龙脑香热带雨林附生苔藓沿宿主垂直梯度的微生境偏好及其指示作用
2019-09-10沈婷宋亮郭新磊
沈婷 宋亮 郭新磊
摘 要:该研究首次借助林冠塔吊调查了西双版纳国家级自然保护区龙脑香热带雨林样地内69棵树13个垂直高度上的附生苔藓植物,结果表明:目标样树上共记录到隶属于25科60属的90种附生苔藓,其中细鳞苔科物种数最多,占比达25.6%。13个垂直高度上共划分出三种生态类型:喜阳苔藓(散生巨树上>45 m的区域),喜阴苔藓(乔木树干上<15 m的区域),广布苔藓(广泛分布于宿主各个垂直高度上,生态位宽),并筛选出对微生境有特殊偏好的17种苔藓指示种(IndVal≥0.7,P<0.05)。随宿主垂直高度的升高,扇型和交织型的苔藓占比降低,悬垂型苔藓占比先升高后降低,细平铺型和粗平铺型的苔藓占比升高。大气湿度、水汽压、胸径以及树皮粗糙度对附生苔藓生活型的分布偏好具有显著影响。总之,沿宿主垂直高度上的附生苔藓对微环境变化在生活型和形态结构上有着不同的响应方式,而同一种生态型的苔藓群落有相似的适应机制。因此,在森林林冠生境变化的监测和管理中,对微生境具有明显偏好的附生苔藓物种可作為有效的指示材料。
关键词:林冠, 附生苔藓, 热带雨林, 垂直分布, 生境偏好, 指示物种
中图分类号:Q948.1
文献标识码:A
文章编号:1000-3142(2019)06-0776-12
Abstract:Epiphytic bryophytes play significant roles in maintaining water and nutrient cycles in the forest ecosystem. However, diversity and distribution patterns of epiphytic bryophytes in forest canopies are understudied because of the limitations of canopy investigation techniques. Epiphytic bryophytes and their life-forms were investigated from 69 trees in thirteen vertical segments using a canopy crane in a tropical dipterocarp rain forest in Xishuangbanna National Nature Reserve. Canonical correlation analysis (CCA) and the species indicator method were applied to analyze the microhabitat preferences of the epiphytic bryophytes and to find indicators along the vertical gradient of the hosts. The main results were as follows:In the study area 90 species, belonging to 60 genera and 25 families, were recorded. Lejeuneaceae was the most species-rich family, representing 25.6% of all bryophyte species recorded. Three special ecological types were extracted from the thirteen vertical segments, including sun-loving bryophytes (>45 m on tree trunks in the forest canopies of emergent trees), shade-loving bryophytes (<15 m on tree trunks) and generalist bryophytes (usually distributed in various host heights with a broad niche width). Seventeen bryophyte species (IndVal≥0.7, P<0.05) were selected as indicators for specific vertical segments. With the increase in the height sampled, the proportion of species with fan and weft life-forms decreased, the proportion of pendant life-forms increased first and then decreased, while the proportion of smooth mat and rough mat life-forms increased. The relative humidity, vapor pressure, diameter at breast height (DBH) and bark roughness had significant influences on the distribution preferences of epiphyte life forms. In conclusion, epiphytic bryophytes show different life-form compositions and morphological structures in response to changes in the micro-environment along the vertical gradient of hosts, while bryophytes assembled in the same micro-habitat usually have similar adaptation mechanisms. The epiphytic species with obvious preferences for specific microhabitats can be used as indicators during the monitoring and management of habitat changes in the forest canopy.
Key words:canopy, epiphytic bryophytes, tropical rain forest, vertical distribution, habitat preference, indicator species
热带森林林冠是生物圈物种丰富度最高的生境之一(Stork, 2007),其独特的环境条件孕育了极其丰富的附生植物,特别是附生苔藓。附生苔藓(epiphytic bryophytes)是指生长在树木或灌木树皮上的苔藓植物(Smith, 1982)。附生苔藓在维持森林生态系统水分和养分循环中发挥着重要的生态作用(Ah-Peng et al., 2017; Shi et al., 2017),并对林冠生物及生境多样性的形成和维持有巨大贡献,因而具有很高的生态保护价值。苔藓植物特别是附生苔藓由于体态微小、没有角质层,且直接暴露在空气中,对生境变化极其敏感,可作为森林生态系统健康状况的生物指示种(Glenn et al., 1988; 吴玉环等,2002)。
热带林冠的复杂性和微生境的多样化孕育了丰富的附生苔藓,使得热带林冠上附着有全世界25%~30%的苔藓物种(Gradstein & Pócs, 1989)。目前,对于热带区域附生苔藓的研究主要集中在热带美洲和热带非洲( Gradstein & Pócs, 1989; Frahm & Gradstein, 1991; Wolf, 1993; Kürschner & Parolly, 1998; Holz et al., 2002),而在热带亚洲开展的研究较少(Kürschner, 1990; Frey et al., 1990; Ariyanti et al., 2008; Sporn et al., 2009, 2010)。亚洲热带雨林不仅孕育了不同于热带美洲和热带非洲雨林的苔藓植物,而且还占据了最多科和属 (Buck & Thiers, 1898; Gradstein & Pócs, 1989)。其中,龙脑香热带雨林是世界上植物群落最高的热带森林(Raes et al., 2014)。西双版纳的龙脑香热带雨林直到1975年才被发现,受到植物学界高度重视(朱华, 2000),该森林以国家一级保护植物望天树(Parashorea chinensis)为优势物种。散生巨树的望天树,高为40~70 m(唐建维等,2008),其高度几乎是热带美洲低地雨林的两倍(Dislich & Mantovani, 2016),为附生植物提供了充足的附生基质和生存空间(Engemann et al., 2016)。此外,林内光照、温度与水分可用性在垂直梯度上发生剧烈变化(Szarzynski & Anhuf, 2001),为附生植物提供了异质的小生境(Woods et al., 2015)。苔藓植物的生长型和生活型可以通过减少空气阻力,增加叶边缘细胞的厚度,形成毛细管空隙及相互保护等方式来帮助保持水分(Glime, 2013),进而适应宿主从树基到林冠顶层梯度性变化的微生境。然而,附生苔藓对于宿主垂直高度上的偏好及其对林冠生境指示作用的研究鲜见报道。
由于林冠访问技术限制,人们对林冠上附生苔藓多样性及其分布格局的研究还很欠缺。附生苔藓植物调查多局限于树干2 m以下的区域(Slack, 1976; Song et al., 2011; Lowman & Schowalter, 2012)。近年来,林冠塔吊(canopy crane)的應用,突破了林冠研究瓶颈(Mitchell et al., 2002),极大地推动了人们对林冠生境及其生物多样性的认识。截止2016年底,国外共有11座林冠塔吊,覆盖森林总面积约11.7 hm2,其中5座位于热带森林(吴毅等,2016)。在中国生物多样性监测与研究网络(Sino BON)“林冠生物多样性监测专项网”的推动下(马克平, 2015, 2016),中国科学院在东北、华南、华中、西南等地的代表性森林类型中拟建立8座林冠塔吊(已建成6座,2座在建)(Nakamura et al., 2017)。其中,在西双版纳龙脑香热带雨林建成有世界最高的林冠塔吊,这为开展林冠附生植物多样性研究提供了坚实的硬件条件(吴毅等,2016)。虽然林冠塔吊已覆盖全球的温带、亚热带、热带森林,但仅有美国Wind River一座塔吊曾被用于非维管植物多样性和空间分布的研究(吴毅等,2016)。
本研究首次运用西双版纳龙脑香热带雨林已建成的林冠塔吊平台,系统调查塔吊样地内附生苔藓植物的组成和分布格局,探讨附生苔藓植物沿宿主垂直梯度的偏好性及其潜在影响因子,揭示附生苔藓对微环境变化的响应与适应机理,筛选出指示特殊微生境的指示种,为苔藓植物的保护和森林资源的管理提供科学依据。
1 研究区域概况
位于东南亚热带雨林北缘的西双版纳国家级自然保护区(101°35′ E, 21°36′ N),拥有我国面积最大、最集中分布的热带雨林,属于中缅生物多样性热点地区(Mittermeier et al., 2004)。在该区域内,中国科学院牵头于2014年在西双版纳傣族自治州勐腊县补蚌村斑马山脚建立了一座能覆盖森林面积约为1 hm2的林冠塔吊。该林冠塔吊塔身高81 m,臂长约为60 m,中心坐标为101°34′56.154″ E,21°37′4.467″ N,占据了以56.4 m为半径的圆形森林样地。该区域属于北热带季风气候区,主要受控于西南季风,年均温为21 ℃左右,全年高温无霜,年降雨量约为1 500 mm,年平均空气相对湿度为86%,且有明显的干湿季之分(Cao et al., 2006; Zhu & Cao, 2006)。
热带望天树林层可分为上、中、下三层(朱华,2000)。该热带样地内,上层乔木(树高大于30 m)所受光照条件最强,主要有望天树(Parashorea chinensis)、毛猴欢喜(Sloanea tomentosa)、橄榄(Canarium album)、绒毛番龙眼(Pometia tomentosa),高度在40~60 m之间的高位芽植物称为散生巨树(朱华,2000);中层乔木(16~30 m)属于群落的郁闭层,受中等强度的光照条件,主要由黑毛柿(Diospyros atrotricha)、青藤公 (Ficus langkokensis)、五桠果叶木姜子(Litsea dilleniifolia)、梭果玉蕊 (Barringtonia fusicarpa)组成;下层乔木(6~16 m)所受光线强度最弱,主要为假海桐(Pittosporopsis kerrii)、木奶果(Baccaurea ramiflora)、版纳柿(Diospyros xishuangbannaensis)、棒柄花(Cleidion brevipetiolatum)。
2 材料与方法
2.1 研究方法
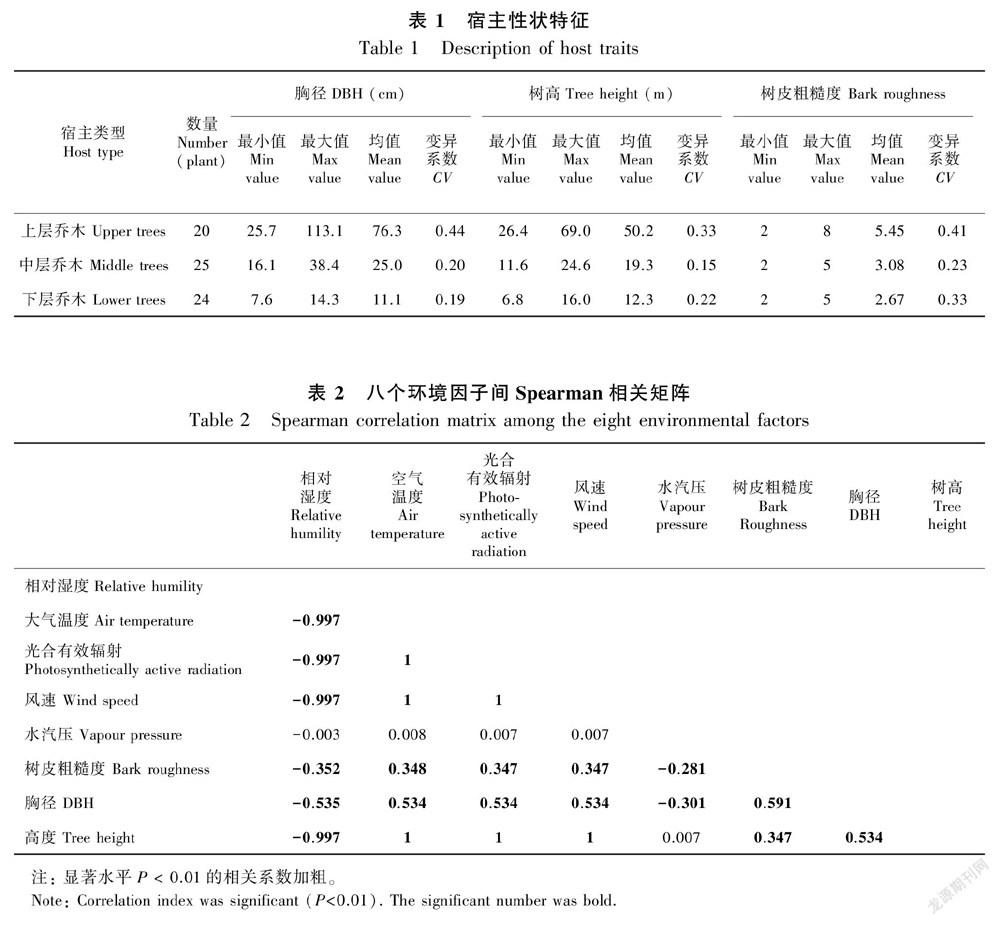
附生苔藓植物的野外调查于2017年4—5月展开。共调查了样地内69棵树,包括20棵上层乔木、25棵中层乔木、24棵来自下层乔木,宿主特征如表1。本研究主要利用龙脑香热带雨林样地内的林冠塔吊且借助特制的伸缩工具采集高度大于10 m的中、上层乔木上的苔藓,通过长梯辅助采集10 m以下的树干上的苔藓。每个样地内,随机选取每个乔木层里没有严重损伤的树种。被选取的每棵下层(U1-U4)和中层(S1-S4)乔木,从树基到林冠顶端被划分为4个垂直区域:U1(S1),为树干基部即树干2 m以下;U2(S2),树干下部;U3(S3),树干上部;U4(S4),为整个树冠即从第一处主分叉到林冠顶部(Sporn et al., 2010)。上层乔木由于树冠分枝多、结构复杂,将其分为6个垂直区域:C1,为树干基部即树干2 m以下;C2,树干下部;C3,树干上部;C4,内部林冠;C5,中部林冠;C6,外部林冠 (Johansson, 1974)。对于树基和树干区域,用20 cm×20 cm的取样框分别在每个垂直区域的迎风面(西南)和背风面(东北)方向取样(González-Mancebo et al., 2004; Holz & Gradstein, 2005),上下各取2份,每个树干上的垂直区域共取4份(每个垂直区域采样面积为1 600 cm2)。林冠上的每个区域根据树枝径级用不同大小的取样框采样4~6份,尽可能选取每个方位上面和侧面的枝干(每个区域采样总面积都为1 600 cm2)。
宿主胸径数据由西双版纳森林生态系统国家野外科学观测研究站提供。用长为100 m的皮尺测得树高(HT)和林冠高度(HC)数据。每个树上垂直区域的高度是指从地面到每个垂直区域中间位置的距离,分别由以下公式计算(H树基=2, H树干=HT-HC -2, H树干下部=1/4(HT-HC-2)+2, H树干上部=3/4(HT-HC-2)+2, H树冠内部=(HT-HC)×1/3×1/2 , H树冠中部=(HT-HC)×1/2, H树冠外部=(HT-HC)×5/6)。为了比较和量化树的区域高度,我们将垂直区域的高度值转化为13个垂直高度( H1=1~5 m, H2=6~10 m, H3=11~15 m, H4=16~20 m, H5=21~25 m, H6=26~30 m, H7=31~35 m, H8=36~40 m, H9=41~45 m, H10=46~50 m, H11=51~55 m, H12=56~60 m, H13=61~65 m)。树皮的粗糙度等级,我们采用Male & Roberts (2005)的1~9个等级划分方式(如图4,1=非常光滑; 3=光滑但有裂纹; 5=浅褶皱; 7=裂纹较深; 9=表面充斥着大量起伏不平的裂痕; 然而 2, 4, 6, 8 粗糙度是介于相邻两个等级之间的)。样地内,空气温度(℃, 型号HMP155A)、相对湿度(%RH, 型號HMP155A)、光合有效辐射(μmol· m-2·s-1, 型号 SQ-311)、风速(m·s-1, 型号CSAT3)和空气压力(kPa, 型号CS106) 数据被位于塔吊垂直梯度上的8套小气候系统实时监控并每小时记录一次。由于实验期间部分数据存在异常,我们仅使用了2017年6月、7月分别位于补蚌塔吊1.4、23.6、47.2、55.2、62.1 m高度上的小气候数据。其中,由于水汽压对于解释苔藓群落的分布更有意义,因而空气压力通过戈夫-格雷奇公式转化为水汽压。
该区域苔藓的生活型可划分为7种,即扇型(fan)、细平铺型(smooth mat)、粗平铺型(rough mat)、悬垂型(pendant)、矮丛集型(short turf)、高丛集型(tall turf)和交织型(weft) (Mgdefrau, 1982)。本研究苔藓除少数难以鉴定的标本由专家帮鉴定外,其余样品均由作者自行鉴定。凭证标本存放于中国科学院西双版纳热带植物园生物多样性研究组实验室。
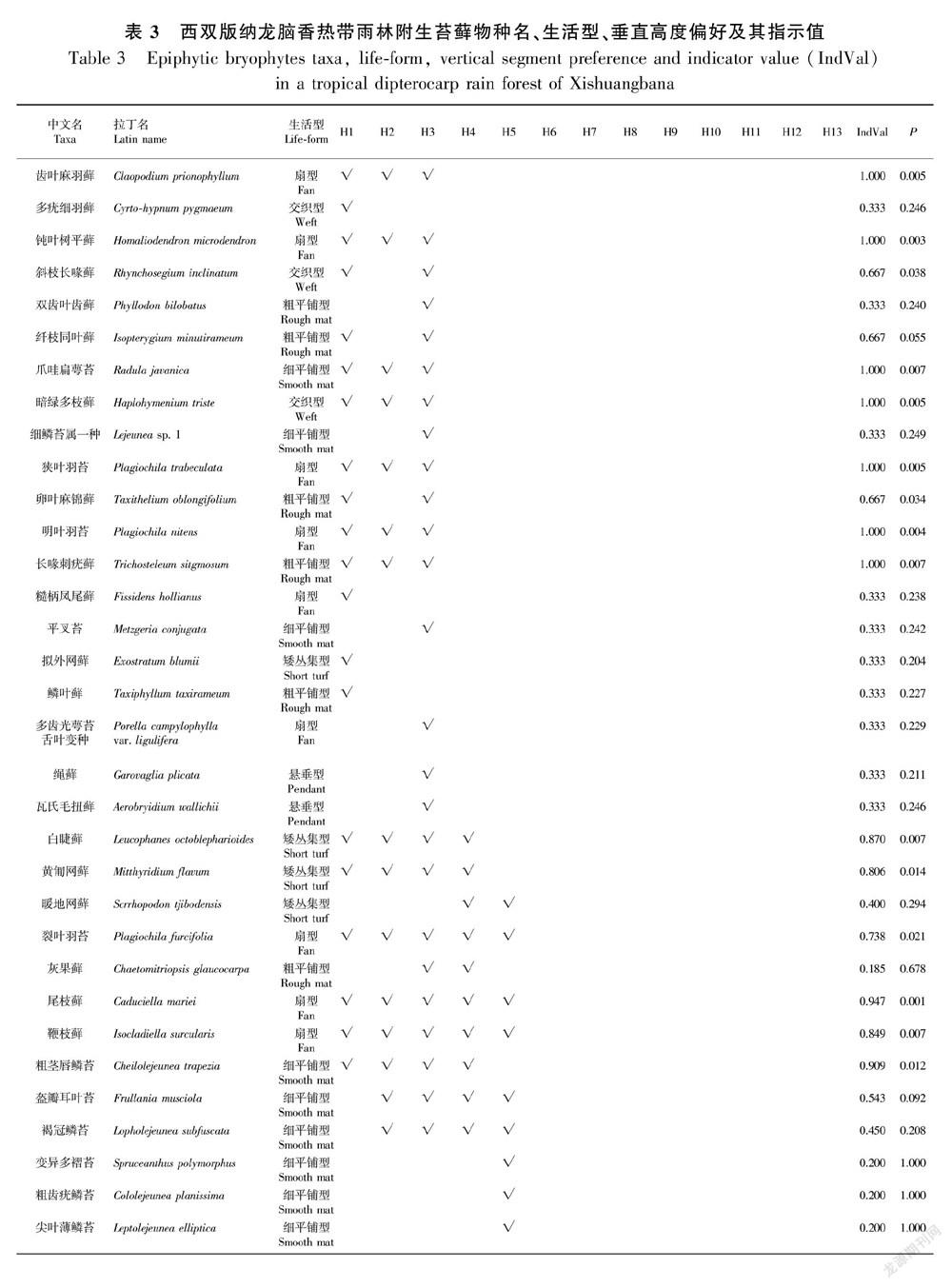
2.2 数据处理
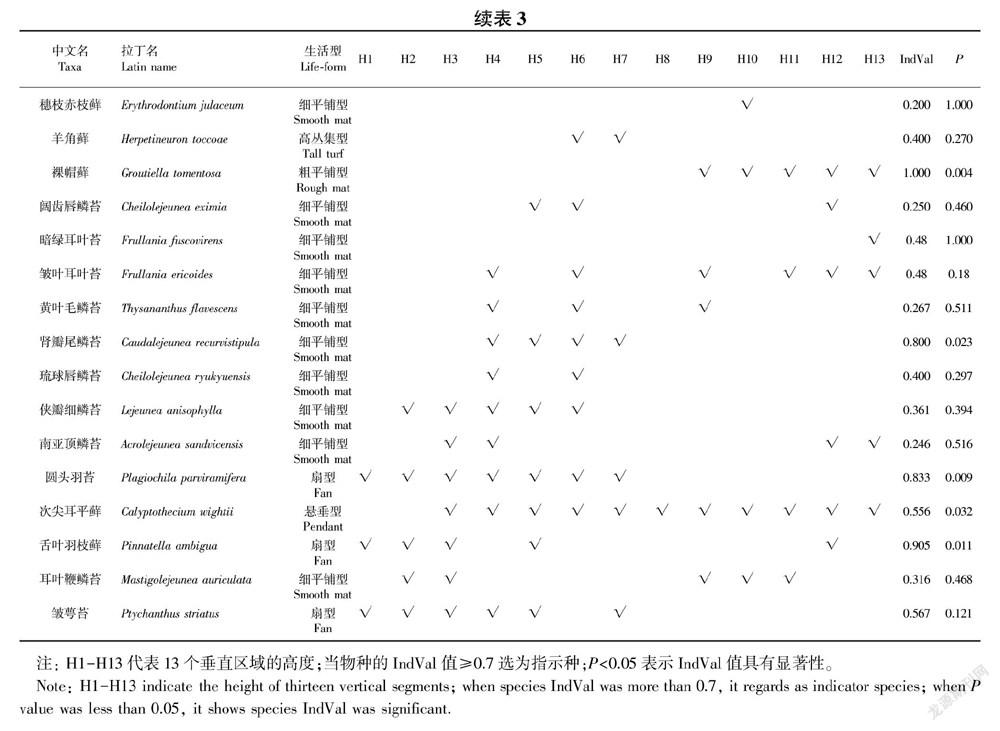
本研究涉及到的统计分析均由开源统计平台3.4.3版本的R (http://www.r-project.org)以及Origin pro 9.0完成。统计各个垂直区域内,每个物种出现的频数。为使结果更可靠,排除了低频的偶见种(排除41种),即13个垂直高度中每个垂直高度的频数仅出现1次或2次的苔藓物种未参与统计。数据集由垂直区域-苔藓数据(每行代表一个垂直区域,每列代表一种苔藓或者生活型)和垂直区域-环境因子数据(每行代表一个垂直区域,每列代表一个环境因子)组成,其中环境因子包括气候因子(空气温度、相对湿度、光合有效辐射、风速和水汽压)和宿主特征因子(胸径、区域高度和树皮粗糙度)。根据spearman做环境因子间的相关性比较分析(表2),大气湿度与空气温度、光合有效辐射、风速、树高都显著相关(相关系数大于0.6,P<0.05)。大气湿度与苔藓生理关系最为密切,所以将其保留,其余相关的因子均舍去。因此,大气湿度、水汽压、胸径与树皮粗糙度选为排序分析的环境因子。利用vegan包 (Oksanen etal., 2017)里的典范对应分析(CCA)方法来分析附生苔藓及其生活型与潜在驱动因子之间的关系。其中,唯一一种分布于H6、H7的高丛集型苔藓,由于该类生活型频度过低未能显示于CCA图中。根据CCA图苔藓群落在垂直区域上的偏好类型(H1-H3区域,H4-H8区域和H9-H13区域),我们利用R语言中的Labdsv软件包(Roberts, 2016)中的IndVal函数计算各个附生苔藓的IndVal值(Dufrêne & Legendre, 1997) 以筛选出指示特殊生境的指示种,并且以IndVal值大于0.7作为标准确定指示物种(李巧,2001)。
3 结果与分析
3.1 附生苔藓的科、属、种的分布样地内共记录到69棵树13个垂直高度上316个垂直区域的附生苔藓植物90种,隶属于60属25科。其中,苔类37种19属6科,藓类53种41属19科。细鳞苔科(Lejeuneaceae)是优势大科,该科所包含的苔藓物种数占所有苔藓总数的25.6%,其次是蔓藓科(Meteoriaceae)、耳叶苔科(Frullaniaceae),其物种数各占总苔藓数的8.9%和6.7%。
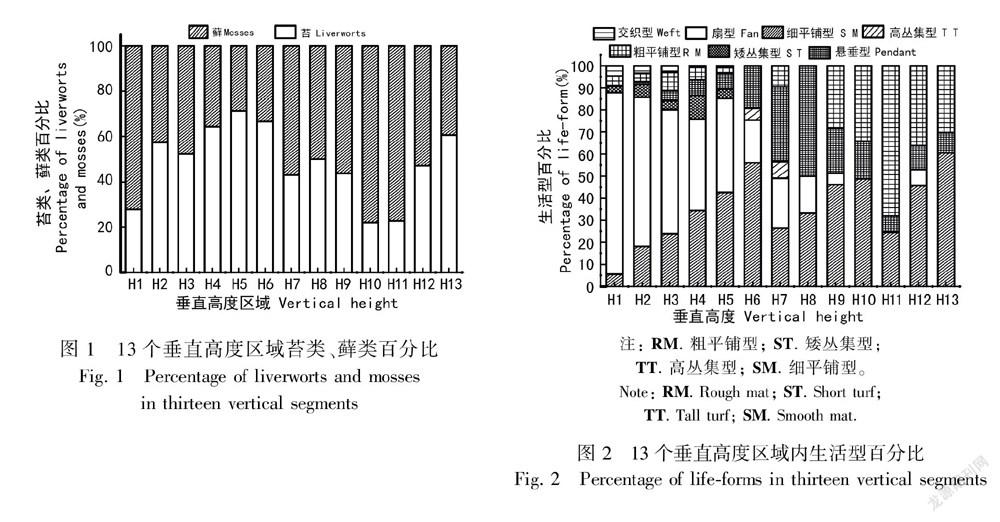
3.2 附生苔类、藓类组成和苔藓的生活型组成
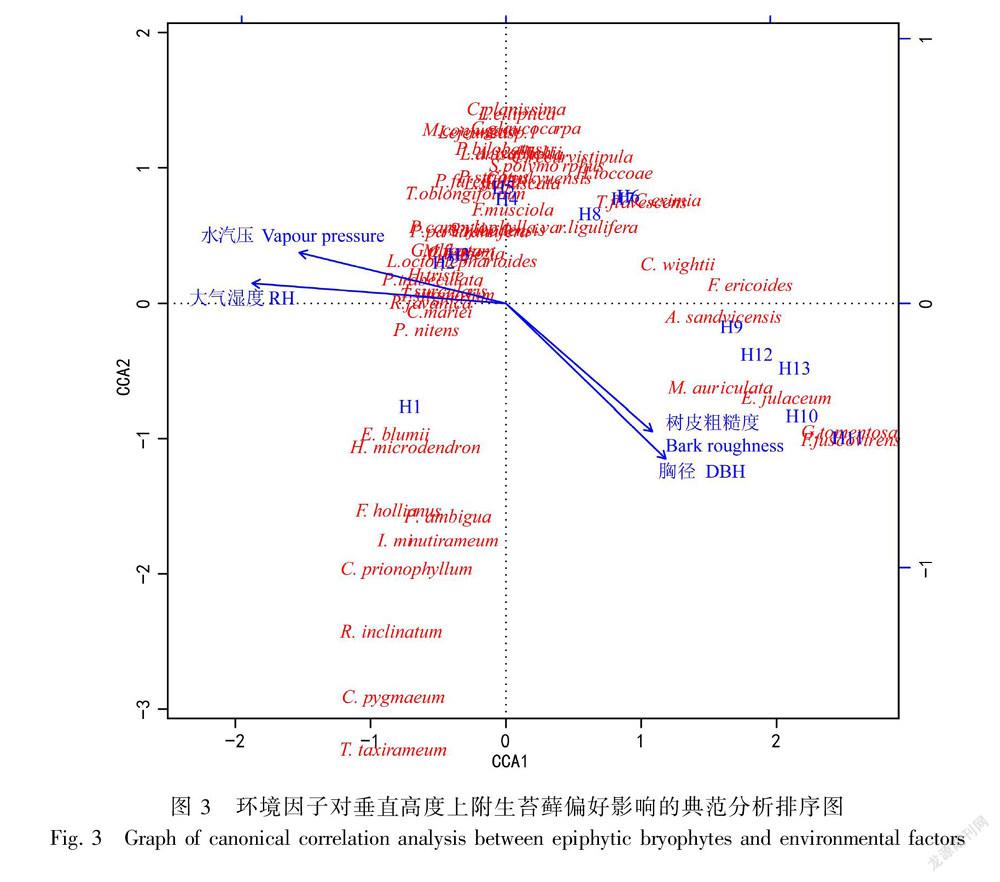
3.3 附生苔藓对不同宿主垂直高度的偏好从环境因子与苔藓物种的CCA分析(图3)可以看出,附生苔藓群落可分为三个聚集的群体:H1-H3区域苔藓群落,H4-H8区域苔藓群落,H9-H13区域苔藓群落。树干H1-H3所在区域与大气湿度高度相关,主要由斜枝长喙藓(Rhynchosegium inclinatum)、齿叶麻羽藓(Claopodium prionophyllum)、纤枝同叶藓(Isopterygium minutirameum)、舌叶羽枝藓(Pinnatella ambigua)、糙柄凤尾藓(Fissidens hollianus)、钝叶树平藓(Homaliodendron microdendron)、拟外网藓(Exostratum blumii)等20种附生苔藓组成(表3),高度大于45 m(>H9)垂直区域与大气湿度呈负相关,该区域主要由耳叶苔科的皱叶耳叶苔 (Frullania ericoides)和暗绿耳叶苔(F. fuscovirens),细鳞苔科的南亚顶鳞苔(Acrolejeunea sandvicensis)和耳叶鞭鳞苔(Mastigolejeunea auriculata),木灵藓科的裸帽藓(Groutiella tomentosa)和穗枝赤齿藓(Erythrodontium julaceum)6个苔藓物种组成(表3);而处于H4-H8区域的24种附生苔藓群落则对大气湿度没有明显偏好。由表3可得,17种附生苔藓可以作为不同垂直区域的指示苔藓。其中,15种的指示物种在H1-H3区域发现,它们分别是齿叶麻羽藓、钝叶树平藓(Homaliodendron microdendron)、狭叶羽苔、明叶羽苔(Plagiochila nitens)、暗綠多枝藓(Haplohymenium triste)、爪哇扁萼苔(Radula javanica)、尾枝藓(Caduciella mariei)、 长喙刺疣藓(Trichosteleum sitgmosum)、舌叶羽枝藓、鞭枝藓(Isocladiella surcularis)、白睫藓(Leucophanes octoblepharioides)、粗茎唇鳞苔(Cheilolejeunea trapezia)、圆头羽苔(Plagiochila parviramifera)、黄匍网藓(Mitthyridium flavum)、裂叶羽苔(Plagiochila furcifolia);肾瓣尾藓苔(Caudalejeunea recurvistipula)和裸帽藓分别为H4-H7和H9-H13区域唯一的指示物种。
3.4 影响附生苔藓沿垂直高度上偏好的主要驱动因子
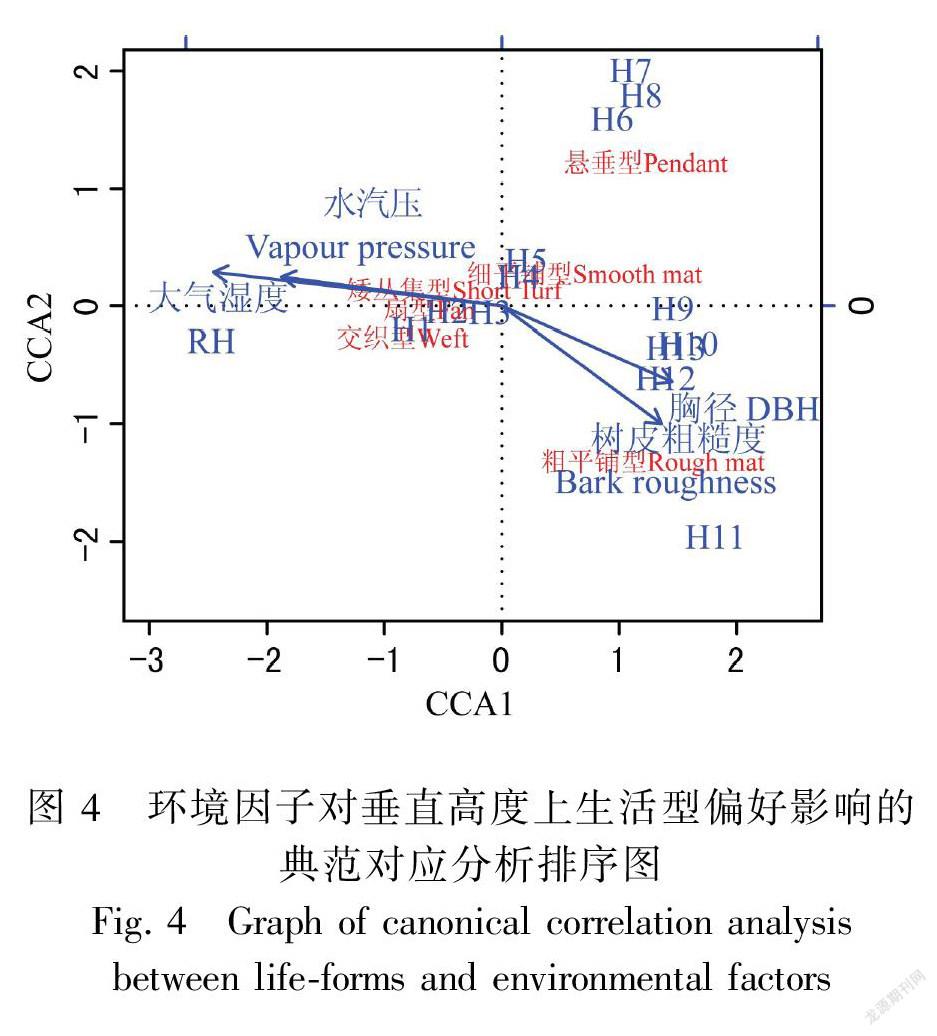
典范对应分析(CCA)方法的结果表明,4个环境因子对于7种苔藓生活型的分布格局共解释了16.98%。蒙特卡罗置换检验结果表明4个环境因子对于苔藓生活型的分布格局的影响具有显著性(P=0.011),因而CCA的结果具有可信性。大气湿度、水汽压、胸径及树皮粗糙度对苔藓生活型的分布均具有显著影响(P=0.001)。由CCA排序结果(图4)可知,交织型、扇型和矮丛集型与大气湿度、水汽压呈高度正相关,且该类生活型主要分布在H1-H3区。粗平铺型苔藓与胸径、树皮粗糙度呈正相关,主要集中分布在高度大于H9的区域。悬垂型的苔藓主要分布在H6-H8区域之间,而细平铺型的苔藓却广泛分布于高于H4的区域。
4 讨论与结论
与Gradstein & Pócs(1989)早期的研究相似,我们所调查到的细鳞苔科、耳叶苔科、羽苔科、扁萼苔科、蔓藓科、蕨藓科、平藓科、木灵藓科、凤尾藓科、花叶藓科、灰藓科、锦藓科占据了绝大多数热带雨林典型的科。细鳞苔科占有绝对的优势,是该热带森林样地内含物种数最多的科,这与其他热带区域的调查结果一致(Cornelissen & Streege, 1989; Sporn et al., 2010; Oliveira et al., 2016)。
沿垂直高度升高,苔类所占比例呈现出先升高后降低又升高的趋势。这表明苔类在树干上部和林冠外部分布较多,即苔类可能更倾向于分布于光照更强的区域。一些苔类具小裂片的腹瓣用于储藏水分(Kelly et al., 2004),紧密覆瓦状排列的侧叶在强光下也可减少水分的蒸发。而一些分布于林冠树枝上喜阳的藓类如木灵藓科物种,在高温、强光的环境下,倾向于呈覆瓦状排列,以减缓水分散失,还能保护茎部不受伤害(Scott, 1982)。分布高度大于H9区域的苔藓植物与湿度呈负相关,而光合有效辐射和湿度高度相关,即该类苔藓植物与PAR显著地呈正相关。分布于该区域的绝大多数苔藓的叶片呈现出红色或红褐色,这些有色色素可通过光化学途径来保护植物体(Krinsky, 1968),如主要分布于上层树冠上(>H9)的耳叶苔科的皱叶耳叶苔(Frullania ericoides)呈现出红棕色的,而盔瓣耳叶苔(F. musciola)呈现出红褐色。此外,有色苔藓所在的垂直高度越高所呈现出的颜色越深(Cornelissen & Streege, 1989),如分布在H12和H13垂直区域的皱叶耳叶苔的红棕色比分布在H6或H4垂直区域的该种苔藓颜色更暗。
在热带雨林,隐花附生植物作为生态指示种是极其重要的(Gradstein et al., 2001)。Holz & Gradstein(2005)指出,它们在森林中的分布格局反映了微生境中光温湿和基质条件的差异。这与我们的研究结果一致,齿叶麻羽藓、钝叶树平藓、狭叶羽苔等附生苔藓多出现在树基和较低的树干区域,对阴暗潮湿的林下生境具有指示作用,而裸帽藓偏好于散生巨树的树冠上,是高光干燥的林冠生境的最优指示种。之前的研究(Holz & Gradstein,2005)将苔藓指示种用于指示三种不同的森林类型,我们首次尝试将物种指示法引入附生植物对宿主垂直高度的生境偏好中,这更符合统计学意义,同时也反映了附生苔藓指示种对特殊微生境的指示作用。
基于以上附生苔蘚在宿主垂直高度上的生态适应性特征,我们将附生苔藓群落划分为三种生态型:喜阳苔藓、喜阴苔藓和广布苔藓。喜阳苔藓(主要分布于高于H9区域,即散生巨树>45 m上的区域)在上层林冠上占优势,适应于较强的阳光;喜阴苔藓(限制于高度低于H3区域,即<15 m的下层乔木树上)分布于较低的树干区域和下层乔木,偏爱湿润、凉爽的生境;广布苔藓(通常可分布于0~70 m间的区域,即三个以上的间断高度区域)则在垂直树干上具有较宽的生态位。这与之前附生苔藓在Johansson区域出现频度划分的结果相似(Cornelissen & Streege, 1989; Acebey et al., 2003),本研究用量化的高度数据进一步为龙脑香热带林上的附生苔藓植物生态型的划分提供了更为直接的证据。
随着宿主垂直高度的升高,交织型和扇型苔藓频度降低,而细平铺型和粗平铺型增多,这反映了垂直梯度上光照和水分条件的梯度性变化。植株矮小、紧密平铺的细平铺型苔藓的形态结构有利于减少水分的散失,以应对干旱、强光的微环境。因而,该类型的苔藓主要分布在垂直高度大于20 m(>H4)的树干上,扇型和交织型则主要分布在湿润的较低区域(H1-H3)(Bates, 1998)。其中,高、矮丛集型的苔藓对宿主微生境具有不同偏好。矮丛集型苔藓主要是主要分布于H2、H3区域的树干上;而高丛集型主要分布于H6、H7区域的水平或分杈的树冠上(Sporn et al., 2010)。悬垂型苔藓主要分布于H7、H8区域树干上,这可能是由于该区域所在的中层林冠有风有雾水,更有利于直接吸收雨水和雾气中的水分(Glime,2013),同时中层乔木茂密的树叶阻挡了一部分阳光,可以避免强光照的直接损伤。此外,悬垂型苔藓的细胞壁上通常都有疣状突起结构,这有利于反射强烈的阳光以减少水分散失(杨武,2008)。粗平铺苔藓与胸径、树皮粗糙度呈正相关,这可能是由于该类型的苔藓群落倾向于在表面积较大、树皮较为粗糙的树干上形成紧密的垫状,这样可有效地减少水分的蒸发和提高其持水能力以适应干热的微生境(吴鹏程,1998)。苔藓生活型组成沿宿主垂直梯度的变化与之前Sporn et al.(2010)在印度尼西亚热带森林内的调查结果是一致的,体现了苔藓植物对水热条件的响应与适应策略。
综上所述,沿宿主垂直梯度微环境的变化,附生苔藓生活型和形态结构有着不同的响应方式(如喜阳物种和喜阴物种在生活型上分别以平铺型和扇型为主;而形态上喜阳物种比喜阴物种植株更小,颜色更深),而聚集在同一微生境的苔藓植物则有着相似的适应机制(如不同喜阳物种的保水力和抵抗强光的适应机制相似)。附生苔藓沿垂直梯度的分布格局及其对微生境的偏好可能是其对林冠异质生境长期适应与自身进化的结果。今后,在森林林冠生境变化的监测和管理中,对微生境具有明显偏好的附生苔藓物种可作为有效的指示材料。
致谢 感谢云南西双版纳森林生态系统国家野外科学观测研究站提供塔吊平台和小气候监测数据。
参考文献:
ACEBEY A, GRADSTEIN SR, KRMER T, 2003. Species richness and habitat diversification of bryophytes in submontane rain forest and fallows of Bolivia [J]. J Trop Ecol, 19:9-18.
AH-PENG C, CARDOSO AW, FLORES O, et al., 2017. The role of epiphytic bryophytes in interception, storage, and the regulated release of atmospheric moisture in a tropical montane cloud forest [J]. J Hydrol, 548:665-673.
ARIYANTI NS, BOS MM, KARTAWINATA K, et al., 2008. Bryophytes on tree trunks in natural forests, selectively logged forests and cacao agroforests in Central Sulawesi, Indonesia [J]. Biol Conserv, 141:2516-2527.
BATES JW, 1998. Is ‘life-form’ a useful concept in bryophyte ecology? [J]. Oikos, 82(2):223-237.
BUCK WR, THIERS BM, 1989. Review of bryological studies in the tropics [M]//CAMBELL DC, HAMMOND HD. Floristic inventory of tropical countries. Bronx, New York:New York Botanical Garden:484-493.
CAO M, ZOU X, WARREN M, et al., 2006. Tropical forests of Xishuangbanna, China [J]. Biotropica, 38(3):306-309.
CHAZDON RL, FIELD N, 1984. Light environment of tropical forest [M]//MEDINA E, MOONEY HA, VZQUEZ-YNES C. Physiological Ecology of Plants of the Wet Tropics. The Hague:Junk Publishers.
CORNELISSEN JHC, STREEGE HT, 1989. Distribution and ecology of epiphytic bryophytes and lichens in dry evergreen forest of Guyana [J]. J Trop Ecol, 5(2):131-150.
DISLICH R, MANTOVANI W, 2016. Vascular epiphyte assemblages in a brazilian atlantic forest fragment:Investigating the effect of host tree features [J]. Plant Ecol, 217(1):1-12.
DUFRNE M, LEGENDRE P, 1997. Species assemblages and indicator species:The need for a flexible asymmetrical approach [J]. Ecol Monogr, 67:345-366.
ENGEMANN KB, SANDEL BJ, ENQUIST PM, et al., 2016. Patterns and drivers of plant functional group dominance across the Western Hemisphere:A macroecological re-assessment based on a massive botanical dataset [J]. Bot J Linn Soc, 180:141-160.
FRAHM JP, GRADSTEIN SR, 1991. An altitudinal zonation of tropical rain forests using bryophytes [J]. J Biogeogr, 18(6):669-678.
FREY W, GOSSOW R, KRSCHNER H, 1990. Verteilungsmuster von Lebensformen,wasserleitenden und wasserspeichernden Strukturen in epiphytischen Moosgesellschaften am Mt. Kinabalu (Nord-Borneo) [J]. Nova Hedwigia, 51:87-119.
GLENN MG, COLE MS, WEBB SL, et al., 1998. Corticolous lichens and bryophytes:Preliminary surveys of old growth and managed northern hardwood stands in Minnesota [M]// GLENN MG, HARRIS RC, DIRIG R. Lichenographia Thomsoniana:North American Lichenology in Honor of John W. Thomson. New York :Mycotax 2 on Ltd:407-422.
GLIME JM, 2013.Adaptive Strategies:Growth and Life Forms. Chapt. 4-5 [M]//GLIME JM. Bryophyte Ecology. Vo-lume 1. Ebook sponsored by Michigan Technological University and the International Association of Bryologists. http://digitalcommons.mtu.edu/bryophyte-ecology1/
GONZLEZ-MANCEBO JM, ROMAGUERA F, LOSADA-LIMA A, et al., 2004. Epiphytic bryophytes growing on Laurus azo-rica, (Seub.) Franco in three laurel forest areas in Tenerife (Canary Islands) [J]. Acta Oecol, 25(3):159-167.
GRADSTEIN SR, CHURCHILL SP, SALAZAR AN, 2001.Guide to the bryophytes of tropical America [J]. Febs Lett, 26(1-2):20-24.
GRADSTEIN SR, PCS T, 1989. Bryophytes [M]// LIETH H, WERGER MJA. Tropical Rain Forest Ecosystems. Amsterdam, the Netherlands:Elsevier Science Publishers:311-325.
HOLZ I, GRADSTEIN SR, 2005. Cryptogamic epiphytes in primary and recovering upper montane oak forests of Costa Rica:Species richness, community composition and ecology [J]. J Plant Ecol, 178(1):89-109.
HOLZ I, GRADSTEIN SR, HEINRICHS J, et al., 2002. Bryophyte diversity, microhabitat differentiation and distribution of life forms in Costa Rican upper montane Quercus forest [J]. Bryologist, 105:334-348.
JOHANSSON D, 1974. Ecology of vascular epiphytes in West African rain forest [J]. Acta Pharm Suec, 59:1-136.
KELLY DL, O′DONOVAN G, FEEHAN J, et al., 2004. The epiphyte communities of a montane rain forest in the andes of Venezuela:Patterns in the distribution of the flora [J]. J Trop Ecol, 20(6):643-666.
KRINSKY NI, 1968. Chapter 5-The protective function of caro-tenoid pigments [M]// GIESE AC. Photophysiology:Current Topics. Elsevier Inc:123-195
KRSCHNER H, PAROLLY G, 1998. Syntaxonomy of trunk-epiphytic bryophyte communities of tropical rain forests. A first pantropical approach [J]. Phytocoenologia, 28:357-425.
KRSCHNER H, 1990. Die epiphytischen Moosgesellschaften am Mt. Kinabalu (Nord-Borneo, Sabah, Malaysia) [J]. Nova Hedwigia, 51:1-75.
LI Q, 2011. Indicator value (IndVal) method and its application [J]. Chin Bull Entomol, 48(2):457-462. [李巧, 2011. 指示值方法及其在昆蟲中的应用 [J]. 应用昆虫学报,48(2):457-462.]
LOWMAN MD, SCHOWALTER TD, 2012. Plant science in forest canopies — The first 30 years of advances and challenges(1980-2010) [J]. New Phytol, 194:12-27.
MA KP, 2015. Biodiversity monitoring in China:From CForBio to Sino BON [J]. Biodivers Sci, 23(1):1-2. [马克平, 2015. 中国生物多样性监测网络建设:从CForBio到Sino BON [J]. 生物多样性, 23(1):1-2.]
MA KP, 2015. Biodiversity monitoring relies on the integration of human observation and automatic collection of data with advanced equipment and facilities [J]. Biodivers Sci, 24(11):1201-1202. [马克平, 2016. 生物多样性监测依赖于地面人工观测与先进技术手段的有机结合 [J]. 生物多样性,24(11):1201-1202.]
MGDEFRAU K, 1982. Life-forms of bryophytes [M]//SMITH AJE. Bryophyte Ecology [M]. Chapman and Hall, London:45-58.
MALE TD, ROBERTS GE, 2005. Host associations of the strangler fig Ficus watkinsiana in a subtropical Queensland rain forest [J]. Austral Ecol, 30:229-236.
MITCHELL AW, SECOY K, JACKSON T, 2002. The global canopy handbook:Techniques of access and study forest roof [M]. Oxford, UK:Global Canopy Programme.
MITTERMEIER RA, ROBLES GP, HOFFMANN M, et al., 2004. Hotspots revisited:Earth’s biologically richest and most endangered terrestrial ecoregions [M]. Mexico City:CEMEX.
NAKAMURA A, KITCHING RL, CAO M, et al., 2017. Forests and their canopies:Achievements and horizons in canopy science [J]. Trend Ecol Evol, 32(6):438-451.
OKSANEN J, BLANCHET FG, FRIENDLY M, et al., 2017. Vegan:Community Ecology Package. R package version 2.4-4. https://CRAN.R-project.org/package=vegan
OLIVEIRA HCD, OLIVEIRA SMD, OLIVEIRA HCD, et al., 2016. Vertical distribution of epiphytic bryophytes in Atlantic forest fragments in northeastern Brazil [J]. Acta Bot Brasi:30(ahead).
RAES N, CANNON CH, HIJMANS RJ, et al., 2014. Historical distribution of sundalands dipterocarp rainforests at quaternary glacial maxima [J]. Proc Natl Acad Sci USA, 111(47):16790-16795.
ROBERTS DW, 2016. Labdsv:Ordination and multivariate analysis for ecology. R package version 1.8-0. https://cran.r-project.org/web/packages/labdsv
SCOTT GAM, 1982. Desert bryophytes [M]// SMITH AJE. Bryophyte Ecology. London:Chapman and Hall:105-122.
SHI XM, SONG L, LIU WY, et al., 2017. Epiphytic bryophytes as bio-indicators of atmospheric nitrogen deposition in a subtropical montane cloud forest:Response patterns, mechanism, and critical load [J]. Environ Poll, 229(X):932-941.
SLACK NG, 1976. Host specificity of bryophytic epiphytes in eastern North America (Geography and Ecology of Bryophytes) [J]. J Hatt Bot Lab, 41:107-132.
SMITH AJE, 1982. Bryophytes and Epiliths in Bryophyte Ecology [M]// SMITH AJE. Bryophyte Ecology. London:Chapman and Hall:1-14.
SONG L, LIU WY, MA WZ, et al., 2011. Bole epiphytic bryophytes on Lithocarpus xylocarpus, (Kurz) Markgr. in the Ailao Mountains, SW China [J]. Ecol Res, 26(2):351-363.
SPORN SG, BOS MM, KESSLER M, et al., 2010. Vertical distribution of epiphytic bryophytes in an Indonesian rainforest [J]. Biodivers Conserv, 19(3):745-760.
SPORN SG, BOS MM, HOFFSTTTER-MNCHEBERG M, et al., 2009. Microclimate determines community composition but not richness of epiphytic understorey bryophytes of rainforest and cacao agroforests in Indonesia [J]. Funct Plant Biol, 36:171-179.
SONG L, LIU WY, MA WZ, et al., 2011. Bole epiphytic bryophytes on Lithocarpus xylocarpus, (Kurz) Markgr. in the Ailao Mountains, SW China [J]. Ecol Res, 26(2):351-363.
STORK NE, 2007. Dynamics and processes in the canopy of an Australian tropical rainforest [J]. Austral Ecol, 32:2-3.
SZARZYNSKI J, ANHUF D, 2001. Micrometeorological conditions and canopy energy exchanges of a neotropical rain forest (surumoni-crane project, venezuela) [J]. Plant Ecol, 153(1/2):231-239.
TANG JW, 2008. A study on the population ecology of Parashorea chinensis in Xishuangbanna, SW China [D]. Mengla:Xishuangbanna Tropical Botanical Garden, Chinese Academy of Sciences. [唐建維, 2008. 西双版纳望天树种群生态学研究 [D]. 勐腊:中国科学院西双版纳热带植物园.]
WOLF JH, 1993. Diversity patterns and biomass of epiphytic bryophytes and lichens along an altitudinal gradient in the northern Andes [J]. Ann Mo Bot Gard, 80:928-960.
WOODS CL, CARDELUS CL, DEWALT SJ, 2015. Microhabitat associations of vascular epiphytes in a wet tropical forest canopy [J]. J Ecol, 103(2):421-430.
WU PC, 1998. Bryological biology [M]. Beijing:Science Press:318-328. [吳鹏程, 1998.苔藓植物生物学 [M]. 北京:科学出版社:318-328.]
WU Y, LIU WY, SONG L, et al., 2016. Advances in ecological studies of epiphytes using canopy cranes [J]. Plant Ecol,40(5):508-522. [吴毅,刘文耀,宋亮,等,2016. 基于林冠塔吊的附生植物生态学研究进展 [J]. 植物生态学,40(5):508-522.]
WU YH, GAO Q, CHENG GD, et al., 2002. Response of bryophytes to global change and its bioindicatortation [J]. Chin J Appl Ecol, 13(7):895. [吴玉环,高谦,程国栋,等, 2002. 苔藓植物对全球变化的响应及其生物指示意义 [J]. 应用生态学报,13(7):895-900.]
YANG W, 2008. On morphological and physiological adaptations of mosses to environments [D]. Jinhua:Zhejiang Normal University:1-2. [杨武, 2008. 藓类植物适应环境的形态结构及生理学机制 [D]. 金华:浙江师范大学:1-2.]
ZHU H, 2000. Ecology and biogeography of the tropical dipterocarp rain forest in Xishuangbanna [M]. Kunming:Yunnan Science & Technology Press:90-91. [朱华, 2000. 西双版纳龙脑香热带雨林生态学与生物地理学研究 [M]. 昆明:云南科技出版社:90-91.]
ZHU H, CAO HB, 2010. Geological history, flora, and vegetation of Xishuangbanna, southern Yunnan, China [J]. Biotropica, 38(3):310-317.
