遮阴对高山杜鹃叶片解剖和光合特性的影响
2019-09-10宋杰李树发李世峰蔡艳飞
宋杰 李树发 李世峰 蔡艳飞
摘 要:為了解高山杜鹃对光能的需求和适应性,该研究以盆栽3 a生高山杜鹃品种cv. Furnivalls Daughter为材料,探讨了遮阴对高山杜鹃叶片解剖结构和光合特性的影响。结果表明:光照强度对高山杜鹃品种cv. Furnivalls Daughter叶片的气孔密度没有显著影响,其气孔密度范围在299.70~327.22个·mm-2之间,但光照对气孔开度和单个气孔器的面积影响显著,100%全光照和30%全光照处理植株分别具有最小和最大的叶片气孔开度。在处理的光强范围内,随着光强减弱,叶片厚度、栅栏组织厚度、海绵组织厚度以及上、下表皮厚度逐渐降低,有利于提高叶片的光能利用效率。100%全光照处理下,高山杜鹃叶片的光饱和点(LSP)、净光合速率(Pn)、饱和光合速率(Pmax)、气孔导度(Gs)、蒸腾速率(Tr)均较低,遮阴处理有效提高了Pn、Pmax、Gs、Tr和光能利用效率(LUE),且30%全光照处理植株的叶片光补偿点(LCP)、暗呼吸速率(Rd)最低,而LSP、Pn、Pmax、Gs、Tr和LUE最高。这表明高山杜鹃在云南昆明地区的最适光照条件是30%左右的全光照,在高山杜鹃的栽培及应用中,应采取适当的遮阴措施以满足其生长的最佳光照条件。
关键词:高山杜鹃, 解剖结构, 光合, 光照
中图分类号:Q945.11
文献标识码:A
文章编号:1000-3142(2019)06-0802-10
Abstract:Rhododendron is a world famous ornamental garden plant and has significant economic and ornamental value. Light has an important impact on the growth and development of Rhododendron. However, the studies on the demand and adaptability of Rhododendron are scarce. In order to understand the demand and adaptability of Rhododendron to light, 3-year-old plants of Rhododendron cv. Furnivalls Daughter were used as materials and the effects of shading on leaf anatomical structure and photosynthesis were studied. The results were as follows:The impact of light intensity on the stomata density of Rhododendron cv. Furnivalls Daughter was not obvious, and the stomatal density ranged from 299.70 to 327.22 mm-2. However, the stomatal size and the area of stomata apparatus were significantly influenced by light intensity. The plants of 100% and 30% full light intensities had the minimum and maximum stomatal size, respectively. As the light intensity decreased, the thickness of leaf, palisade tissue, sponge tissue, as well as the thickness of adaxial epidermis and abaxial epidermis decreased so as to improve the light use efficiency of leaves. The plasticity analysis showed that the plasticity index of mesophyll-related parameters such as leaf thickness, abaxial epidermis thickness, and the thickness of palisade and sponge tissue were higher, while that of stomatal-related parameters such as stomatal density, stomatal length and width were lower, which indicates that mesophyll tissue plays a more important role in the process of adaption to different light environments. The analysis of light response curves and photosynthetic parameters showed that the plants of 100% full light intensity were inhibited and damaged by strong light, and the plants showed the lowest light saturation point (LSP), net photosynthetic rate (Pn), photosynthetic rate at light saturation point (Pmax), stomatal conductance (Gs) and transpiration rate (Tr). After shading treatments, Pn, Pmax, Gs, Tr and light use efficiency (LUE) were improved. Compared with other shading treatments, the plants of 30% full light intensity not only had the lowest light compensation point (LCP), dark respiration rate (Rd), but also had the highest LSP, Pn, Pmax, Gs, Tr and LUE. The above results suggest that the optimum light intensity of Rhododendron cv. Furnivalls Daughter in Kunming is about 30% full light intensity. In the cultivation and application of Rhododendron, we should take some shading measures to meet the optimum light conditions for its growth.
Key words:Rhododendron, anatomical structure, photosynthesis, light
高山杜鹃是杜鹃花中的一大类,一般指无鳞杜鹃花亚属、有鳞杜鹃花亚属、马银花亚属中的常绿杜鹃,以及它们的杂交品种(李倩等,2009)。高山杜鹃色彩鲜艳,花姿优美,株型漂亮,具有极高的经济价值和观赏价值(李志斌等,2007)。然而,在高山杜鹃的栽培及园林应用过程中发现,光照对高山杜鹃的生长发育具有重要影响。强光下高山杜鹃的生长缓慢,叶片发黄,并伴有日灼现象发生,而过度遮阴则会导致高山杜鹃的枝条徒长,株型散乱,花芽数量减少,花色变淡,大大降低高山杜鹃的观赏性和经济价值(郑宝强等,2010)。
光照是影响植物形态和功能的重要因子,对植物的生长、发育和演化具有极其重要的作用(Aleric & Kirkman,2005;Sofo et al., 2009;裘珍飞等,2017;赵斌等,2017)。植物在长期适应不同光环境的过程中,其叶片形态和生理特性会发生相应改变,以最大程度维持光合能力,从而适应不同的光环境(Evans,1989;Cregg et al., 1993;Niinemets & Tenhunen,1997;Pandey et al., 2009)。植物对环境的适应能力直接或间接地与光合能力相关,反过来环境因子对植物叶片的结构和功能产生影响,进而影响整个植株的生长(Chandra,2003;吴正花等,2018)。近年来,关于杜鹃属植物形态结构与环境适应关系已引起科研人员的研究興趣。Wang et al.(2008)研究了2种杜鹃R. catawbiense和R. ponticum过冬叶的结构,阐述强光照和干旱胁迫对叶片结构的影响,以及植物自身对环境的适应性。王艳萍等(2012)研究了3种长白山高山杜鹃叶片解剖结构与高山环境的适应性;容丽等(2009)分析了杜鹃属13种植物叶片解剖结构的生态适应性;石登红和陈训(2005)研究了贵州省杜鹃花属植物叶片解剖结构的生态适应性;曹晓娟等(2009)对太白山5种杜鹃属植物的叶片解剖结构生态适应性的研究表明,5种杜鹃的栅海组织比均远小于1,属耐荫植物。其中,强光适应性较好、耐荫性较差的杜鹃栅栏组织发达、排列紧密,而强光适应性较差、耐荫性较强的杜鹃叶片气孔密度最高、海绵组织排列疏松。从上述研究可以发现,目前关于高山杜鹃对光强的需求和适应性研究较少,且主要集中在一些高山杜鹃集中分布的区域和一些野生高山杜鹃种质资源,而被广泛应用于生产和园林绿化中的园艺品种的叶片结构和光合特性对光强的适应性研究较少。
本研究中以市场流行的主要种植的高山杜鹃品种cv. Furnivalls Daughter为材料,研究遮阴对其叶片解剖结构和光合特性的影响,了解高山杜鹃对光能的需求和适应性,寻找其最佳的生长光强,为高山杜鹃的规模化栽培和园林应用提供理论基础和实践指导。
1 材料与方法
1.1 材料
研究材料是3年生高山杜鹃品种cv. Furnivalls Daughter,种植于云南省农业科学院花卉研究所大春河基地,进行常规水肥管理。试验地属低纬高原山地季风气候,干湿季分明,5—10月为雨季,11月至次年4月为旱季。8月份开始用黑色遮阳网进行遮阴处理,遮阴程度采用不同层数的遮阳网叠加获得,遮阴处理分别设1层、2层和3层遮阳网。采用DRM-FQ双辐射测量其光强,分别为全光照的53%、30%和17%(3种遮阴处理分别记为L1、L2、L3),全光照为对照(L0表示)。试验过程中所有处理植株均种植在同一个试验大棚内,除光照具有显著差异外,不同处理间的温度、湿度变化没有显著差异。每个处理10盆,3次重复,随机区组排列,定期移动花盆以消除边缘影响。次年3月份测定各指标。
1.2 方法
1.2.1 叶片解剖结构观察 取成熟叶片中间部分(避开叶脉),用FAA固定液固定1周,用梯度酒精脱水,二甲苯透明,浸蜡、包埋。包埋后的材料在切片机(MICROM HM315, Germany)上进行叶片横切面的组织切片,切片厚度8~10 μm,苏木精染色制成永久切片。表皮结构观察采用50%次氯酸钠溶液浸泡1周后用刀片刮去叶肉,将表皮制成临时装片,在生物显微镜(Nikon Eclipse E800, Yokohama, Japan)下观察并用Nikon-DXM1200数码显微摄影系统拍照。用图形分析软件(Image-Pro Plus 6.0)测量叶片厚度、角质层厚度、上下表皮厚度、栅栏组织及海绵组织厚度,气孔长、气孔宽、气孔密度、气孔器长、气孔器宽。气孔密度(SD)=视野内气孔个数/视野面积。单个气孔器的面积(As)=π×气孔器长度(l)×气孔器宽度(w)/4,气孔器面积百分率(At)=As×SD×100(James & Bell,2001)。每个处理取30个视野中的平均值。
1.2.2 叶片光合参数测定 叶片的光合作用采用便携式光合作用测定系统LI-6400 XT (Li-Cor Inc. Nebraska, USA)进行测定。以LI-6400-02 LED提供红蓝光源来获得不同的光强梯度(2 000、1 600、1 200、1 000、800、600、500、400、300、200、150、100、50、20和0 μmol·m-2·s-1),CO2采用开放气流通路。测定前用1 000 μmol·m-2·s-1的光强诱导15 min。测定时,叶室温度设为20 ℃,相对湿度50%~60%。在每一光强平衡3 min左右,待ΔH2O、ΔCO2、净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)等读数稳定时记录。每个处理至少重复测定3片叶子。
用光合助手软件(Photosyn Assistant, V1. 1, Dundee Scientific, UK)拟合光响应曲线(Rothstein & Zak,2001),该软件应用的模型方程是非直角双曲线方程(Prioul & Chartier,1977)。根据此方程计算饱和光合速率(Pmax)、光饱和点(LSP)、光补偿点(LCP)、暗呼吸速率(Rd)等光合参数。
1.2.3 数据分析 所有数据采用统计软件SPSS 13.0 for Windows软件包(SPSS Inc., Chicago, USA)进行分析,处理间比较采用方差分析和LSD检验。统计图采用SigmaPlot 9.0(Systat Software, Inc.)绘制。据Valladares et al.(2000)的方法来计算相关变量的可塑性指数(plasticity index, PI),可塑性指数PI=(某变量的最大平均值-最小平均值)/最大平均值。可塑性指数范围在0~1之间,数值越大,表明可塑性越大。
2 结果与分析
2.1 遮阴对高山杜鹃叶片表皮特征的影响
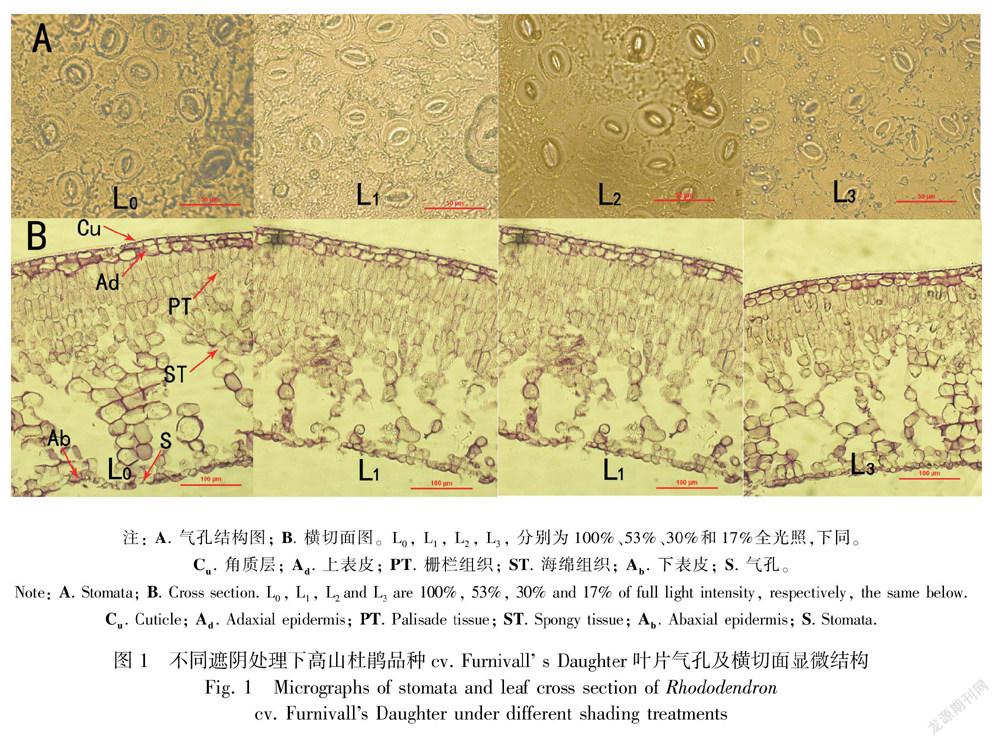
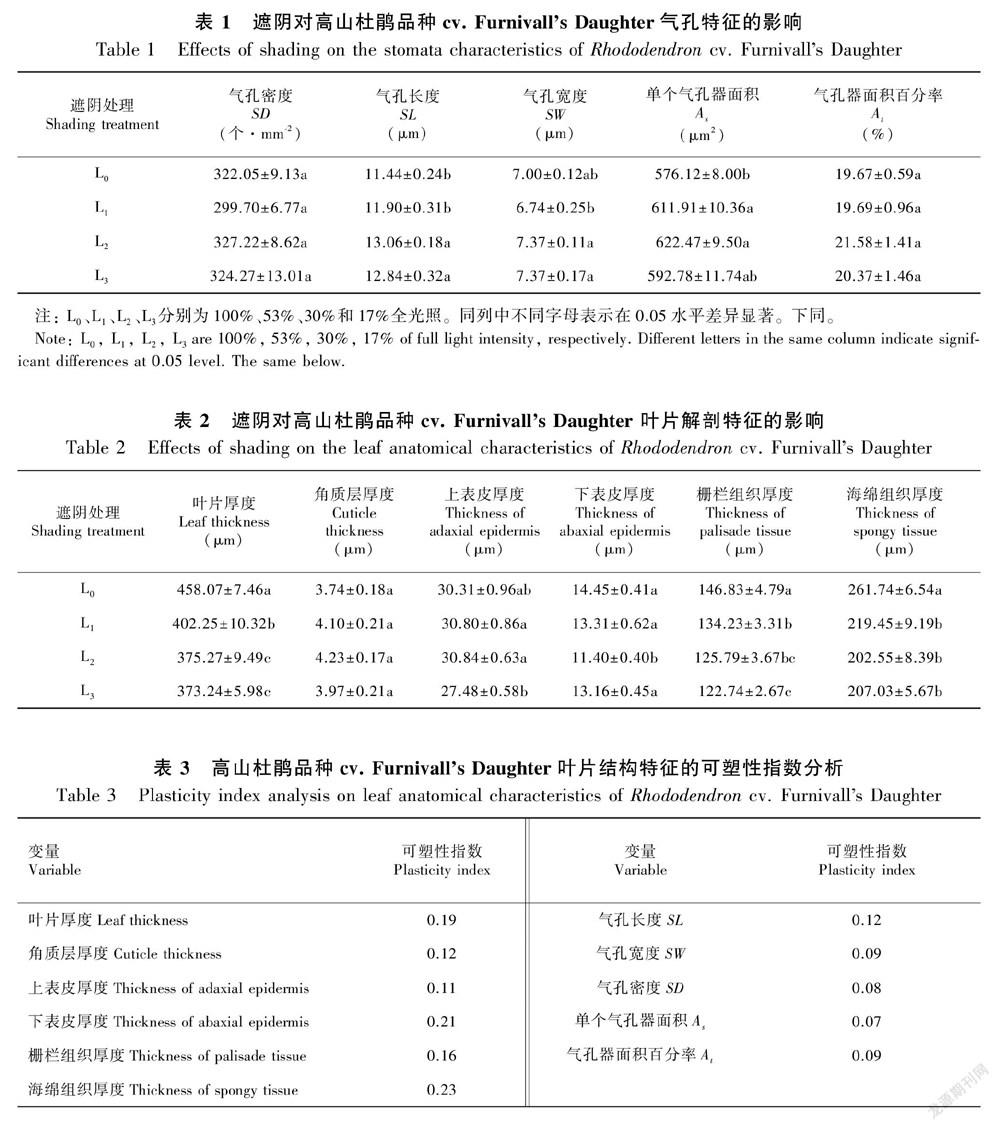
经过对处理植株叶片的上、 下表皮进行观察,上表皮均无气孔分布, 下表皮气孔呈卵圆形或椭圆形(图1:A);4个处理的气孔密度(SD)变化范围在299.70~327.22个·mm-2之间,不同光强下气孔密度未发生显著变化,气孔长(SL)和气孔宽(SW)则随着遮阴程度的增加而增加,单个气孔器面积(As)和总的气孔面积百分比(At)也显著增加,其中以L2处理植株的气孔密度及气孔开度最大(表1)。
2.2 遮阴对高山杜鹃叶片叶肉特征的影响
高山杜鹃品种cv. Furnivalls Daughter叶片解剖结构见图1:B。光照强度对叶片厚度有明显的影响,表现为L0处理植株叶片最厚,分别显著高于L1、L2、L3处理植株13.9%、22.1%、22.7%;叶片上表皮由2层细胞构成,下表皮则由1层细胞构成。上表皮细胞体积较大,上面覆盖有一薄层角质层。随着光强的减弱,上、下表皮厚度逐渐降低,但角质层厚度无显著差异。栅栏组织和海绵组织厚度随光照强度的降低呈显著下降趋势,且对光强下降十分敏感。L0植株栅栏组织厚度分别显著高于L1、L2、L3处理植株9.39%、16.73%、19.63%,海绵组织厚度分别显著高于L1、L2、L3处理植株19.27%、29.22%、26.43%(表2)。
2.3 高山杜鹃叶片结构特征的可塑性指数分析
对4个处理叶片结构特征的可塑性分析表明:叶片厚度、下表皮厚度、栅栏组织厚度和海绵组织厚度的可塑性指数高于0.15,角质层厚度、上表皮厚度、气孔长度的可塑性指数在0.10~0.15之间,其余参数的可塑性指数均小于0.10(表3)。
2.4 遮阴对高山杜鹃光响应曲线的影响
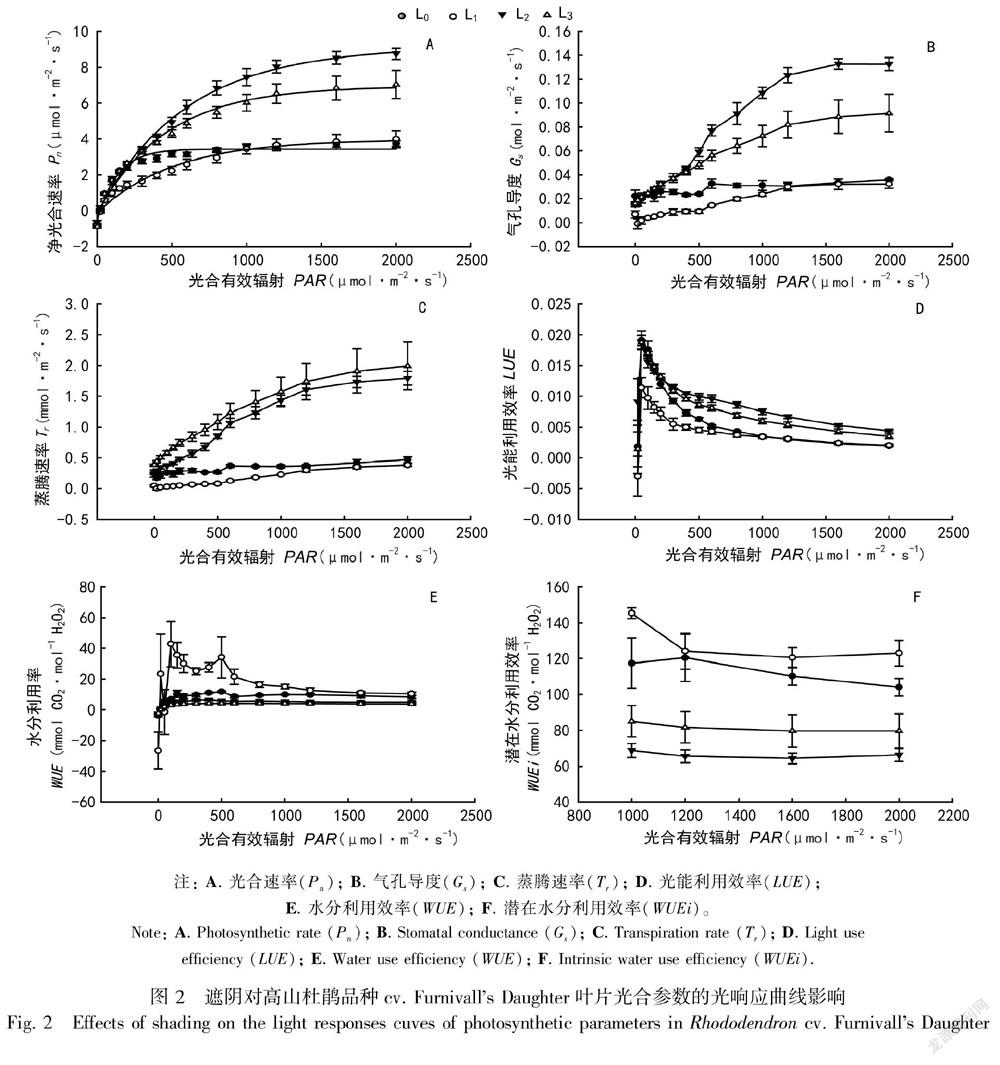
植物光响应曲线是研究植物光合特性的重要指标。4个处理的Pn随光强的变化趋势类似,当PAR低于200 μmol·m-2·s-1时,Pn随着PAR的增加迅速增加,此后随着PAR的增强,Pn增加较为缓慢,在光强达到光饱和点后较为稳定(图2:A)。从图2:H可以看出, 不同处理的Pn出现了显著差异,即L2>L3>L1>L0。
低光强时,Gs和Tr随着光强的增加迅速增加;当光强增大时,Gs和Tr增加缓慢,并在达到光饱和点后最终趋于稳定。4个处理中,Gs表现出L2>L3>L0>L1的趋势(图2:B),Tr则表现出L3>L2>L0>L1的趋势(图2:C),且4个处理间具有显著差异出现,L2和L3处理的Gs和Tr显著高于L0和L1。
在较低的光强下(PAR<50 μmol·m-2·s-1),光能利用效率(LUE)随着光强的增加迅速增加,此后则随着光强的增加逐渐降低。4个处理中,L2处理具有最高的LUE,其次为L3处理,L0和L1处理的LUE接近,且L2和L3處理的LUE显著高于L0和L1处理(图2:D),可见适当的遮阴有效提高了LUE。4个处理的水分利用效率(WUE)和潜在水分利用效率(WUEi)也表现出显著差异,L1处理的WUE和WUEi最大,其次为L0处理,光饱和点后4个处理的WUE和WUEi均随着光强的增加降低,但L0和L1处理的WUE和WUEi显著高于L2和L3处理(图2:E,F)。
2.5 遮阴对高山杜鹃叶片光合参数的影响
从图3可以看出,L2和L1处理分别具有最低和最高的光补偿点,而L0处理的光饱和点最低,显著低于L1、L2和L3处理的67.1%、69.6%和63.5%,L2处理具有最低的光补偿点和最高的光饱和点,表明L2处理具有较大的光能利用范围。L0和L1处理的饱和光合速率显著低于L2和L3处理,其中,L2处理具有最高的饱和光合速率,显著高于其余3个处理。L0处理的暗呼吸速率最高,显著高于其余3个处理,L2处理具有最低的暗呼吸速率。
3 讨论
3.1 遮阴对高山杜鹃叶片解剖结构的影响
叶片是植物进化过程中对环境变化敏感且可塑性较大的器官,在不同的选择压力下可形成各种适应类型,其结构特征最能体现环境因子对植物的影响或植物对环境的适应(Aasamaa et al., 2001)。一般认为,厚叶和发达的栅栏组织是强光、旱生生境叶片的典型特征,而薄叶、发达的海绵组织则是叶片对于弱光、水分供给充足环境的一种适应特性(李芳兰和包维楷,2005)。本研究中,随着遮阴程度的增加,高山杜鹃叶片厚度和叶肉组织厚度呈降低趋势。光照最强的L0处理植株具有较厚的叶肉组织,不仅能保持植株体内水分,还具有较强的折光性,防止过强光照引起的伤害。叶肉组织厚度增加,叶肉细胞面积扩大,细胞中叶绿体增加,有利于维持较高的光合能力,但L0植株厚的叶肉组织增加了CO2从气孔下室到光合作用场所的传导阻力,限制了光合同化物的积累,从而降低了叶片的光合速率(Niinemets,1999;马慧丽和吕德国,2014)。L2和L3处理植株的栅栏组织和海绵组织厚度显著小于L0植株,水分和CO2的传导路径缩短,使CO2和水分的交换更容易进行,从而使植株具有较大的光合速率。
葉片气孔和叶表皮是植物水分蒸腾的主要部位,气孔的大小和密度与植物光合作用及水分利用效率密切相关(戴凌峰等,2008;史刚荣和蔡庆生,2006)。一般来说,生长在高光强下的植物通常比低光强下的具有较高的气孔密度,弱光环境下叶片气孔密度减小、排列疏松、气孔开张度和气孔阻力均较对照显著降低(罗俊等,2006;贺安娜等,2012)。本研究未得到完全相同的结论,光照强度的变化对高山杜鹃气孔密度的影响不明显,但对气孔大小及单个气孔器的面积影响显著。光照较弱的L2和L3处理植株的气孔开度较大,眭晓蕾等(2009)对辣椒幼苗的研究得到了类似结果。较大的气孔开度,使水分和CO2更容易进出,从而使Gs和Tr显著高于光照较强的L0和L1处理植株,有效提高水分和CO2进入光合器官的速率,表现在其光合作用主要特征参数Pmax、Pn在4个处理中均较高,有利于碳的净积累。光照最强的L0植株的气孔及气孔器最小,能快速调控气孔的开关,当光强过强时,气孔部分关闭,使Gs迅速下降,有效避免严重的蒸腾失水,这是气孔对高光照环境适应的又一表现。气孔关闭后,叶表皮是水分散失的主要通道,表皮上的角质层能防止水分过度蒸发。本研究中,随光照强度的降低,4个处理间的角质层厚度未出现显著差异,而上、下表皮厚度则逐渐降低,有效增加叶内光强度,有利于增强对漫射光和散射光的接受能力,提高光能利用效率,是植物对低光环境的一种适应(覃凤飞等,2012)。
为了评价叶片解剖结构特征参数在适应不同光环境中的作用,对高山杜鹃叶片结构特征参数进行了可塑性分析,结果表明与叶肉相关的参数如叶片厚度、下表皮厚度、栅栏组织厚度和海绵组织厚度等的可塑性指数较高,而与气孔相关的参数如气孔密度、气孔长度、气孔宽度等的可塑性指数较低,这表明叶肉组织在高山杜鹃适应不同光强度的过程中起了较为重要的作用,而气孔在其适应光环境的变化过程中作用较小。
3.2 遮阴对高山杜鹃光合特性的影响
光饱和点和光补偿点可以作为衡量植物需光特性的生理指标,分别代表植物对光照强度耐受性的上限和下限,其范围可以反映植物对光能的利用能力(尚海琳等,2008)。本研究中,遮阴处理对高山杜鹃的光饱和点和光补偿点影响较大。其中,L2处理植株具有最低的LCP和最高的LSP,表明其对光能的利用范围最广,这是该处理植株具有最大净光合速率的重要原因之一;此外,L2处理植株的Rd显著低于其他3个处理,表明其以较少的净光合消耗来维持碳平衡,这对植物体内有机物质的积累及碳平衡的维持具有重要意义(蔡艳飞等,2011),在实验中也观察到,L2处理植株的长势最好,叶片嫩绿。从光合速率的光响应曲线可以看出:在光强超过光饱和点以后,其光合速率趋于稳定没有出现显著的下降,这说明短时间的强光照不会对高山杜鹃的光合器官造成损伤。然而,长期的强光照会对光合器官造成严重损伤并产生光抑制(Hanba et al., 2002;Raven,2011;Li et al., 2008),如在本研究中,100%全光照条件下,晴天中午的光合有效辐射最强在2 000 μmol·m-2 ·s-1左右,其光强在较长时间内大大高于光饱和点(447.41 μmol·m-2·s-1),在该条件下叶片难以消耗过剩光能,过剩光能损伤光合器官产生光抑制,直观表现在该条件下高山杜鹃生长不好,叶片发黄并伴有较多的枯叶,光合速率仅为光强较为合适的L2处理的39.18%,说明长时间的强光照射造成了高山杜鹃的逆境环境。
光能利用效率是衡量植物对光能利用效率高低的重要指标。4个处理中,L2处理具有最高的LUE,且L2和L3处理植株的LUE显著高于L0和L1处理。这可能与低强度的光辐射下红蓝光之比的增加刺激了光化学反应和气孔的开张,从而使其具有较高的光合速率有关(张长芹等,1993)。此外,L2和L3处理植株的上、下表皮厚度、叶片厚度均显著低于L0和L1处理植株,较薄的叶片有利于漫射光和散射光在叶肉细胞内的传导,能有效提高植物在低光环境下的光能利用效率。WUE是较稳定的衡量碳固定与水分消耗比例的良好指标。L2和L3处理的WUE和WEUi均比光照较强的L0和L1处理低,而在任何光照强度下L2和L3处理的Pn远远高于L0和L1处理,这就是说L2和L3处理植株以更多的水分消耗来维持其高的光合效率,这与L2和L3处理植株具有较大的气孔密度和气孔开度密切相关。此外,L2和L3处理的Gs和Tr都显著高于L0和L1处理,但其WUE明显低于后者,表明叶片气孔导度对高山杜鹃叶片气体交换和水分利用具有明显的调控作用。
综上所述,云南昆明地区高山杜鹃春季最适光环境为30%左右的全光照,在该条件下高山杜鹃的叶片具有较大的气孔开度,适宜的叶片厚度,有利于CO2和水分的交换。同时,还具有最高的光合能力、较低的LCP、Rd和较高的LSP、Gs、Tr。因此,在高山杜鹃的栽培及应用中,应采取适当的遮阴措施以创造其适宜生长的最佳光环境。
参考文献:
AASAMAA K, SOBER A, RAHI M, 2001. Leaf anatomical characteristics associated with shoot hydraulic conductance, stomatal conductance and stomatal sensitivity to changes of leaf water status in temperate deciduous trees [J]. Aust J Plant Physiol, 28(8):765-774.
ALERIC KM, KIRKMAN LK, 2005. Growth and photosynthetic responses of the federally endangered shrub, Lindera melissifolia, to varied light environments [J]. Am J Bot, 92(4):682-689.
CAI YF, LI SF, XIE WJ, et al., 2011. Influences of growth environments on photosynthetic characteristics and leaf traits of Clematis variety ‘Vyryan Pennell’ [J]. Acta Hortic Sin, 3(7):1377-1384. [蔡艳飞, 李世峰, 解玮佳, 等, 2011. 不同光照环境对‘薇安’铁线莲光合特性的影响 [J]. 园艺学报, 3(7):1377-1384.]
CAO XJ, LIU JJ, YANG M, 2009. Photosynthetic characteristics and anatomical structure of five species of Rhododendron in the Taibai Mountain [J]. Acta Bot Boreal-Occident Sin, 29(12):2483-2491. [曹晓娟, 刘建军, 杨梅, 2009. 太白山5种杜鹃属植物叶片光合特性及解剖结构的生态适应性研究 [J]. 西北植物学报, 29(12):2483-2491.]
CHANDRA S, 2003. Effects of leaf age on transpiration and energy exchange of Ficus glomerata, a multipurpose tree species of central Himalayas [J]. Physiol Mol Biol Plants, 9(5):255-260.
DAI LF, CUI LJ, ZHANG ZX, 2008. Influence of shading treatment on growth of Jatropha curcas seedling [J]. J Anhui Agric Sci, 36(14):5729-5731. [戴凌峰, 崔令军, 张志翔, 2008. 遮阴处理对小桐子幼苗生长的影响 [J]. 安徽农业科学, 36(14):5729-5731.]
EVANS JR, 1989. Photosynthesis and nitrogen relationships in leaves of C3 plants [J]. Oecologia, 78(1):9-19.
HANBA YT, KOGAMI H, TERASHIMA I, 2002. The effect of growth irradiance on leaf anatomy and photosynthesis in Acer species differing in light demand [J]. Plant Cell Environ, 25(8):1021-1030.
HE AN, OU LJ, LI SH, et al., 2012. Effect of shading on the photosynthetic characteristics of Saxifrage stolonifera Curt [J]. Bull Bot Res, 33(5):587-592. [贺安娜, 欧立军, 李胜华, 等, 2012. 虎耳草不同光温条件下光合特性及有效成分含量的相关性分析 [J]. 植物研究, 33(5):587-592.]
JAMES SA, BELL DT, 2011. Leaf morphological and anatomical characteristics of heteroblastic Eucalyptus globulus ssp. globulus (Myrtaceae) [J]. Aust J Bot, 49(2):259-269.
LI FL, BAO WK, 2005. Responses of the morphological and anatomical structure of the plant leaf to environmental change [J]. Chin Bull Bot, 22(Z1):118-127. [李芳兰, 包维楷, 2005. 植物叶片形态解剖结构对环境变化的响应与适应 [J]. 植物学通报, 22(增刊):118-127.]
LI Q, XIAO JZ, LI ZB, et al., 2009. Physiological and biochemical research on critical period of floral initiation in Rhododendron hybrids [J]. J Agric Univ Hebei, 32(1):47-50. [李倩, 肖建忠, 李志斌, 等, 2009. 高山杜鵑花芽分化临界期生理生化研究 [J]. 河北农业大学学报, 32(1):47-50.]
LI ZB, BAI XX, LI P, 2007. The alpine rose will become our country top grade landscaping new favorite — the past, now, future of alpine rose studies [J]. Agric Sci Inform, 7(1):82-85. [李志斌, 白霄霞, 李萍, 2007. 高山杜鹃将成为我国高档园林绿化的新秀——高山杜鹃研究的过去·现在·未来 [J]. 农业科技与信息(现代园林), 7(1):82-85.]
LI ZR, ZHANG SB, HU H, et al., 2008. Photosynthetic performance along a light gradient as related to leaf characteristics of a naturally occurring Cypripedium flavum [J]. J Plant Res, 121(6):559-569.
LUO J, ZHANG H, CHEN YQ, et al., 2006. Relationship of energy sugarcane leaf forms and gas exchange with its yield [J]. Chin J Appl Environ Biol, 12(6):754-760. [罗俊, 张华, 陈由强, 等, 2006. 能源甘蔗不同叶位叶片形态、光合气体交換及其与产量关系 [J]. 应用与环境生物学报, 12(6):754-760.]
MA HL, L DG, 2014. Effects of light condition on structure and photosynthetic characteristics of leaves in ‘Hanfu’ apple [J]. Chin J Appl Ecol, 25(7):1927-1932. [马慧丽, 吕德国, 2014. 光照条件对‘寒富’苹果叶片结构和光合特性的影响 [J]. 应用生态学报, 25(7):1927-1932.]
NIINEMETS , 1999. Components of leaf dry mass per area-thickness and density-alter leaf photosynthetic capacity in reverse directions in woody plants [J]. New Phytol, 144(1):35-47.
NIINEMETS , TENHUNEN JD, 1997. A model separating leaf structural and physiological effects on carbon gain along light gradients for the shade-tolerant species Acer saccharum [J]. Plant Cell Environ, 20(7):845-866.
PANDEY SK, SINGH H, SINGH JS, 2009. Species and site effects on leaf traits of woody vegetation in a dry tropical environment [J]. Curr Sci, 96(8):1109-1114.
PRIOUL JL, CHARTIER P, 1977. Partitioning of transfer and carboxylation components of intercellular resistance to photosynthetic CO2 fixation:A critical analysis of the methods used [J]. Ann Bot, 41(4):789-800.
QIN FF, LI Q, CUI ZM, et al., 2012. Leaf anatomical structures and ecological adaptabilities to light of three alfalfa cultivars with different fall dormancies under shading during overwintering [J]. Chin J Plant Ecol, 36(4):333-345. [覃凤飞, 李强, 崔棹茗, 等, 2012. 越冬期遮阴条件下3个不同秋眠型紫花苜蓿品种叶片解剖结构与其光生态适应性 [J]. 植物生态学报, 36(4):333-345.]
QIU ZF, ZENG BS, GUO GS, et al., 2017. Effects of various LED lights on growth and development of Tectona grandis plantlets in vitro [J]. Guihaia, 37(5):592-598. [裘珍飞, 曾炳山, 郭光生, 等, 2017. 不同光源对柚木组培苗生长发育的影响 [J]. 广西植物, 37(5):592-598.]
RAVEN JA, 2011. The cost of photoinhibition [J]. Physiol Plant, 142(1):87-104.
RONG L, CHEN X, WANG XC, 2009. Leaf anatomical characters and its ecological adaptation of 13 species of Rhododendron in Baili Azalea Area [J]. J Anhui Agric Sci, 37(3):1084-1088. [容丽, 陈训, 汪小春, 2009. 百里杜鹃杜鹃属13种植物叶片解剖结构的生态适应性 [J]. 安徽农业科学, 37(3):1084-1088.]
ROTHSTEIN DE, ZAK DR, 2001. Photosynthetic adaptation and acclimation to exploit seasonal periods of direct irradiance in three temperate, deciduous-forest herbs [J]. Funct Ecol, 15(6):722-731.
SHANG HL, LI FM, LIN Y, et al., 2008. Photosynthetic cha-racteristics of Sinopodophllum hexandrum from different distribution areas in China [J]. Acta Bot Boreal-Occident Sin, 28(7):1440-1447. [尚海琳, 李方民, 林玥, 等, 2008. 桃儿七光合生理特性的地理差异研究 [J]. 西北植物学报, 28(7):1440-1447.]
SHI DH, CHEN X, 2005. Study on the leaf blade structure of six species of Rhododendron [J]. Guizhou Sci, 23(3):39-45. [石登红, 陈训, 2005. 6种杜鹃花属(Rhododendron)植物叶片结构的研究 [J]. 贵州科学, 23(3):39-45.]
SHI GR, CAI QS, 2006. Leaf anatomic plasticity of white clover and its response to different light intensities [J]. Acta Agr Sin, 14(4):301-305. [史刚荣, 蔡庆生, 2006. 白三叶叶片解剖可塑性及其对光强的响应 [J]. 草地学报, 14(4):31-305.]
SOFO A, DICHIO B, MONTANARO G, et al., 2009. Shade effect on photosynthesis and photoinhibition in olive during drought and rewatering [J]. Agric Water Manag, 96(8):1201-1206.
SUI SL, MAO SL, WANG LH, et al., 2009. Response of anatomical structure and photosynthetic characteristics to low light in leaves of capsicum seedlings [J]. Acta Hortic Sin, 36(2):195-208. [眭晓蕾, 毛胜利, 王立浩, 等, 2009. 辣椒幼苗叶片解剖特征及光合特性对弱光的响应 [J]. 园艺学报, 36(2):195-208.]
VALLADARES F, WRIGHT SJ, LASSO E, 2008. Plastic phenotypic response to light of 16 congeneric shrubs from a panamanian rainforest [J]. Ecology, 81(7):1925-1936.
WANG X, ARORA R, HORNER TH, et al., 2008. Structural adaptations in overwintering leave of thermonastic and nonthermonastic Rhododendron species [J]. J Am Soc Hortic Sci, 133(6):768-776.
WANG YP, LIU SL, CHEN YZ, et al., 2012. Leaf structural characteristics of three wild Rhododendron plants and their adaptability to Changbai Mountains’ northeastern China [J]. J Beijing For Univ, 34(4):18-25. [王艷萍, 刘胜利, 陈玉珍, 等, 2012. 3种长白山高山杜鹃叶片结构及其对环境的适应性 [J]. 北京林业大学学报, 34(4):18-25.]
WU ZH, YU LF, YAN LB, et al., 2018. Responses of Akebia trifoliate leaf anatomical structure and photosynthetic characteristic to drought stress [J]. J Southern Agric, 49(6):1156-1163. [吴正花, 喻理飞, 严令斌, 等, 2018. 三叶木通叶片解剖结构和光合特征对干旱胁迫的响应 [J]. 南方农业学报, 49(6):1156-1163.]
ZHANG CQ, ZHANG H, ZHANG NY, et al., 1993. The effect of light of different colour on growth and photosynthesis on Rhododendron irroratum [J]. Acta Bot Yunnan, 15(4):392-394. [张长芹, 张禾, 张能义, 等, 1993. 不同光质对露珠杜鹃生长发育和光合作用的影响 [J]. 云南植物研究, 15(4):392-394.]
ZHAO B, FU NF, XIANG YC, et al., 2017. Effects of light intensity and planting substrates on the growth of Begonia ningmingensis ‘Ningming Silver’, a new begonia cultivar [J]. Guihaia, 37(9):1153-1160. [赵斌, 付乃峰, 向言词, 等, 2017. 光照强度及栽培基质对秋海棠新品种‘宁明银’生长的影响 [J]. 广西植物, 37(9):1153-1160.]
ZHENG BQ, WANG Y, FENG YJ, 2010. Flower regulation of Rhododendron in Beijing area [J]. Chin Flowers Hortic, 10(1):34-36. [郑宝强, 王雁, 冯艺佳, 2010. 北京地区高山杜鹃花期调控 [J]. 中国花卉园艺, 10(1):34-36.]
