言
克氏原螯虾()俗称小龙虾, 隶属于节肢动物门(Arthropoda)、甲壳纲(Crustacea)、十足目(Decapoda)、螯虾科(Cambaridae), 原产于墨西哥东北部和美国南部地区, 是世界分布最广泛的淡水螯虾入侵种类[1–3], 现已广泛分布于我国中东部地区的各种淡水水体, 尤其是长江中下游地区[4–9]。克氏原螯虾具有较强的生态可塑性, 其生态幅宽、适应性广, 具有食性杂、生长快、成熟早、繁殖力高等生物学特性, 使得克氏原螯虾可以在入侵水体迅速建立种群[10–12]。克氏原螯虾作为淡水水体重要的消费者, 对水体初级生产者和次级消费者的生物多样性和群落结构具有较大的影响[13–15], 其可以通过取食或者挖掘等行为, 使大型水生植物大量减少, 从而可使水体由周丛藻类和沉水植物为主的清水状态转变为以浮游植物为主的浑浊状态[16–17]。
植物适口性(Plant palatability)是植物一个重要的多功能性状的综合生态特征, 决定了牧食者的牧食选择和植物可能承受的牧食压力[18], 主要包括植物组织的物理性状(Physical characteristics)和化学性状(Chemical characteristics), 物理性状主要包括植物体的外层结构和一些特殊的防御结构; 化学性状主要是植物体内的营养物质含量、次生代谢产物(酚、生物碱、类三萜和皂角苷等)的含量[19–21]。植物中营养物质含量(C、N)和碳氮比(C:N)也是评价植物适口性的重要依据[22–23], 植物体具高氮(N)、低碳(C)和低碳氮比(C:N), 其适口性相对较高[24]。Bakker等[25]研究沉水植物化学计量特征对适口性影响时, 发现氮(N)含量高的植物具有更好的适口性, 且水生植物组织含氮(N)量和水生动物对其取食消耗成正相关。除了元素特征差异决定了植物的适口性以外, 纤维素和多酚含量与植物的适口性也密切相关。纤维素是植物细胞壁的主要组成成分, 纤维素含量增加, 可以使植物叶片的韧性增加, 降低植物的适口性[26]。朱顺清等[27]在草食性鱼类对水草的牧食研究中, 发现纤维素含量较低的苦草、轮叶黑藻和菹草比纤维素含量高的黄丝草具有更高的适口性, 更易于鱼类的消化吸收, 草食性鱼类也更为喜食。多酚作为广泛存在于植物各器官中的一种复杂酚类的次生代谢产物, 含量仅次于纤维素、半纤维素和木质素, 可作为化学防御物质来抵御牧食者的牧食[22,28]。入侵欧洲和美洲地区的穗状狐尾藻体内具有较多的酚类物质, 其高浓度的酚类不仅可以降低本身的适口性抵御牧食者牧食, 还可以使鳞翅目的幼虫生长速度降低[29–30]。
克氏原螯虾主要通过取食和挖掘等行为来影响水生植物的生长和群落演替, 并且受到水深和捕食者的影响[17,31–33]。Carreira等[31]在克氏原螯虾对5种水生植物(,,,,)的牧食实验中, 发现克氏原螯虾更喜食, 而不喜食和, 不同种类植物的取食消耗和断枝数量主要取决于克氏原螯虾的大小而并非性别, 但对造成取食速率差异和植物适口性差异的机制并未进行深入探讨。国内多数研究主要集中于克氏原螯虾的人工养殖技术、食物组成、摄食节律和肌肉品质[34–36], 对克氏原螯虾的入侵生态学研究也相对较少[7–10,37], 对克氏原螯虾对沉水植物的牧食选择性研究更少[38]。
本研究主要通过适口性实验, 比较分析了入侵物种克氏原螯虾对3种水鳖科沉水植物的取食速率以及沉水植物C、N元素和防御物质对牧食的影响, 探讨了克氏原螯虾对3种沉水植物的牧食选择性差异及其原因, 可以为牧食理论的研究、沉水植被恢复和沉水植物过度生长控制提供数据支持和理论依据, 为克氏原螯虾的防控提供一定的参考。
1 材料与方法
1.1 实验材料
本研究所用沉水植物3种, 隶属于水鳖科(Hydrocharitaceae), 分别为刺苦草(V)、轮叶黑藻()和伊乐藻(), 其中伊乐藻为外来种。所用水生植物均采集于武汉市青菱湖, 采集清洗干净后, 培养于基质为沙子的水族缸中。实验开始时, 取刺苦草的叶片、轮叶黑藻和伊乐藻的顶枝(均为20 cm)用清水漂洗干净、备用。
实验用克氏原螯虾采集于青菱湖及周边池塘、沟渠, 采集后带回实验基地室内暂养, 为防止克氏原螯虾互相打斗受伤, 分别单独放入具盖塑料桶(防止逃逸)中, 并用沉水植物投喂, 定期换水, 更换植物。实验开始前, 挑取大小相对一致、生命力强的雌性克氏原螯虾进行实验(保持性别一致), 实验开始之前饥饿处理24 h。
1.2 实验设计
本实验在华中农业大学实验基地室内进行。实验时间为2016年9月11日-9月18日, 实验周期为1周。实验设置2个处理组(牧食组和对照组), 3个重复, 牧食组内放入1株植物和1只克氏原螯虾(体重: (28.53±2.92) g, Mean±SE), 各处理组克氏原螯虾大小没有显著差异(2,8= 0.075;= 0.929), 对照组内仅放入顶枝, 用于实验周期内沉水植物生长指标的计算。实验选取3种水生植物顶枝(苦草为叶片), 每株20 cm, 且每种留取大小相似的顶枝或叶片20个, 烘干、称重计算干湿比。实验容器为锥形塑料盒(上部直径×下部直径×高, 17 cm×13 cm×8.5 cm), 水深为6.5 cm。每个容器中放置带孔盒盖防止克氏原螯虾逃逸。
1.3 日取食量的测定
投喂实验开始前, 称量每组植物鲜重和克氏原螯虾体重, 1周实验结束后, 将克氏原螯虾取出后用捞网和镊子将取食后的植物碎屑和空白组内植物取出, 晾置数分钟后称取鲜重。
采用Carreira[31]的方法计算克氏原螯虾对沉水植物的日取食量, 即取食速率, 比较水生植物的适口性。
取食速率(Daily consumed biomass,B)按下式计算:
B= (W×–W)×W
式中,W为初始鲜重;为空白对照组顶枝的自生长率(实验结束时鲜重/初始鲜重);W为实验组结束时的鲜重;W为顶枝的干湿比。
1.4 植物化学成分的测定
用Folin-Ciocaltau方法[39]测定植物多酚含量, 取少量各处理组3种沉水植物叶子和茎烘干并磨成粉末, 然后取大约2.0—8.0 mg用来测定多酚。
称取4.0 mg植物粉末样品采用蒽酮比色法[40]测定纤维素含量。取大约4.0 mg的植物粉末, 利用CHNS/O元素分析仪(Elementar Vario PYRO cube, German), 采用杜马斯燃烧法进行碳、氮连续分析, 测定碳、氮元素含量, 每个植株样品测量3次, 每个处理测量3个重复。
1.5 数据分析
在数据分析前, 为满足数据的正态分布和方差的齐次性, 所有的数据都经过log转换。利用单因素方差分析(One-way ANOVA), 对克氏原螯虾取食速率和3种沉水植物多酚、纤维素、碳含量、氮含量和碳氮比(C/N)进行显著性分析, 利用Turkey's HSD的方法对不同处理间差异进行多重比较。数据分析采用SPSS 24.0统计分析软件进行。
2 结果与分析
2.1 克氏原螯虾对3种沉水植物的牧食
克氏原螯虾对3种沉水植物的取食速率具有显著性差异(<0.01)(图1a), 其取食速率大小为: 轮叶黑藻>苦草>伊乐藻, 实验周期内鲜重取食量分别为(2.39±0.57) g 、(1.89±0.27) g和(0.61±0.07) g(表1), 取食速率分别为(7.24±0.24) mg·d-1、(3.70±1.14) mg·d-1和(0.60±0.12) mg·d-1, 表明克氏原螯虾更喜食轮叶黑藻, 其适口性最好, 而伊乐藻的适口性最差。
2.2 3种沉水植物的纤维素和多酚含量
3种沉水植物的纤维素和多酚含量没有显著性差异(>0.05)(图1b,c), 轮叶黑藻、刺苦草、伊乐藻的纤维素含量分别为(128.32±12.49) mg·g-1、(126.02±29.58) mg·g-1和(149.30±9.63) mg·g-1, 多酚含量分别为(3.11±0.39) mg·g-1、(2.44±0.23) mg·g-1和(2.70±0.23) mg·g-1, 表明纤维素和多酚不是影响克氏原螯虾对3种沉水植物牧食的主要因素。

表1 克氏原螯虾牧食前后3种沉水植物的鲜重(Mean±SE)
2.3 3种沉水植物的碳氮含量
3种沉水植物之间的C含量和C:N比值之间具有显著性差异, N含量无显著性差异(图1d,e,f)。轮叶黑藻、刺苦草和伊乐藻的碳(C)含量分别为(359.00±1.49) mg·g-1、(333.21±3.35) mg·g-1和(379.28±3.84) mg·g-1; 氮(N)含量分别为(34.90±0.96) mg·g-1、(31.62±1.93) mg·g-1和(28.95±1.47) mg·g-1; C:N比值分别为(10.30±0.24)、(10.62±0.64)和(13.16± 0.59)。伊乐藻的C含量显著高于轮叶黑藻和刺苦草, 而伊乐藻的C:N比值显著高于轮叶黑藻和刺苦草, 但轮叶黑藻和刺苦草之间C:N比值却没有显著性差异。
3 讨论
3.1 克氏原螯虾对3种沉水植物的牧食
克氏原螯虾作为淡水生态系统中重要的杂食性消费者[1–2,11–12], 以底栖无脊椎动物、碎屑、水生植物和藻类为食, 并且可以通过挖掘和直接切割成植物碎片来影响水生植物的生长和群落演替[41–42]。Cronin等[23]研究发现, 克氏原螯虾更喜食丝状或细小分枝、易碎的水生植物, 而本研究中, 轮叶黑藻相对于另外两种沉水植物来说, 其具有细小分枝、易碎等特性, 克氏原螯虾对其的鲜重和干重的取食量都是最高的, 结果表明, 水生植物的物理结构对克氏原螯虾的牧食具有一定的影响。
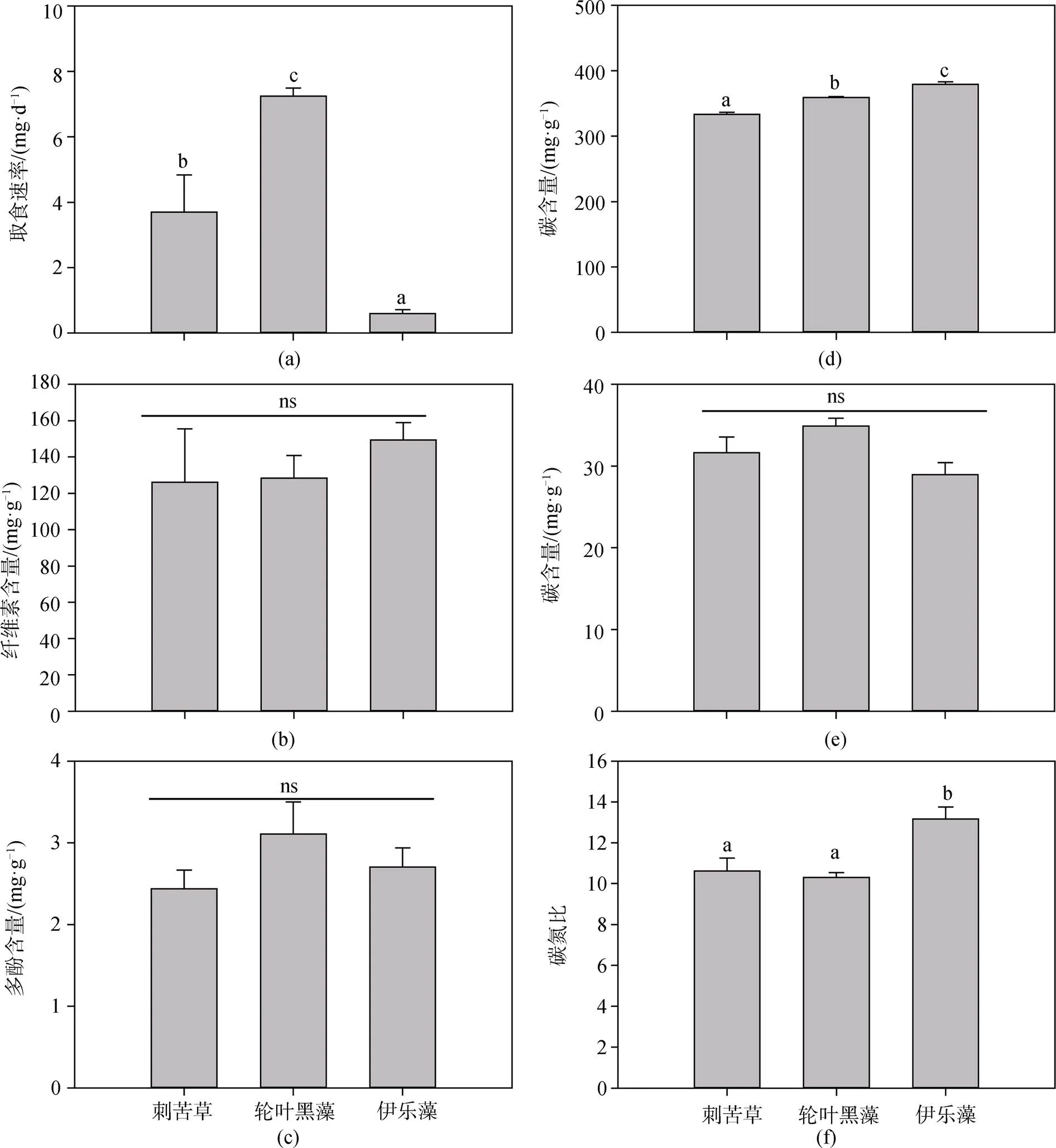
图1 克氏原螯虾对3种沉水植物的取食速率及3种沉水植物纤维素、多酚、碳、氮含量和碳氮比
Figure 1 The daily consumed biomass by herbivory of, and the cellulose content, polyphenol content, C content, N content and C:N ratios of three submerged plants
Carreira等[31]研究5种水生植物的适口性时发现, 克氏原螯虾每天可以消耗单位体重0.6%—2.4%的水生植物干物质量, 取食速率为42.6—114.9 mg·d-1; 而Cronin等[23]研究发现克氏原螯虾对水生植物的平均鲜重取食量大约为440 mg·d-1, 而本研究中克氏原螯虾对轮叶黑藻、苦草、伊乐藻的取食速率分别为(7.24±0.24) mg·d-1、(3.70±1.14) mg·d-1和(0.60±0.12) mg·d-1, 鲜重取食速率约为232.66 mg·d-1, 取食速率相对偏低, 可能与研究的物种个体差异有关, 在Carreira等[31]和Cronin等[23]的研究中都以挺水植物为主, 而挺水植物相对沉水植物来说含有更多的碳和更低的氮[43], 克氏原螯虾为了满足自身的生长代谢需求, 需要摄食更多的水生植物。
3.2 纤维素和多酚含量对3种沉水植物适口性的影响
植物适口性决定了草食性动物对植物的牧食选择性, 也决定了植物可能承受的牧食压力[28], 纤维素作为植物细胞壁的主要组成成分, 在一定程度上增加了植物叶片的物理防御能力, 降低了植物的适口性[21], 同一种植物在不同生长发育期或不同植物在同一生长发育期, 其纤维素含量也会有所差异[44]。但本研究中, 3种沉水植物的纤维素含量却没有显著性差异(图1b), 其中, 轮叶黑藻、刺苦草、伊乐藻的纤维素含量分别为(128.32±12.49) mg·g-1、(126.02±29.58) mg·g-1和(149.30±9.63) mg·g-1, 结果表明这3种沉水植物的适口性的差异可能与其它结构性物质或防御物质有关, 如化学防御物质[44]。
多酚作为植物体内重要的化学防御物质不仅可以降低植物的适口性, 在一定程度上反应沉水植物对环境的适应能力[28], 还可以影响牧食者的生长发育[29–30]。但也有研究表明, 酚类的含量不仅受到牧食的影响, 还受到植物生活型、光照、营养等环境因素的影响, 同时酚类含量的多少并不会影响植物的适口性[19,22,45–46]。而Bolser等[47]研究发现昆虫牧食过的植物, 对克氏原螯虾来说, 适口性更低, 但却不是由于多酚和蛋白质的差异造成的, 而可能是一些其它的次生代谢产物或营养物质的差异造成的[30]。本研究中, 3种沉水植物的多酚含量并没有显著差异(图1c), 这与Bolser等[47]人的研究结果一致, 说明这3种沉水植物的适口性差异很可能时由于植物的其它次生代谢产物或营养物质含量的差异造成的。
3.3 C、N和C:N对3种沉水植物适口性的影响
植物适口性与植物体内的营养物质含量密切相关, Bryant等[48]提出了植物防卫物质类型(“量”型和“质”型)不同很可能是由于植物体内C、N元素的不平衡所引起的, 在高养分或碳受限的条件下, 碳基防御物质会减少, 而氮基防御物质就会相应增加。Bakker等[25]关于3种不同生境(陆生、淡水、海洋)水生植物碳氮含量及其比值对草食性动物取食速率的研究表明, 水生植物氮(N)含量高、碳氮比(C:N)较低, 对牧食者来说适口性就更高, 反之则适口性更低。本研究中, 3种沉水植物的氮(N)含量没有显著性差异, 但伊乐藻的碳(C)含量和碳氮比(C:N)显著高于刺苦草和轮叶黑藻(图1d,f), 说明碳含量在克氏原螯虾的牧食选择性上具有重要的作用。植物体内的碳主要用于合成含碳化合物, 主要用于增加细胞壁的厚度或韧性, 增加牧食者的消化难度[49], 而本研究中, 富碳化合物—纤维素的含量没有显著性差异, 说明这3种沉水植物的适口性可能与其它富碳化合物有关, 如木质素等。
4 结论
(1)对水鳖科3种沉水植物刺苦草、轮叶黑藻和伊乐藻来说, 克氏原螯虾更喜食轮叶黑藻, 最不喜食伊乐藻, 伊乐藻的适口性最差。
(2)轮叶黑藻的适口性最好, 伊乐藻的适口性最低, 初步推断, 可能是由于水生植物的物理结构差异和富碳化合物含量较高造成的。但由于本实验周期较短, 影响牧食者的选择性是否还存在其他因素, 还需要进一步深入研究。
[1] Hobbs III H H, Jass J P, Huner J V. A review of global crayfish introductions with particular emphasis on two North American species (Decapoda, Cambaridae)[J]. Crustaceana, 1989, 56(3): 299–316.
[2] Loureiro T G, Anastácio P M S G, Araujo P B, et al. Red Swamp crayfish: biology, ecology and invasion-an overview[J]. Nauplius, 2015, 23(1): 1–19.
[3] Francesca G. Crayfish invading Europe: the case study of[J]. Marine and Freshwater Behaviour and Physiology, 2006, 39(3): 175–191.
[4] 王卫民. 软壳克氏原螯虾在我国开发利用的前景[J]. 水生生物学报, 1999, 23(4): 375–381.
[5] 李振宇, 解焱. 中国外来入侵种[M]. 北京: 中国林业出版社, 2002.
[6] 王亚民, 曹文宣. 中国水生外来入侵物种对策研究[J]. 农业环境科学学报, 2006, 25(1): 13–19.
[7] 曹玲亮, 周立志, 张保卫. 安徽三大水系入侵物种克氏原螯虾的种群遗传格局[J]. 生物多样性, 2010, 18(4): 398–407.
[8] 王长忠, 李忠, 梁宏伟, 等. 长江下游地区4个克氏原螯虾群体的遗传多样性分析[J]. 生物多样性, 2009, 17(5): 518–523.
[9] 张萌, 白俊, 金辉, 等. 不同地理群体的克氏原螯虾形态差异多元分析[J]. 南昌大学学报(理科版), 2016, 40(2): 188–196.
[10] 蔡凤金, 武正军, 何南, 等. 克氏原螯虾的入侵生态学研究进展[J]. 生态学杂志, 2010, 29(1): 124–132.
[11] Holdich D M, Lowery R S. Freshwater crayfish: biology, management, and exploitation[M]. London: Croom Helm, 1988.
[12] Twardochleb L A, Olden J D, Larson E R. A global meta-analysis of the ecological impacts of nonnative crayfish[J]. Freshwater Science, 2013, 32(4): 1367–1382.
[13] Correia A M, Anastácio P M. Shifts in aquatic macroinvertebrate biodiversity associated with the presence and size of an alien crayfish[J]. Ecological Research, 2008, 23(4): 729–734.
[14] Gamradt S C, Kats L B, Anzalone C B. Aggression by non-native crayfish deters breeding in California Newts[J]. Conservation Biology, 1997, 11(3): 793–796.
[15] Reynolds J D, Soutygrosset C, Kozak P, et al. A review of ecological interactions between crayfish and fish, indigenous and introduced[J]. Knowledge and Management of Aquatic Ecosystems, 2011, 401(10): 1–21.
[16] Matsuzaki S I S, Usio N, Takamura N, et al. Contrasting impacts of invasive engineers on freshwater ecosystems: an experiment and meta-analysis[J]. Oecologia, 2009, 158(4): 673–686.
[17] Van der Wal J M, Dorenbosch M, Immers A K, et al. Invasive crayfish threaten the development of submerged macrophytes in lake restoration[J]. PLoS ONE, 2013, 8(10): e78579.
[18] Fraser L H, Grime J P. Interacting effects of herbivory and fertility on a synthesized plant community[J]. Journal of Ecology, 1999, 87(3): 514–525.
[19] Cronin G. Influence of macrophyte structure, nutritive value, and chemistry on the feeding choices of a generalist crayfish[M]. New York: Springer, 1998.
[20] Mithöfer A, Boland W. Plant defense against herbivores: chemical aspects[J]. Annual Review of Plant Biology, 2012, 63(1): 431–450.
[21] Turley N E, Godfrey R M, Johnson M T. Evolution of mixed strategies of plant defense against herbivores[J]. New Phytologist, 2013, 197(2): 359–361.
[22] Lodge D M. Herbivory on freshwater macrophytes[J]. Aquatic Botany, 1991, 41(1/3): 195–224.
[23] Cronin G, Lodge D M, Hay M E, et al. Crayfish feeding preferences for freshwater macrophytes: the influence of plant structure and chemistry[J]. Journal of Crustacean Biology, 2002, 22(4): 708–718.
[24] Cebrian J, Shurin J B, Borer E T, et al. Producer nutritional quality controls ecosystem trophic structure[J]. PLoS ONE, 2009, 4(3): e4929.
[25] Bakker E S, Wood K A, Pagès J F, et al. Herbivory on freshwater and marine macrophytes: a review and perspective[J]. Aquatic Botany, 2016, 135: 18–36.
[26] Wardle D A, Bonner K I, Barker G M. Linkages between plant litter decomposition, litter quality, and vegetation responses to herbivores[J]. Function Ecology, 2010, 16(5): 585–595.
[27] 朱清顺, 周刚, 于能, 等. 以滆湖为例湖泊种植饲料水草的养鱼效果研究[J]. 水产学报, 1993, 17(3): 189–197.
[28] Smolders A J P, Vergeer L H T, Velde G V D, et al. Phenolic contents of submerged, emergent and floating leaves of aquatic and semi-aquatic macrophyte species: why do they differ[J]? Oikos, 2000, 91(2): 307–310.
[29] Choi C, Bareiss C, Walenciak O, Gross E M. Impact of polyphenols on growth of the aquatic herbivore[J]. Journal of Chemical Ecology, 2002, 28(11): 2245–2256.
[30] Gross E M, Meyer H, Schilling G. Release and ecological impact of algicidal hydrolysable polyphenols in[J]. Phytochemistry, 1996, 41(1): 133–138.
[31] Carreira B M, Dias M P, Rebelo R. How consumption and fragmentation of macrophytes by the invasive crayfishshape the macrophyte communities of temporary ponds[J]. Hydrobiologia, 2014, 721(1): 89–98.
[32] Gherardi F, Aquiloni L, Diéguezuribeondo J, et al. Managing invasive crayfish: is there a hope[J]? Aquatic Sciences, 2011, 73(2): 185–200.
[33] Aquiloni L, Gherardi F. The use of sex pheromones for the control of invasive populations of the crayfish: a field study[J]. Hydrobiologia, 2010, 649(1): 249–254.
[34] 肖鸣鹤, 肖英平, 吴志强, 等. 养殖密度对克氏原螯虾幼虾生长、消化酶活力和生理生化指标的影响[J]. 水产学报, 2012, 36(7): 1088–1093.
[35] 刘琦. 克氏原螯虾对两种环境异质性的响应及对隐蔽所的选择[D]. 南京: 南京大学, 2017.
[36] 田娟, 许巧情, 田罗, 等. 洞庭湖克氏原螯虾肌肉成分分析及品质特性分析[J]. 水生生物学报, 2017, 41(4): 870–877.
[37] 武正军, 蔡凤金, 贾运锋, 等. 桂林地区克氏原螯虾对泽蛙蝌蚪的捕食[J]. 生物多样性, 2008, 16(2): 150–155.
[38] 徐增洪, 周鑫, 水燕. 克氏原螯虾的食物选择性及其摄食节律[J]. 大连海洋大学学报, 2012, 27(2): 166–170.
[39] Ragan M A, Craigie J S. Phenolic compounds in brown and red algae[M]. London: Cambrige University Press, 1978.
[40] Goering H K, Van Soest P J. Forage fiber analysis: apparatus reagents, procedures and some applications. Washington D.C.: USDA, 1970.
[41] Nyström P, Brönmark C, Granéli W. Patterns in benthic food webs: a role for omnivorous crayfish[J]? Freshwater Biology, 1996, 36: 631–646.
[42] Nyström P, Strand J. A. Grazing by a native and an exotic crayfish on aquatic macrophytes[J]. Freshwater Biology, 2010, 36(3): 673–682.
[43] Xia Chengxing, Yu Dan, Wang Zhong, et al. Stoichiometry patterns of leaf carbon, nitrogen and phosphorous in aquatic macrophytes in eastern China[J]. Ecological Engineering, 2014, 70(5): 406–413.
[44] 郭新红, 喻达时, 王婕, 等.6种植物中木质纤维素含量的比较研究[J].湖南大学学报(自然科学版), 2008, 35(9): 76–78.
[45] 程春龙, 李俊清. 植物多酚的定量分析方法和生态作用研究进展[J]. 应用生态学报, 2006, 17(12): 2457–2460.
[46] 袁昌波, 曹特, 周存宇, 等. 洱海三种生活型水生植物叶片中碳、氮与总酚含量的比较研究[J]. 水生生物学报, 2016, 40(5): 1025–1032.
[47] Bolser R C, Hay M E, Lindquist N, et al. Chemical defenses of freshwater macrophytes against crayfish herbivory[J]. Journal of Chemical Ecology, 1998, 24(10): 1639–1658.
[48] Bryant J P, Chapin F S, Klein D R. Carbon/nutrient balance of boreal plants in relation to vertebrate herbivory[J]. Oikos, 1983, 40(3): 357.
[49] Gruner D S, Smith J E, Seabloom E W,et al. A cross-system synthesis of consumer and nutrient resource control on producer biomass[J]. Ecology letters, 2008, 11(7): 740–755.
Herbivory of the invasive crayfishon three submerged macrophytes
YUAN Ye1, ZHANG Yinzhe1, LV Chaochao1, MA Xufa1,2,3, XIA Chengxing1,2,3,*
1. Key Lab of Freshwater Animal Breeding, Ministry of Agriculture, College of Fisheries, Huazhong Agricultural University, Wuhan 430070, China 2. Freshwater Aquaculture Collaborative Innovation Center of Hubei Province, Wuhan 430070, China 3. Hubei Provincial Engineering Laboratory for Pond Aquaculture, Wuhan 430070, China
This study investigated the palatability of three species (,and) by feeding testto figure out the feeding preferences and the grazing intensity of submerged macrophytes for invasive crayfish. Our results showed that the daily consumed biomass of crayfish onwas the largest (7.24±0.24) mg·d-1, followed by(3.70±1.14) mg·d-1, and(0.60±0.12) mg·d-1, indicating that the palatability ofwas the worst,was the best, andwas in the middle. Our findings suggested that the cellulose content, polyphenol content, and nitrogen content (N) of the three submerged macrophytes were not significant difference, buthad more C and higher C:N ratio. In a word, the content of physical characteristics, C and C:N ratio of the three submerged macropyhtes played an important role on the herbivory of crayfish on submerged macrophytes.
; submerged macrophytes; herbivory; palatability; C:N ratio
10.14108/j.cnki.1008-8873.2019.05.004
Q948.8
A
1008-8873(2019)05-023-07
2018-10-19;
2018-11-14
国家自然科学基金青年科学基金项目(31500293); 中央高校基本科研业务费专项(2662015BQ038)
袁野(1990—), 男, 湖北十堰人, 硕士, 主要从事水生植物生理生态学研究, E-mail: 200831315@qq.com
夏成星, 男, 博士, 讲师, 主要从事淡水生态学研究, E-mail: xiachengxing@mail.hzau.edu.cn
袁野, 张尹哲, 吕超超, 等. 外来种克氏原螯虾()对3种沉水植物的牧食研究[J]. 生态科学, 2019, 38(5): 23-29.
YUAN Ye, ZHANG Yinzhe, LV Chaochao, et al. Herbivory of the invasive crayfishon three submerged macrophytes[J]. Ecological Science, 2019, 38(5): 23-29.
