尼罗罗非鱼PPARδ的原核表达及其多抗制备和纯化
2019-09-10潘传燕冯鹏霏张永德罗洪林
潘传燕 冯鹏霏 张永德 罗洪林



摘要:[目的]为研究尼罗罗非鱼过氧化物酶体增殖物激活受体δ(PPARδ)的表达情况,通过大肠杆菌表达系统表达PPARδ,纯化重组蛋白并获得多克隆抗体。[方法]对PPARδ进行生物信息学分析后设计特异性引物,扩增PPARδ,将其克隆至原核表达载体pET-B2m中,构建重组表达载体;重组质粒转人大肠杆菌B21并用IPTG进行诱导表达,用SDS-PAGE鉴定表达产物。用镍柱纯化重组蛋白后免疫日本大耳兔制备多克隆抗体,用间接ELISA技术检测抗体效价,WesternBlot鉴定抗体的特异性。[结果]成功构建了原核表达载体pET-B2m-PPAR8,实现了重组蛋白的原核表达和纯化,重组蛋白以包涵体蛋白和可溶性蛋白2种形式存在,分子量约为90kD;制备的多克隆抗体效价为1:2048000,WesternBlot检测结果表明该抗体能特异性识别尼罗罗非鱼PPARδ。[结论]成功实现了尼罗罗非鱼PPARδ重组蛋白的融合表达,在大肠杆菌中高效表达后纯化重组蛋白,免疫日本大耳兔获得了效价高、特异性强的多克隆抗体。为尼罗罗非鱼PPARδ的功能研究奠定了基础。
关键词:尼罗罗非鱼;PPARδ;原核表达;多克隆抗体
中图分类号:S917.4文献标志码:A 文章编号:1008-0384(2019)09-1053-06
0引言
(研究意义)罗非鱼具有生长快、食性杂、耐低氧、个体大、产量高和肥满度高等多种优良的养殖性状,是世界各国养殖户和消费者喜爱的品种之一,也是我国重要的养殖品种之一.近年来,脂肪含量作为影响鱼肉品质的一项重要因素受到了越来越多的关注,而过氧化物酶体增殖物激活受体(peroxisome proliferator-activated receptor,PPARs)在调控脂肪细胞分化、脂质代谢中起着重要作用,研究鱼类PPARs的功能及其作用机制,有利于提高鱼类肉质。因此,开展尼罗罗非鱼PPARs表达调控及其分子机制等相关研究具有重要意义。(前人研究进展)过氧化物酶体增殖物激活受体(PPARs)是核激素受体家族中的配体激活受体,是英国科学家首先发现的,具有调控脂质代谢、脂肪细胞增殖分化、胰岛素敏感、炎症反应等功能。有PPARα、PPARδ和PPARγ等3种亚型,在灵长类、啮齿类、两栖类、鸟类和部分水产动物中,这3种亚型已经被成功克隆和鉴定。其中,PPARα基因通过调控脂肪酸氧化相关基因的表达,在脂肪细胞的分化、脂质转运及脂肪酸氧化等方面发挥重要作用;PPARγ是最具脂肪组织特异性的PPARs成员,PPARγ的生物学功能主要有调节脂质代谢、抑制细胞凋亡、缓解炎症反应等;PPARδ参与皮肤和脑的发育、骨的形成、肥大细胞免疫以及肿瘤的发生等多种生物学过程,也参与许多疾病的发生和发展过程。PPARδ基因总长约35kb,有6个功能结构域,N端是配体非依赖的转录激活结构域,该区域保守性差;中间区域高度保守,为DNA结合域(DBD),含一个α螺旋的DNA结合基序和2个锌指结构;C端是配体结合域(LBD),比PPARa和PPARγ的配体结合域小,是一个可以与多种酸性亲脂化合物结合的袋装结构,PPARδ因其配体的特异性而具有特异的生理功能。PPARδ在动物各组织中均表达,在脂质代谢活跃的组织中表达水平较高,如脂肪组织、心脏、骨骼肌等部位,在脾、肺、内皮细胞、T淋巴细胞等也有表达。目前关于鱼类PPARs的研究较多,已在黄颡鱼、卵形鲳鰺、大黄鱼和虹鳟等多种鱼类中开展PPARs基因克隆和功能研究,但鱼类PPARδ的研究较少且主要集中在脂质代谢上。(本研究切入点)目前尚未见尼罗罗非鱼PPARδ的相关报道,尼罗罗非鱼PPARδ的功能和作用机制尚不清楚。(拟解决的关键问题)本研究通过构建尼罗罗非鱼PPARδ原核表达载体,利用大肠杆菌原核表达系统表达PPARδ重组蛋白,分析重组蛋白的表达情况,并将纯化后的重组蛋白免疫日本大耳兔,制备多克隆抗体,为进一步研究尼罗罗非鱼PPARδ的功能及其作用机制奠定基础。
1材料与方法
1.1 材料
参照潘传燕等前期研究中的材料制备方法。
1.2试验方法
试验方法主要参照潘传燕等的方法。
1.2.1PPARδ蛋白结构分析和抗原预测 PPARδ蛋白质疏水性用ExPASy中的ProtScale模块(https://web.expasy.org/protscalc/)进行分析,蛋白序列的跨膜区间用TMHMM server v.2.0(http://www.cbs.dtu.dk/services/TMHMM-2.0/)进行预测。使用DNAstar软件的Protean模块预测蛋白的亲水性、抗原指数等参数。
1.2.2PPARδ基因扩增及原核表达质粒构建 根据尼罗罗非鱼PPARδ基因序列(NCBI No.NP 001276-565.1)设计特异性引物,引物设计软件为PrimerPremier 5.0,引物由上海生工生物工程技术服务有限公司合成。引物序列为:PPARS-F:5'-TCCACTGGGT-TCTCGGACTATGGATGGTTTTCAGCA,PPARS-R:5'-TAAGGCCGCACTCGAGCACCACATACATATC-TTTATAGAT-3’。PCR扩增条件如下:94℃4min;94℃45s,52℃45s,72℃45s,28个循环;72℃10min。琼脂糖凝胶电泳检测扩增产物,之后进行切胶回收。
用SalΙ与BamHΙ对PCR产物进行酶切处理后,将其克隆至原核表达载体pET-B2m,构建重组表达质粒pET-B2m-PPARδ,转至大肠杆菌B21中培养,挑选阳性克隆,由上海生工生物工程技术服务有限公司完成测序鉴定。
1.2.3重组蛋白的表达和纯化 将重组质粒(pET-B2m-PPARδ)转化至大肠杆菌B21中,在LB培养基上进行扩大培养,培养基已提前加入100g.mL的氨苄青霉素(Amp),培养温度为37℃,当OD为0.6后,將培养温度降为30℃;此时加入异丙基-B-D硫代半乳糖苷(IP7G),IPTG的终浓度为0.5mmol·L,30℃条件下培养180min。然后收集菌液,离心收集菌体并用超声破碎,4℃20000g离心0.5h,收集上清液和沉淀,然后进行SDS-PAGE分析。
利用Ni-NTA树脂层析柱纯化重组蛋白,收集菌体,用10倍柱床体积的NTABuffer冲洗,流速控制在1mL·min左右,收集洗脱液。用紫外吸收法检测纯化后的蛋白浓度,并用SDS-PAGE电泳检测纯化后的重组蛋白。
1.2.4多克隆抗体制备和效价检测 PPARδ多克隆抗体的多肽序列如下:MDGFQQTAPEKHDGVNGYCEPNSPQDAADVRWTPPQAESVGSDSCGATSVSEVADLKELKRRESEDEEEKEEKEEKEGVPASKYPKRDQKKRRKEGEDQENSDNKQNSSASSSYTDLSHTSSPSLSEQLRLGREDSTSSGISVECKVCGDKASGFHYGVHACEGCKGFFRRTVRMKLEYERCERSCKIQKKNRNKCQYCRFQKCLSLGMSHDAIRYGRMPEAERKKLVAGILAEELNVSKPGGSDLKTLAKQVNTAYQKNLSMTKKRARSILMGKTSSTSPFVIYDVDTLWKAESGLVWSQLTPGAPLTKEIGVHVFYRCQCTTVETVRELTEFAKCIPGFVDLFLNDQVTLLKYGVHEAIFAmLPSLMNKDGLLVANGKGFVTREFLRSLRKPFSEIMEPKFEFAVKFNALELDDSDLALFVAAIILCGDRPGLMNVKQVEQSQDNILQALDLHLQANHSDSAYLFPKLLQKMADLRQLVTENAQLVQKIKKTESETSLHPLLQEIYKDMY。
试验用日本大耳兔2只(编号为G479、G480),耳静脉采集免疫前血清做阴性对照。将纯化后的重组蛋白与弗氏完全佐剂按1:1的容积比混合,皮下多点注射日本大耳兔,注射剂量为500ug·只。每只兔子最少免疫4次,每次免疫间隔2周,每次免疫1周后耳静脉采集免疫血清,用间接ELISA法检测抗体的效价。
重组PPARδ蛋白在96孔板中完成抗原包被,蛋白浓度为2ug·mL,加样量为100uL·孔,置于4℃过夜,以免疫前血清作阴性对照,待检样品按1:2000、1:4000作梯度稀释至1:32768000,将稀释液作为一抗加入到96孔板中;以HRP标记的羊抗兔IgG作为二抗。用TMB显色液进行显色反应,显色液用量为100pL·孔,显色时间为20min,加入50uL HSO终止显色反应,用酶标仪检测OD值,产生阳性反应的最大稀释度即为抗体的效价。
1.2.5抗体特异性的Western blotting分析 以免疫前血清为对照,取25、10ng纯化后的蛋白进行SDS-PAGE电泳,之后将蛋白转移至聚偏氟乙烯膜(PVDF膜)上,放置1h后用PBST缓冲液洗涤3次,每次5min,于封闭液中37℃封闭2h,封闭液是1%的酪蛋白。以制备的兔抗血清1:1000倍稀释液为一抗,在37℃孵育60min;洗涤后再用HRP标记的羊抗兔IgG作为二抗检测多克隆抗体的特异性。
1.2.6抗体纯化
采用Protein G亲和层析柱对PPAR5多抗进行纯化,操作步骤详见张永德等的方法,将抗血清与2×PBS缓冲液按1:1混合,经10倍柱体积以上的PBS洗滌,直至流出液无蛋白检出,加入2倍柱体积0.1mol'L柠檬酸(pH2.7)洗脱,收集洗脱产物,SDS-PAGE电泳检测纯化后的多抗。
2结果与分析
2.1 跨膜区和抗原性分析
PPARδ蛋白的跨膜区、表面可及性、亲疏水性等参数的分析结果(图1)显示,PPARδ蛋白编码512个氨基酸,1个跨膜区,270-287为跨膜区,该段区域疏水性较强;无信号肽序列,局部亲水性较好,全长蛋白抗原指数得分适中,进行全长表达,可以引起良好的免疫。
2.2 表达载体的构建
将双酶切载体和目的基因连接酶切所需片段,酶切位点为SaI和BamHI,转化至大肠杆菌B21培养后挑斑鉴定,经PCR扩增后,琼脂糖凝胶电泳检测扩增产物,目的条带大小为1536bp(图2),与预期大小一致。测序结果显示:目标序列100%正确,证实PPARδ基因已正确插入载体pET-B2m中,表达载体构建成功。
2.3融合蛋白的诱导表达和纯化
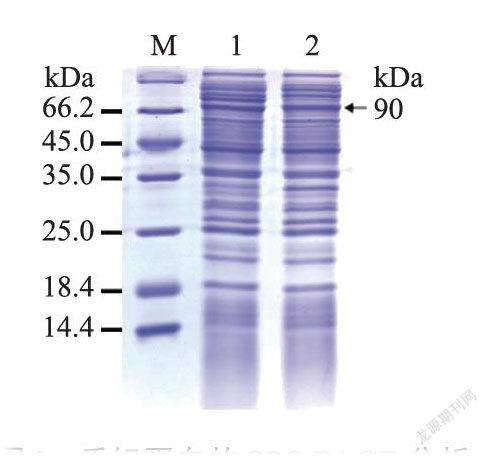
通过诱导剂对重组蛋白进行诱导表达,当诱导剂IPTG浓度为0.5mmol·L,30℃诱导3h,PPARδ得到较高水平的表达。结果显示,上清液与沉淀在90kD处都出现了清晰的蛋白条带(图3),上清液和沉淀的蛋白条带粗细相当,说明上清液和沉淀中的目的蛋白含量接近(图3),表明表达的融合蛋白有两种存在形式,即可溶性蛋白和包涵体蛋白。SDS-PAGE鉴定纯化后的重组蛋白,结果显示在90kD处有一条清晰的蛋白条带(图4),纯化后PPARδ蛋白量占总蛋白量的85%,质量浓度达1.5mg.mL,是适合免疫日本大耳兔的抗原。
2.4多克隆抗体的效价测定
以免疫前血清作为阴性对照,免疫一周后,耳静脉采血并分离血清,用间接ELISA法检测多抗的效价。结果表明,多克隆抗体的效价为1:2048000,抗体效价高。
2.5抗体特异性检测结果
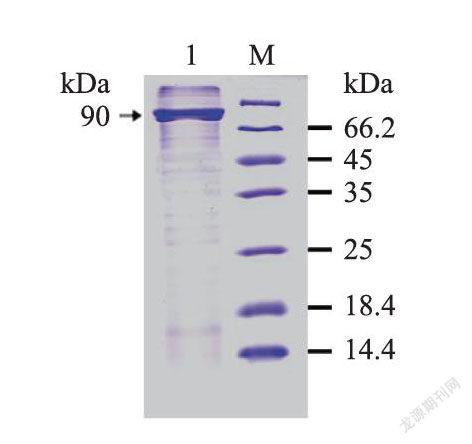
利用蛋白质印迹法(WB)检测抗体的特异性,结果表明:SDS-PAGE在约90kD处检测到清晰条带,且无杂带(图5),25ng重组蛋白的蛋白条带较10ng的蛋白条带粗,而阴性对照无特异性条带,多克隆抗体能特异性地结合尼罗罗非鱼PPARδ表达蛋白。
2.6抗体纯化结果
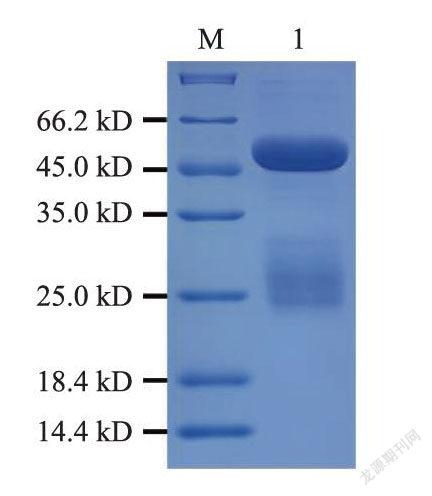
收集的抗血清经纯化后,纯度达85%以上,质量浓度为10mg.mL。SDS-PAGE分析结果显示,在约50、25ku处有清晰的条带(图6),表明抗体纯化效果良好。
3讨论
本研究通过大肠杆菌原核表达系统表达了尼罗罗非鱼PPARδ蛋白,该蛋白编码512个氨基酸,重组蛋白有2种表达形式,即包涵体蛋白和可溶性蛋白;纯化的重组蛋白免疫日本大耳兔后获得了效价高特异性好的多克隆抗体。钱云霞等表达了鲈PPAR?基因,其融合蛋白主要以包涵体蛋白存在,制备了效价达1:16000的多克隆抗体。关于鱼类PPARs的研究主要集中在基因克隆和组织表达,且PPARα和PPARγ的研究较多,而PPARδ的研究较少.Cho等克隆并分析了牙鲆PPARγ基因,cDNA全长1667bp,编码532个氨基酸,在饥饿条件下其表达增加。贾成霞等发现虹鳟PPARα在腹部肌肉、肠、肾脏等脂肪含量较高的组织中表达较高。高俊等克隆了刀鲚PPARγ基因并研究了PPARγ应激应答,刀鲚PPARγ的cDNA全长1951bp,ORF为1470bp,预测编码489个氨基酸。Zhao等指出,PPARα在团头鲂精卵巢中表达,精巢支持细胞和间质细胞中的PPARs可能直接参与生殖细胞的成熟。陈亮等克隆了草鱼PPARδ基因并比较了其在不同组织中的表达水平,结果显示,草鱼PPARδ基因cDNA核心序列片段大小为604bp,编码201个氨基酸,在肝脏、肌肉和心脏的表达水平较高。钱伦从大黄鱼肝脏中克隆了PPARδ全长eDNA,序列长3390bp,编码510氨基酸,在脾、肾、肝、鳃等多个组织均表达,在肝脏中表达量最高。尼罗罗非鱼PPARδ的原核表达和多克隆抗体制备尚未见相关报道。
多克隆抗体是由多个B淋巴细胞克隆所产生的,受到多种抗原决定簇刺激并可与多种抗原表位结合的抗体,存在于免疫动物的血清中。可通過直接分离血清获得。多克隆抗体制备过程较简单,用抗原免疫动物即可,经过3至4次免疫,ELISA检测抗血清效价,收集抗血清,纯化后即可获得多克隆抗体。效价高、特异性良好的抗体为基因表达和基因功能研究提供了物质基础。多克隆抗体具有制备快速、技术简单、稳定性良好、成本较低等优点,广泛应用于多个科研和生产领域。张曼等克隆并表达了锦鲤疱疹病毒ORFl基因,制备了多克隆抗体;李素一等制备了抗rFUR蛋白大鼠多克隆抗体。本研究获得的多克隆抗体效价达1:2048000,钱云霞等制备的鲈PPARγ多克隆抗体效价仅为1:16000。尼罗罗非鱼PPARδ多克隆抗体能特异性的识别尼罗罗非鱼PPARδ蛋白,说明该抗体具有良好的免疫学活性,为研究尼罗罗非鱼PPARδ的生物学特性和功能提供了分子工具。
生物信息学分析是开展基因组学和蛋白质组学研究的基础,它从核酸和蛋白质序列出发,分析序列中表达的结构功能的生物信息。本研究在制备多克隆抗体之前,对尼罗罗非鱼PPARδ进行了生物信息学分析。PPARδ基因,结构上高度保守,编码512个氨基酸,1跨膜区,无信号肽序列,局部亲水性好,抗原表位较好,可用于多抗制备。本研究制备的多抗效价高、特异性好,获得的重组蛋白和多克隆抗体为进一步探究尼罗罗非鱼PPARδ的功能和作用机制提供了分子工具。
