古林箐秋海棠叶斑结构对叶色的影响
2019-09-10杜文文崔光芬王继华段青马璐琳贾文杰王祥宁
杜文文 崔光芬 王继华 段青 马璐琳 贾文杰 王祥宁


摘 要:該研究以古林箐秋海棠(Begonia gulinqingensis)为材料,通过分析叶片形态特征、上表皮光学特性、组织结构、叶绿素含量及叶绿素荧光参数(Fv/Fm),探讨了叶片色斑的形成原因。结果表明:(1)古林箐秋海棠叶斑发生频率和数量无明显规律,但发生部位相对稳定,叶斑主要发生在正对叶柄的两条主脉之间。(2)斑区有两种光反射模式,点状反射和多角形反射,栅栏组织细胞呈近等轴的圆形,排列疏松,与上表皮细胞间存在空隙;非斑区只有点状反射模式,栅栏组织细胞为漏斗型,排列紧密,与上表皮细胞间不存在空隙。(3)斑区和非斑区叶绿体均有密集的堆积基粒和丰富的类囊体膜,斑区叶绿素a、b及总叶绿素含量仅比非斑区分别低24.9%、25.2%、25.1%。(4)叶绿素荧光参数(Fv/Fm)值斑区为0.793,非斑区为0.790。虽然斑区叶绿素含量比非斑区略低,但叶绿体结构完整,且叶绿素荧光参数与非斑区无显著差异。斑区上表皮与栅栏组织细胞间的空隙可使光线到达绿色组织时发生二次反射,在叶片表皮细胞边缘形成白色多边形光反射使该区域相对周围正常叶片区域偏白,基于上述结果可推测古林箐秋海棠的淡绿色块斑形成与特殊的叶片结构有关。
关键词:古林箐秋海棠, 表皮光学特性, 叶片结构, 叶绿素含量,叶绿素荧光参数(Fv/Fm)
中图分类号:Q944
文献标识码:A
文章编号:1000-3142(2019)06-0812-09
Abstract:Using Begonia gulinqingensis as experimental material, the causes of variegated leaves were discussed through analyzing leaf morphological feature, optical properties of adaxial surface, tissue structure, chlorophyll content and chlorophyll fluorescence parameter (Fv/Fm). The results were as follows:(1) The frequency and number of the patch on a leaf had no obvious regularity. In the same plant, some leaves had patches, some had none, the variegated leaf usually had only one patch, while sometimes also had two or several ones, and the occurrence site of the patch was relatively stable, which often occured between the two main veins opposite the petiole. (2) There were two kinds of light reflection patterns in variegated areas, spotted pattern(SP) and polygonal pattern(PP). The cells of palisade tissue were nearly equiaxed and arranged loosely, with intercellular space between the abaxial epidermis cells and palisade tissue cells. The non-variegated areas only had the spotted pattern reflection and the palisade tissue cells were funnel-shaped, arranged closely, there was no intercellular space between theabaxial epidermis cells and palisade tissue cells. (3) Chloroplasts from both variegated areas and non-variegated areas showed dense stacking of grana and stroma thylakoid membranes, chlorophyll a, b and total chlorophyll contents in the variegated areas were only 24.9%, 25.2% and 25.1% lower than those in the non-variegated areas respectively. (4) Chlorophyll fluorescence parameter (Fv/Fm) value was 0.793 in variegated areas, and non-variegated areas was 0.790. Although the contents of chlorophyll in the variegated areas were slightly lower than those in the non-variegated areas, the chloroplast structure was intact, and chlorophyll fluorescence parameter (Fv/Fm) did not differ significantly between the variegated areas and the non-variegated areas. The intercellular space between the abaxial epidermis cells and palisade tissue cells could cause secondary reflection when the light reached the green tissue, and white polygon light reflection was formed at the edge of the epidermal cell, which made this area was whiter than the surrounding normal areas. Based on the above results it can be inferred that the pale green patch on a leaf in B. gulinqingensis is related to a special leaf structure.
Key words:Begonia gulinqingensis, adaxial surface optical properties, leaf structure, chlorophyll content, chlorophyll fluorescence parameter (Fv/Fm)
秋海棠(Begonia L.)是世界上广为栽培的重要园艺植物,花叶兼美,其丰富的叶色在植物中不多见,15 000多种园艺品种多是斑叶类。我国有丰富的秋海棠野生资源,其中斑叶种类约占41.4%,叶斑的大小、形状、位置、颜色多样,一些种类就是根据斑纹而命名,如花叶秋海棠(B.cathayana)、变色秋海棠(B.versicolor)等(崔卫华和管开云,2013)。植物叶斑(variegation)是指植物叶面上出现异色斑点或条纹(李孝辉,2010)。Hara(1957)研究了24个科的55个斑叶种,将叶斑产生的原因分为两类4种:色素型(包括叶绿素型和色素型)和结构型(包括表皮型和空隙型)。目前,研究最多的是色素型叶斑(Aluru et al., 2001;Jiang et al., 2004),而对结构型叶斑研究的较少(Fooshee & Henny,1990;Tsukaya et al., 2004)。叶片是植物进行光合作用最重要的部位,叶片中色素含量及组织结构的变化会引起叶色变化,同时植物光合生理指标也会有所变化(王振兴等,2016)。
叶绿素型叶斑主要是由于叶绿体结构发生变异(基粒片层降解、基质片层减少、叶绿体无膜结构、无类囊体、产生嗜锇颗粒、叶绿体结构不完整等),叶绿素合成受阻,导致叶片呈白色或黄色,叶斑的组织结构与正常绿色部位无明显差异,但叶绿素含量明显低于正常绿色部位,叶绿素荧光参数(Fv/Fm)降低(范燕萍等,2006;常青山等,2008;王啸晨等,2012;卫晶晶,2013;杨海芸,2015;李卫星等,2016)。色素型叶斑是由于花青素的存在而使叶片成红、紫等彩色,斑叶叶片结构与正常绿色叶片无明显差异,叶绿素含量偏低,花青素含量则较高(袁明等,2010;杜慶鑫等,2017)。表皮细胞变异使光线发生干涉、衍射和折射等,空隙结构存在使光线发生二次反射,两者都会改变入射光在叶表和叶片内的光路, 进而影响叶片的吸收光谱和反射光谱,最终使叶片呈现蓝晕色、白色、银白色、淡绿色或银绿色,从而形成结构型叶斑,属于物理色(Zhang et al., 2009;王卅等,2012;Sheue et al., 2012)。结构型叶斑的叶片斑区部位结构与正常绿色部位不同,有些植物叶表皮细胞变厚(Hara,1957),有的表皮存在薄膜干涉滤器、多层薄膜结构或晕色体等微结构(王卅等,2012),广西落檐、仙客来、意大利魔芋的栅栏组织细胞与上表皮存在空隙,结构型叶斑叶绿素含量斑区部位一般比非斑区部位略低,叶绿素荧光参数(Fv/Fm)有的植物斑区比非斑区略大,也有些植物斑区比非斑区的略小,但无显著差别(Tsukaya et al., 2004;Konoplyova et al., 2008;Rocca et al., 2011;Sheue et al., 2012)。近来有研究表明植物的叶色可以有多种机制共同起作用,如色素含量和空隙结构共同影响猕猴桃的叶色(王振兴等,2016),表皮细胞、细胞间隙、叶肉细胞、叶绿体变异和晶体共同作用强化柏拉木幼苗的白色斑点(Chen et al., 2017)。
丰富多彩的叶斑不仅能改良植物观赏性状,还能提高植物观赏价值和经济价值。古林箐秋海棠(Begonia gulinqingensis)为常绿草本,叶片圆形,腹面青翠浓绿,背面略带红色,同株叶片或具形状不规则的淡绿色斑块,或无斑块,叶斑在幼叶萌出时已有。该种为云南特有种,目前已处于濒危状态,其独特的叶斑样式,在观叶秋海棠中并不多见,具有极高的观赏与开发价值(马宏等,2005)。目前,对古林箐秋海棠叶斑的研究鲜见报道,本研究以古林箐秋海棠为材料,研究叶斑的组织结构特征、叶绿素含量及叶绿素荧光参数(Fv/Fm),探讨叶斑形成原因,为阐明叶斑的形成机理提供理论依据。
1 材料与方法
1.1 材料
古林箐秋海棠(Begonia gulinqingensis)种植于云南省农业科学院花卉研究所球宿根花卉育种基地,本研究以有斑叶片上的斑区和非斑区组织为材料。
1.2 方法
1.2.1 叶斑形态特征观察 自2016年4月开始观察古林箐秋海棠叶斑的发生频率、数量、部位,并采集有斑叶片和无斑叶片进行扦插,统计扦插苗叶片的斑块发生情况。
1.2.2 叶片表皮光学特性观察 将叶片擦拭干净后置于滤纸上,用配有汞灯的解剖镜于暗室内观察(Zhang et al., 2009),表皮亮度和表皮细胞反射模式能够反映叶片光学特性。
1.2.3 组织结构观察 半薄切片的制作参照巩玥等(2014)的方法, 取成熟有斑叶片的斑区和非斑区部位,用刀片取1 mm × 1 mm × 1 mm的样品,重复3次,采用2.5%戊二醛固定,0.1 mol·L-1PBS缓冲液浸洗3次, 每次15 min;1%锇酸固定1.5~2 h,再用0.1 mol·L-1PBS缓冲液漂洗3次,每次15 min,30%、50%、70%、80%、90%、95%、100%乙醇脱水各1次,每次15 min;100%乙醇二次脱水,20 min;100%丙酮脱水,20 min;包埋剂∶丙酮(1∶1)室温下置换1 h,包埋剂∶丙酮(3∶1)室温下置换3 h;纯包埋剂过夜,之后将包埋剂移至0.5 mL离心管或包埋具中,再放样品进去,然后放入聚合箱中,70 ℃ 12 h或者60 ℃ 24 h两次,切片厚度为1 μm,甲苯胺蓝染色,显微镜观察并照相。
1.2.4 超微结构观察 方法同半薄切片,切片厚度50~70 nm,醋酸双氧铀先染5~10 min,冲洗,柠檬酸铅后染5 min,再冲洗,烘烤灯烘烤后透射电镜观察、拍照。
1.2.5 叶绿素含量测定 叶绿素含量测定参照李合生等(2000)方法称取叶片(去除叶脉)0.2 g并剪碎,由于一个叶片中斑区质量不足0.2 g,本文采用混合不同植株不同叶片斑区取样,从10棵植株上取12片带有斑块的叶片,4片叶为一组,称取0.2 g斑区和非斑区叶片,将取好的叶片置于研钵中,加入少量石英砂和碳酸钙粉以及95%乙醇溶液2~3 mL,研磨至匀浆;经过滤、冲洗后装入容量瓶用乙醇溶液定容至25 mL;以分光光度计测量叶绿素溶液在665、649和470 nm处的吸光度,计算叶绿素浓度。每个样本重复6次,浓度值取平均值。
1.2.6 叶绿素荧光参数(Fv/Fm)测定 运用叶绿素荧光成像仪测定古林箐秋海棠叶片非斑区和斑区的荧光参数。取古林箐秋海棠成熟带斑叶片,锡箔纸包裹叶片暗处理20 min,测定叶绿素荧光参数(Fv/Fm),包括暗适应状态下的最小荧光(Fo)、暗适应状态的最大荧光(Fm), Fv=Fm-Fo,重复9次。
2 结果与分析
2.1 叶斑形态观察
前期观察发现,古林箐秋海棠不同植株间叶斑的发生频率和数量无明显规律,但发生部位相对稳定。同一植株上同时生长有斑叶片和无斑叶片,用有斑和无斑叶片分别进行扦插,扦插苗均会长出有斑和无斑的叶片,并且有斑叶片上的叶斑数量不定, 通常为一个,也有两个或多个。当叶片上只有一个叶斑时,叶斑常发生在正对叶柄的两条主脉之间(图1:A),当叶片上有两个或多个叶斑时,总会有一个叶斑发生在正对叶柄的主脉间,其余叶斑则分布在其周围的主脉间,极少有叶斑跨越主脉(图1:B,C)。叶斑自幼叶萌发时就带有,随着叶片长大,叶斑面积也逐渐增加,并在叶片完全长大时叶斑大小和形状趋于稳定,有斑叶片的非斑区域与无斑叶片的颜色一致,腹面均呈浓绿(图1:D),背面叶脉为红色(图1:E)。
2.2 叶表面光学特性
通过体视镜观察(图2:A,B)发现,古林箐秋海棠的有斑叶片有两种光反射模式:点状反射(spotted pattern,SP)和多角形反射(polygonal pattern,PP)。斑區淡绿色部位的上表皮细胞不仅可以观察到点状反射(SP),而且还可观察到表皮细胞周围存在较弱的亮圈,这些亮圈在一定程度上反映了细胞的形状。因为多数细胞为不规则多边形,所以称这类反射为多角形反射(PP)(图2:C,D);非斑部位的上表皮细胞可观察到细胞中央存在一个亮点,称为点状反射(SP)(图2:C,D)。
2.3 组织结构与超微结构观察
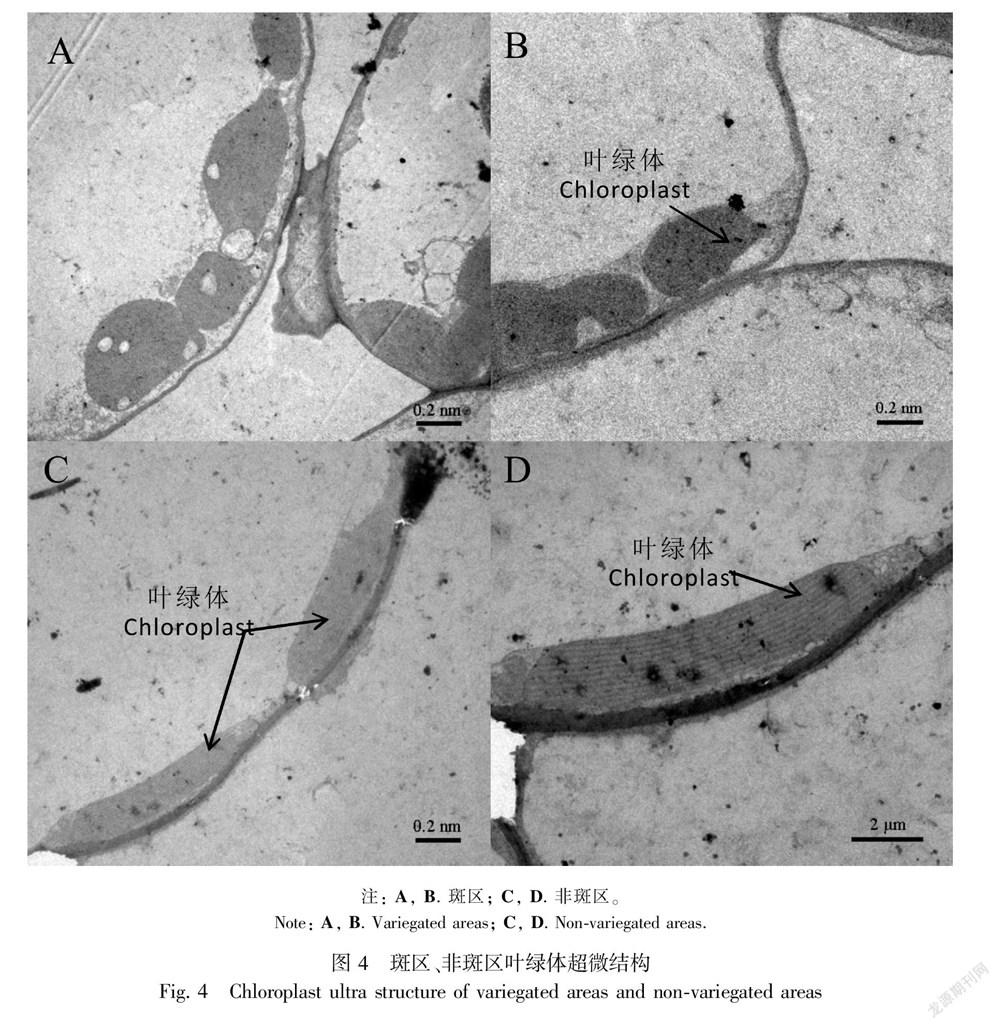
通过半薄切片观察,淡绿色斑区上表皮细胞较扁平,偶有小突起,非斑区细胞表面有长方形突起,形态较斑区略大。斑区栅栏组织细胞较小,呈不规则圆形,排列松散,细胞内含有少量或极少量叶绿体,栅栏组织层与表皮细胞间存在较大的空隙,形成‘气室’,海绵组织细胞大小、形状与栅栏组织细胞类似,两层,排列疏松,均含有叶绿体(图3:A);非斑区栅栏组织细胞较大,呈圆柱形,排列紧密,内含大量叶绿体,与表皮细胞之间无空隙,海绵组织细胞两层,排列紧密,呈不规则圆形,靠近栅栏组织的一层较小,含有叶绿体,下面一层较大,无叶绿体(图3:B)。斑区和非斑区均由5层细胞组成,斑区细胞大小不均匀,排列不整齐,非斑区细胞分层排列,大小均匀且整齐紧密,非斑区细胞略大,所以斑区厚度与非斑区厚度差别不大。
通过超薄切片观察到斑区叶绿体形状呈椭圆形,含有淀粉粒(图4:A,B),非斑区叶绿体细长呈椭圆形,基粒片层清晰(图4:C,D);两部分的叶绿体超微形态虽然不一致,但结构均完整,含有丰富的类囊体膜和密集的堆积基粒。
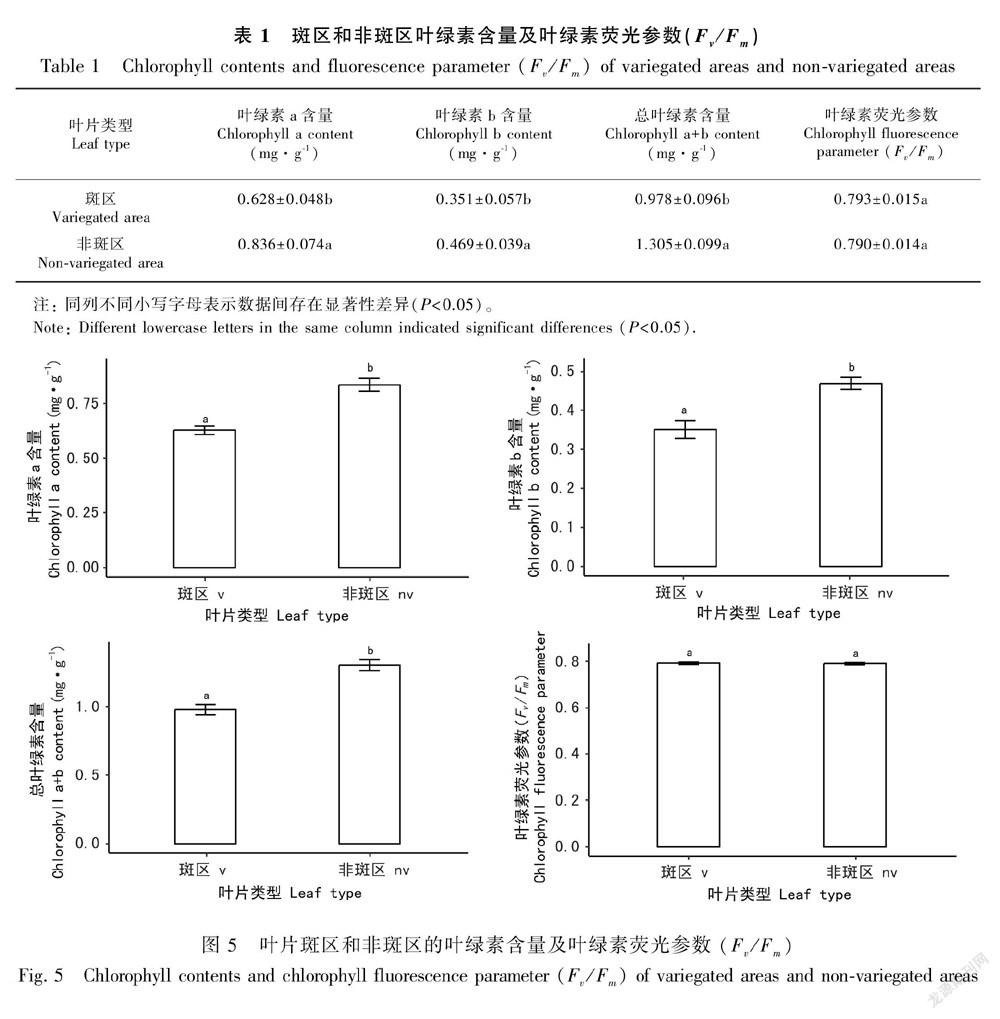
2.4 叶绿素含量和叶绿素荧光参数(Fv/Fm)测定通过叶绿素含量测定发现斑区叶绿素a、b及总叶绿素含量均低于非斑区(表1,图5:A,B,C),斑区叶绿素a、b和总叶绿素含量分别比非斑区低24.9%、25.2%和25.1%。结合半薄切片观察,斑区栅栏组织细胞内所含的叶绿体明显少于非斑区,但其海绵组织细胞中含有较多的叶绿体,弥补了绿色组织细胞内叶绿体的含量。
离体条件下检测斑区和非斑区的叶绿素荧光参数(Fv/Fm)值,斑区为0.793,非斑区为0.790,二者并无显著差别(图5:D),Fv/Fm代表PSⅡ的最大光化学效率,反映了当所有的光系统域(PSⅡ)反应中心均处于开放态时的量子产量,可直接作为原初光化学效率的指标。斑区的叶绿体超微结构完整,虽其总叶绿体含量比非斑区低25.1%,但斑区功能性叶绿体能够充分吸收光能,提高叶片光能利用率,最大光化学效率与非斑区相似,因此古林箐秋海棠在拥有色斑的同时仍能保证正常的光合作用,加上古林箐秋海棠生长在热带雨林中的低层树林下,这种叶色样式是其为适应低光照环境而长期进化的结果。
3 讨论与结论
在前人研究中已有8种秋海棠被发现斑叶中由于空隙结构存在而产生结构型叶斑。大王秋海棠(Begonia rex)、水鸭脚秋海棠(B. formosana)、掌叶秋海棠(B. hemsleyana)、变色秋海棠(B. versicolor)、B. chlorosticta、B. diadema、B. pustulata和Begonia ‘K030960’。7种秋海棠叶斑的空隙结构存在于栅栏组织细胞与上表皮细胞间,栽培种Begonia ‘K030960’斑叶上表皮下方的储水组织细胞与栅栏组织细胞间存在空隙结构(Zhang et al., 2009;Sheue et al., 2012)。本研究观测到的古林箐秋海棠斑叶空隙结构存在于上表皮细胞和栅栏组织细胞间,且斑区的上表皮细胞比非斑区略小。Zhang et al.(2009)和Sheue et al.(2012)在大王秋海棠、掌叶秋海棠、变色秋海棠、B. chlorosticta、B. pustulata和Begonia‘K030960’中未发现斑区和非斑区的表皮细胞存在差异,而Sheue et al.(2012)在水鸭脚秋海棠和B. diadema的斑叶扫描中观察到斑区上表皮细胞比非斑区的略小,说明秋海棠属植物在斑区和非斑区的表皮细胞大小上并没有绝对的特征属性。
古林箐秋海棠斑区的栅栏组织细胞呈圆形,含有较少的叶绿体,非斑区栅栏组织细胞呈漏斗状,含有大量的叶绿体,与Zhang et al.(2009)、Sheue et al.(2012)所述的其他8种斑叶具空隙结构的秋海棠类似,此外,这8种秋海棠非斑区海绵组织细胞排列疏松,与斑区海绵组织细胞大小、形态类似,而在古林箐秋海棠中发现非斑区海绵组织细胞排列紧密,形态、大小与斑区均有明显差异。超微结构显示古林箐秋海棠斑区和非斑区叶绿体结构形状虽然不一致,但均含有丰富的类囊体膜和密集的堆积基粒,而且在斑区还观察到淀粉粒,这与前人的研究一致(Zhang et al., 2009;Sheue et al., 2012;Chen et al., 2017)。
在叶绿素含量方面,古林箐秋海棠斑区叶绿素含量比非斑区略低,但不足以影响叶片的颜色,斑区叶片仍呈现绿色,仅颜色变浅。王振兴等(2016)在狗枣猕猴桃白色叶中也发现此类现象。本研究古林箐秋海棠斑区叶绿素荧光参数(Fv/Fm)与非斑区相差不大,水鸭脚秋海棠、B. diadema和 B. pustulata 3种秋海棠也被报道斑区和非斑区的Fv/Fm值无显著差异(Sheue et al., 2012)。Rocca et al.(2011)通过研究意大利魔芋光系统Ⅱ (PSⅡ) 的活性发现不同颜色的叶片在最大光化学效率(Fv/Fm)上无显著差异;Chen et al.(2017)在柏拉木中发现斑区和非斑区的Fv/Fm无显著差异,但有斑叶片非斑区的Fv/Fm值比无斑叶片大。
综上所述,古林箐秋海棠叶斑属于空隙结构型叶斑,由于空隙结构的存在使光线到达绿色组织时发生二次反射,在叶片表面表皮细胞边缘形成白色多边形的光反射而不是普通叶片表皮细胞中央形成的白色点状光反射,使该区域相对周围的正常叶片偏白,从而形成叶斑(Hara,1957;Zhang et al., 2009)。这与前期的叶片表皮光学特征观察相符,且在大王秋海棠中也有类似发现(Zhang et al., 2009)。另外,Sheue et al.(2012)发现具有空隙结构叶斑的B. chlorosticta叶片边缘呈绿色,叶背红色,腹面暗绿色,叶斑呈淡绿色,这被认为是由于叶背面红色与绿色相结合而使叶腹面呈现深绿色,深绿色在有空隙结构存在的情况下会呈现浅绿色, 而不是白色。古林箐秋海棠幼叶刚萌出时呈红色,之后叶色逐渐转绿,但叶背的叶脉一直为红色,并且其叶斑同样为空隙型结构叶斑,因此推测其淡绿色叶斑显色机理与B. chlorosticta叶斑的显色机理相同。
参考文献:
ALURU MR, BAE H, WU D, et al., 2001. The Arabidopsis immutans mutation affects plastid differentiation and the morphogenesis of white and green sectors in variegated plants [J]. Plant Physiol, 127(1):67-77.
CHANG QS, CHEN FD, TENG NJ, et al., 2008. Chlorophyll contents and chloroplast microstructure and ultrastructure of leaves in yellow-green mutant of Chrysanthemum [J]. Acta Bot Boreal-Occident Sin, 28(9):1772-1777. [常青山, 陈发棣, 腾年军, 等, 2008. 菊花黄绿叶突变体不同类型叶片的叶绿素含量和结构特征比较 [J]. 西北植物学报, 28(9):1772-1777.]
CHEN YS, CHESSON P, WU HW, et al., 2017. Leaf structure affects a plant’s appearance:Combined multiple mechanisms intensify remarkable foliar variegation [J]. J Plant Res,130(2):1-15.
CUI WF, GUAN KY, 2013. Diversity of leaf variegation in Chinese Begonias [J]. Plant Divers Resour, 35(2):119-127. [崔衛华, 管开云, 2013. 中国秋海棠属植物叶片斑纹多样性研究 [J]. 植物分类与资源学报, 35(2):119-127.]
DU QX, QING JZ, JING Y, et al., 2017. Pigments and active ingredients in leaves of seven Eucommia ulmoides variation-types [J]. Bull Bot Res, 37(3):468-473. [杜庆鑫, 庆军朱, 景乐, 等, 2010. 7个变异红叶杜仲叶片色素及活性成分分析 [J]. 植物研究, 37(3):468-473.]
FAN YP, LI HL, LI HJ, 2006. Pigment composition and ultrastructural difference of chloroplast in three kinds of variegation leaf of Cymbidium sinense [J]. J S Chin Agric Univ, 27(2):8-12. [范燕萍, 李慧玲, 李浩健, 2006. 几种花叶线艺兰叶片色斑色素组成和叶绿体超微结构研究 [J]. 华南农业大学学报, 27(2):8-12.]
FOOSHEE WC, HENNY RJ, 1990. Chlorophyll leaves and anatomy of variegated and nonvarigated areas of Aglaonema nitudum leaves [J]. Proc Flor State Hortic Soc, 103:170-172.
GONG Y, CHEN HM, JIANG CD, et al., 2014. Quantification of leaf anatomical structure and its application in a C4 plant, Sorghum [J]. Bull Bot, 49(2):173-182. [巩玥, 陈海苗, 姜闯道, 等, 2014. 植物叶片解剖结构的量化及其在C4植物高粱中的应用 [J]. 植物学报, 49(2):173-182.]
HARA N, 1957. Study of the variegated leaves with special re-ference to those caused by air spaces [J]. Jpn J Bot, 117(6):86-101.
JIANG WB, ZHUANG M, HAN HZ, et al., 2005. Progress on color emerging mechanism and photosynthetic characteristics of colored-leaf plants [J]. Acta Hortic Sin, 32(2):352-358.
KONOPLYVA A, PRTROPOULOU Y, YIOTIS C, et al., 2008. The fine structure and photosynthetic cost of structural leaf variegation [J]. Flora, 203(8):653-662.
LA ROCCA N, RASCIO N, PUPILLO P, 2011. Variegation in Arum italicum leaves:A structural-functional study [J]. Plant Physiol Biochem, 49(12):1392-1398.
LI HS, SHI Q, ZHAO SJ, et al., 2000. The experiment principle and technique on plant physiology and biochemistry [M]. Beijing:Higher Education Press:134-137. [李合生, 石群, 赵世杰, 等, 2000. 植物生理生化实验原理与技术 [M]. 北京:高等教育出版社:134-137.]
LI XH, LI MY, ZHANG XS, et al., 2010. An investigation of the variegated plant resources in Longmen Mountain [J]. J Mianyang Norm Univ, 29(8):61-65. [李孝辉, 李明银, 张学适, 等, 2010. 龙门山区斑叶观赏植物资源调查初報 [J]. 绵阳师范学院学报, 29(8):61-65.]
LI WX, JIANG F, MA JL, et al., 2017. Ultrastrucuture and chlorophyll contents of the variegated leaves in Ginkgo biloba L. [J]. J Yangzhou Univ (Agric Life Sci Ed), 37(1):103-107. [李卫星, 蒋菲, 马江黎, 等, 2016. 银杏斑叶超微结构观察及叶绿素含量分析 [J]. 扬州大学学报(农业与生命科学版), 37(1):103-107.]
MA H, LI HZ, GUAN KY, et al., 2007. Begonia gulinqingensis, a rare plant and its wild resource status and biological characters [J]. J Yunnan Agric Univ, 22(3):337-339. [马宏, 李宏哲, 管开云, 等, 2007. 珍稀植物古林箐秋海棠的资源状况及生物学特性 [J]. 云南农业大学学报, 22(3):337-339.]
SHEUE CR, PAO SH, CHIEN LF, et al., 2012. Natural foliar variegation without costs? The case of Begonia [J]. Ann Bot, 109(6):1065-1074.
TSUKAYA H, OKADA H, MOHAMED M, 2004. A novel feature of structural variegation in leaves of the tropical plant Schismatoglottis calyptrate [J]. J Plant Res, 117(6):477-480.
WANG S, LI YH, ZHANG Y, 2012.Research progress on structural color mechanisms and biological functions of low-light plants in tropical rainforest [J]. Acta Hortic Sin, 39(11):2291-2230. [王卅, 李玉花, 张旸, 2012. 热带雨林低光植物结构色产生机制及生物学功能研究进展 [J]. 园艺学报, 39(11):2291-2230.]
WANG ZX, CAO JR, QIN HY, et al., 2016. Common effect of pigment content and leaf structure on leaf color in Actinidia kolomikta [J]. Plant Physiol J, 52(12):1921-1926. [王振興, 曹建冉, 秦红艳, 等, 2016. 狗枣猕猴桃彩叶色素含量和结构共同影响叶色 [J].植物生理学报, 52(12):1921-1926.]
WANG XC, YUE XH, WU J, et al., 2012. Appearance and structure analysis of chimeric leaves from two ornamental bamboos [J]. Chin Agric Sci Bull, 28(16):233-238. [王啸晨, 岳祥华, 吴杰, 等, 2012. 2种观赏彩叶竹形态结构的观察与分析 [J]. 中国农学通报, 28(16):233-238.]
WANG ZX, YU YF, CHEN L, et al., 2016. Advances in leaf pigment composition, structure and photosynthetic characte-ristics of colored-leaf plants [J]. Plant Physiol J, 52(1):1-7. [王振兴, 于云飞, 陈丽, 等, 2016. 彩叶植物叶片色素组成、结构以及光合特性的研究进展 [J]. 植物生理学报, 52(1):1-7.]
WEI JJ, 2013. Leaf morphological ultrastructure, chlorophyll biosynthesis and photosynthesis of the yellow-leaf mutant of Rosa beggeriana [D]. Beijing:Chinese Academy of Agricultural Sciences. [卫晶晶, 2013. 金叶弯刺蔷薇叶片形态、叶绿素生物合成及光合作用特性的研究 [D]. 北京:中国农业科学院.]
YANG HY, 2015. Study on mechanism of spontaneous leaf color variation of Pseudosasa japonica f. [D]. Beijing:Beijing Forestry University. [杨海芸, 2015. 花叶矢竹叶色变异机理研究 [D]. 北京:北京林业大学.]
YUAN M, WANG XZ, DU L, et al., 2010. Mechanism on leaf color transformation of Loropetalum chinense Olive var. rubrum [J]. Acta Hortic Sin, 37(6):949-956. [袁明, 万兴智, 杜蕾, 等, 2010. 红花檵木叶色变化机理的初步研究 [J]. 园艺学报, 37(6):949-956.]
ZHANG Y, HAYASHI T, HOSOKAWA M, et al., 2009. Metallic lustre and the optical mechanism generated from the leaf surface of Begonia rex Putz [J]. Sci Hortic, 121 (2):213-217.
