百子莲胚性愈伤组织在超低温过程中的蛋白质组学研究
2019-09-10吕珊张荻
吕珊 张荻


摘要:【目的】从蛋白质组学层面揭示植物胚性细胞对超低温保存复合逆境的响应调控机制,筛选出百子莲胚性愈伤组织在超低温保存中的差异表达蛋白,进一步丰富观赏植物超低温保存的分子生物学理论。【方法】利用iTRAQ技术鉴定百子莲胚性愈伤组织在超低温保存四个关键步骤(预培养、玻璃化脱水处理、洗涤和恢复培养)中的差异表达蛋白,应用生物信息学分析方法构建细胞响应超低温保存复合逆境的蛋白调控网络,筛选出植物超低温逆境保护类蛋白。【结果】共鉴定到2730种蛋白,筛选出821个差异表达蛋白。GO功能注释分析结果表明差异表达蛋白主要参与代谢、细胞过程和刺激应答等生物学过程,而催化活性和分子结合是最主要的分子功能类别。KEGG通路分析結果表明差异表达蛋白主要富集于能量代谢、蛋白的合成与降解、信号转导及细胞质膜修复等通路。通过STRING软件构建差异表达蛋白互作调控网络,发现上调表达蛋白主要参与糖代谢、氧化还原过程和核糖体翻译过程,下调表达蛋白主要参与ROS代谢、DNA修复、抑制细胞程序性死亡过程和细胞骨架。此外,筛选到96个与逆境保护相关的差异表达蛋白,分别参与能量代谢、氧化还原平衡、蛋白质合成与降解和信号转导。【结论】在超低温保存过程中,百子莲胚性愈伤组织的能量代谢、氧化应激与蛋白降解及PCD有所加剧,而细胞的ROS清除和胁迫防御过程明显下降,即百子莲胚性愈伤组织在超低温复合逆境下形成以糖代谢和抗氧化系统为主的抗逆保护机制。
关键词: 百子莲;胚性愈伤组织;蛋白质组学;超低温保存;差异表达蛋白
中图分类号: S682.19 文献标志码: A 文章编号:2095-1191(2019)11-2410-11
Proteomics analysis of embryogenic callus of Agapanthus praecox during cryopreservation
LYU Shan, ZHANG Di*
(School of Design, Shanghai Jiaotong University, Shanghai 200240, China)
Abstract:【Objective】The aim of the study was to reveal the response and protection mechanism of plant embryogenic cells under complex stress during cryopreservation on the proteomics level, and to find the differentially expressed proteins(DEPs) during cryopreservation of embryogenic callus of Agapanthus praecox to enrich the molecular biology theory. 【Method】iTRAQ(isobaric tags for relative and absolute quantification) technology was performed to identify DEPs in the four key steps of cryopreservation(preculture,vitrification dehydration,dilution,recovery) of the embryogenic callus of A. praecox,bioinformatics analysis method was used to construct the protein regulatory network in response to complex stress during cryopreservation and select stressed-protected proteins in plants. 【Result】A total of 2730 proteins were identified,of which 821 DEPs were selected. GO annotation analysis showed that these DEPs were mainly involved in biological process such as metabolic process,cellular process and responses to stimulus,while catalytic activity and molecular binding were the most important parts of molecular function. KEGG pathway analysis indicated that DEPs were mainly enriched in energy metabolism,protein synthesis and degradation,signal transduction and membrane repair. The DEPs interaction regulatory network was constructed by STRING software,it was found that up-regulated DEPs were mainly involved in carbohydrate metabolism,oxidation-reductive process and ribosome translation,while down-regulated DEPs were mainly involved in ROS metabolism,DNA repair,inhibition of programmed cell death(PCD) process and cytoskeleton. In addition,96 DEPs related to stress protection were detected for energy metabolism,redox homeostasis,protein synthesis and degradation and signal transduction. 【Conclusion】Energy metabolism,oxidative stress,protein degradation and PCD events of embryogenic callus of A. praecox increase during cryopreservation;while the ROS clearance and stress defense process of cells decrease greatly. Embryogenic callus of A. praecox form resistance protection mechanism with glycometabolism and antioxidant system as the majority under cryopreservation compound stress.
Key words: Agapanthus praecox; embryogenic callus; proteomics; cryopreservation; differentially expressed protein
0 引言
【研究意义】百子莲(Agapanthus praecox)为百子莲科百子莲属植物,源于非洲南部,为多年生草本花卉。作为优良种质资源,百子莲于2003年首次被引入我国,但由于夏季高温高湿气候条件的影响,其胚胎败育率较高,不利于其推广应用。通过建立百子莲组织培养与体细胞胚胎发生体系,可實现百子莲优良品种的无性快繁(李晓丹,2013)。体胚技术诱导的胚性愈伤组织(Embryogenic callus,EC)是植物离体快繁、分子育种及资源保存的重要材料,但长期继代培养会使细胞的胚性逐渐退化甚至丧失,利用玻璃化法超低温保存是实现百子莲种质资源离体保存的重要途径。玻璃化法超低温保存是先将材料放置于由高浓度渗透和非渗透性保护剂组成的玻璃化溶液(Plant vitrification solution,PVS)中,使细胞在快速降温过程中被固化成玻璃化态,以降低胞内致命冰晶的产生,最终在低温条件下实现长期保存(任丽,2014)。然而,植物材料在保存过程中也面临严重脱水、低温和机械损伤等复合胁迫,引起胞内活性氧(ROS)失衡、质膜过氧化、蛋白变性及离子毒害等胁迫伤害(任丽,2014)。降低超低温保存过程中的细胞胁迫伤害可有效提高植物超低温保存效率。因此,通过蛋白质组学研究揭示植物胚性细胞在超低温保存过程中的应答与保护机制,可为补充完善植物超低温保存的逆境生理理论提供科学依据。【前人研究进展】前人已在百子莲胚性愈伤组织超低温保存的逆境生理方面开展了相关研究工作。陈冠群(2014)以添加适当浓度外源抗氧化物的方式优化了百子莲胚性愈伤组织玻璃化法超低温保存体系;何广深等(2014)探究了超低温保存过程中百子莲胚性愈伤组织Ca2+分布情况及浓度变化对细胞适应能力的影响,发现Ca2+主要分布在细胞代谢活动旺盛的区域,且在细胞抗氧化过程中发挥重要作用;Zhang等(2015)揭示了ROS介导的氧化胁迫和细胞凋亡事件是直接影响超低温保存过程中百子莲胚性细胞活性的两个关键因素,其中H2O2是引起氧化损伤最主要的ROS分子。近年来,以RNA-seq和双向凝胶电泳为主的组学技术逐渐应用于一些农林作物和模式植物的超低温保存研究领域。Carpenier等(2007)利用双向电泳技术检测香蕉分生组织在冻存过程中蛋白的差异表达变化,发现一些参与能量代谢和细胞防御、死亡及衰老过程的蛋白上调表达,而参与蛋白合成与细胞生长过程的蛋白下调表达;Forni等(2010)在低温冻存的苹果茎尖组织中发现了与细胞周期减缓和应激反应相关的重要蛋白;Ren等(2013,2015)研究发现,由于冷冻保护剂处理导致的膜脂过氧化是影响拟南芥低温保存成活率的重要原因,随后又从转录水平揭示calcium-binding protein、OXI1、WRKY及MYB家族成员作为ROS信号转导的关键因子,对ROS的产生和清除发挥了重要作用; Folgado等(2014)在马铃薯茎尖组织的冻存研究中发现氧化还原平衡、碳代谢和蛋白修复过程增强,并证实糖类和参与氧化还原过程的蛋白对植物细胞在冷冻保存过程中的耐受性和适应性起重要作用。【本研究切入点】前人研究主要集中在百子莲胚性愈伤组织的超低温保存体系优化和逆境生理方面,但缺乏从蛋白质组学层面对其分子调控机制和蛋白保护功能进行深入探究。【拟解决的关键问题】利用iTRAQ技术结合生物信息学分析技术研究百子莲胚性愈伤组织在超低温保存过程中蛋白的表达情况,从蛋白质组学层面揭示植物胚性细胞对超低温复合逆境的响应调控机制,并筛选出具有重要保护作用的差异表达蛋白,以进一步丰富观赏植物超低温保存的分子生物学理论。
1 材料与方法
1. 1 试验材料
供试植物材料为蓝色大花百子莲(A. praecox ssp. orientails)胚性愈伤组织,由上海交通大学幼苗实践培训基地提供。
主要设备仪器:人工气候箱(RXZ智能型,宁波江南仪器厂)、恒温水浴锅(DK-S22型,上海精宏实验设备有限公司)、LC-20AB液相色谱仪(岛津,日本)和Q-EXACTIVE质谱仪(ThermoFisher Scientific,美国)。
1. 2 试验方法
1. 2. 1 超低温保存方法及取样 百子莲胚性愈伤组织超低温保存参照陈冠群(2014)的方法。主要步骤:(1)预培养:选取状态良好的百子莲胚性愈伤组织,放在含有0.5 mol/L蔗糖的MS固体培养基上,4 ℃低温下暗培养48 h。(2)装载:将预培养后的百子莲胚性愈伤组织加入含有装载液(MS+2.0 mol/L 丙三醇+0.4 mol/L蔗糖+10.0 mmol/L KNO3)的冻存管中,室温下处理60 min。(3)玻璃化溶液脱水处理:将装载液处理后的百子莲胚性愈伤组织移入含有玻璃化保护液PVS2(MS液体培养基+15% DMSO+15%乙二醇+30%丙三醇+0.4 mol/L蔗糖)的冻存管中,在0 ℃下脱水处理50 min。(4)快速冷冻与解冻:液氮冻存1 h后,将放有百子莲胚性愈伤组织的冻存管于40 ℃条件下水浴解冻90 s。(5)洗涤:在室温下使用洗涤液(MS液体培养基+10.0 mmol/L KNO3+1.2 mol/L蔗糖)每间隔10 min对百子莲胚性愈伤组织洗涤一次,重复3次。(6)恢复培养:将洗涤过后的百子莲胚性愈伤组织放在MS固体培养基上,恢复培养24 h。取上述预培养、玻璃化溶液脱水处理、洗涤及恢复培养后的百子莲胚性愈伤组织为试验组样品,以未经处理的百子莲胚性愈伤组织为对照组样品。
1. 2. 2 蛋白提取与测定 分别称取1 g对照组和试验组样品,在液氮中充分研磨成粉末,加入适量蛋白裂解溶液溶解,分别添加终浓度为1 mmol/L的PMSF和2 mmol/L的EDTA,5 min后添加终浓度10 mmol/L的DTT;超声溶解15 min后,25000×g离心20 min取上清液;加入5倍体积预冷丙酮,在-20 ℃沉淀2 h,1600×g离心20 min,收集沉淀;加入适量蛋白裂解溶解液,分别添加终浓度为1 mmol/L的PMSF和2 mmol/L的EDTA,5 min后添加终浓度10 mmol/L的DTT;超声溶解15 min后,25000×g离心20 min取上清;上清液加入DTT至终浓度10 mmol/L,56 ℃水浴1 h;取出后迅速加入IAM至终浓度55 mmol/L,暗室静置45 min;混合样品后再次加入5倍体积预冷丙酮,-20 ℃沉淀2 h;随后25000×g离心20 min取沉淀;沉淀在200 μL 0.5 mol/L TEAB中超声溶解15 min后,25000×g离心20 min后取上清液。利用Brandford法测定蛋白浓度。
1. 2. 3 蛋白酶解与iTRAQ标记 提取后的各样品蛋白经SDS-PAGE分析检测后,精确称取100 μg蛋白按蛋白与酶20∶1的比例加入Tryspin,在37 ℃下酶解2 h,然后按上述比例补加Tryspin再酶解8 h。真空离心泵抽干肽段后,用0.5 mol/L TEAB复溶肽段,利用分子质量为114、116、118、119和121的基团依次标记对照组与试验组(预培养、玻璃化脱水、洗涤及恢复培养)蛋白,室温培养2 h。
1. 2. 4 肽段分离與蛋白鉴定 将标记好的各组肽段等量混合后,采用岛津LC-20 AB系统对样品液相分离,分离后的肽段进入到串联ESI质谱仪Q-EXACTIVE(ThermoFisher Scientific,San Jose,CA),进行液相串联质谱分析。
1. 2. 5 数据处理 利用Mascot(2.3.02)将原始质谱数据与已建立的百子莲转录组数据库(https://www.ncbi.nlm.nih.gov/sra,登录号:SRR850813)搜索匹配。基于数据库搜索策略的肽段质量误差为20 ppm,且蛋白至少含一个特异性肽段,被认为鉴定有效。
1. 2. 6 生物信息学分析 以差异表达变化倍数上调或下调大于1.5倍,筛选出差异表达蛋白。鉴定到的差异表达蛋白进行GO(Gene ontology)注释,利用KEGG(Kyoto encyclopedia of genes and genomes)数据库明确超低温保存四个关键步骤的差异表达蛋白参与的显著性富集通路;使用Genesis(1.8.1)对不同表达模式的差异蛋白进行聚类,并以此为基础在STRING网站(https://string-db.org)上构建上下调差异表达蛋白的互作调控网络。
2 结果与分析
2. 1 iTRAQ分析与差异蛋白筛选结果
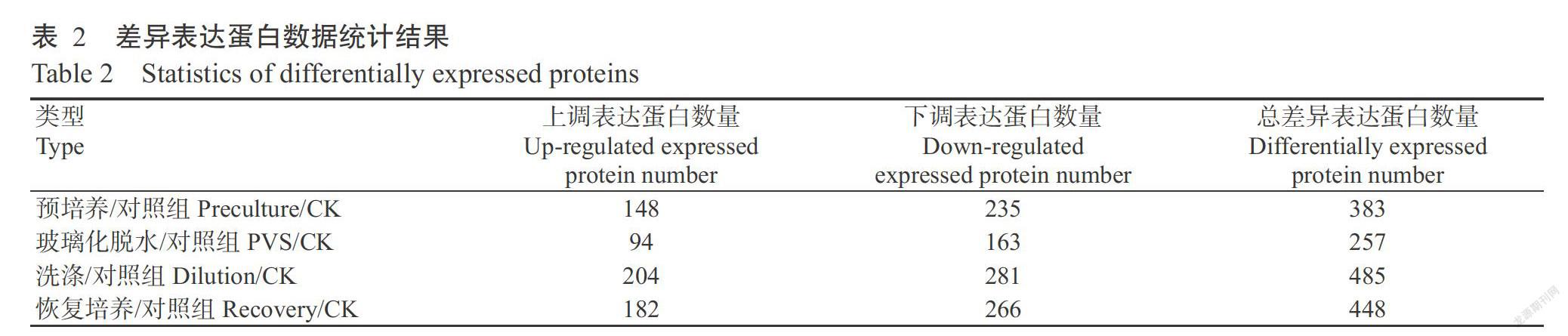
对5个测试的百子莲胚性愈伤组织样品进行蛋白iTRAQ定量分析,共识别获得349591张二级谱图,其中有42008张谱图与已知谱图匹配,含有特异性肽段的质谱图数为37718张;共鉴定到肽段8466条,特有肽段7941条,蛋白2730种(表1)。在全蛋白的基础上筛选出变化倍数大于1.5倍的作为差异表达蛋白(表2),与对照组进行对比,在预培养、玻璃化脱水、洗涤和恢复培养阶段的差异表达蛋白数量依次为383、257、485和448个,四个阶段样品合计共有821个差异表达蛋白。
2. 2 差异表达蛋白的GO功能注释分析结果
对差异表达蛋白进行GO功能注释,以确定其主要行使的生物学功能。结果共有682个差异表达蛋白注释到121个显著性GO条目之中(图1)。在细胞组分(Cell component)中,差异表达蛋白主要涉及细胞(Cell)、细胞器(Organelle)和膜结构(Membrane)等,分别占注释差异表达蛋白的79.6%、59.4%和35.8%;在分子功能(Molecular function)中,差异表达蛋白在催化活性(Catalytic activity)、分子结合(Binding)及结构分子活动(Structural molecule acti-vity)上显著富集,分别占注释差异表达蛋白的59.5%、52.1%和5.4%;而在生物学过程(Biology process)中,差异表达蛋白主要参与了代谢过程(Metabolic process)、细胞过程(Cellular process)和刺激应答(Response to stimulus),分别占注释差异表达蛋白的65.0%、58.4%和36.8%。由此可见,超低温保存中百子莲胚性愈伤组织的刺激应答和代谢过程具有明显变化,并对细胞和细胞膜结构产生明显影响。
2. 3 差异表达蛋白Pathway注释结果
为深入了解超低温保存过程中差异表达蛋白参与的代谢通路,用散点图展示KEGG显著富集的前20条Pathway(图2)。在预培养阶段(图2-A),差异表达蛋白主要富集在代谢(Metabolic pathways)和次生代谢产物的生物合成(Biosynthesis of secondary metabolites)通路;在脱水阶段(图2-B),差异表达蛋白主要参与植物激素的信号转导(Plant hormone signal transduction)和一些氨基酸的代谢活动(Alanine,aspartate and glutamate metabolism);洗涤阶段(图2-C)的差异表达蛋白富集通路明显增多,且以糖代谢为主,主要作用于剪接体(Spliceosome)、鞘脂代谢(Sphingolipid metabolism)、鞘糖脂生物合成(Glycosphingolipid biosynthesis)、糖胺聚糖降解(Glycosaminoglycan degradation)和半乳糖代谢(Galactose metabolism)等;在最终恢复培养过程中(图2-D),显著富集的通路分别是淀粉和蔗糖代谢(Starch and sucrose metabolism)及色氨酸代谢(Tryptophan metabolism)等。由此可见,糖代谢与氨基酸代谢对超低温保存具有积极的响应,在维持细胞的基础生理功能,植物激素信号在脱水保护与冻存过程中发挥重要的调控作用。
2. 4 差异表达蛋白的表达模式分析结果
对所有差异表达蛋白进行聚类,以蛋白变化倍数的log2值绘制聚类分析热图,差异表达蛋白的表达模式共分为六组聚类(图3),其中A组含有96个蛋白,B组含有29个蛋白,C组含有259个蛋白,D组含有104个蛋白,E组含有221个蛋白,F组含有112个蛋白。从总体变化趋势来看,A和C两组差异表达蛋白的趋势相似,在超低温四个阶段中呈上调表达趋势,而B、E和F三组差异表达蛋白表达趋势均下调,D组蛋白的表达水平随超低温处理过程呈波动性变化。
根据聚类分析结果,将差异表达蛋白分为上调表达组(A和C组)和下调表达组(B、E和F组),分别含有355和362个蛋白。经STRING在线软件分析蛋白的互作调控网络,其中上调表达组中有187个蛋白存在相互调控作用(图4),主要参与淀粉和蔗糖的代谢(Starch and sucrose metabolism)、糖酵解与糖异生(Glycolysis/gluconeogenesis)、三羧酸循环(TCA-cycle)、氧化还原过程(Oxidation-reductive process)和核糖体翻译(Ribosome & translation)等,说明在超低温保存过程中糖的代谢、氧化还原平衡及蛋白质加工修饰过程增强;下调表达组有249个蛋白构成互作网络(图5),主要包括ROS代谢(ROS metabolism)、抵御氧化胁迫(Against oxidative damage)、细胞骨架(Cytoskeleton)、结构分子活性(Structural molecule activity)、DNA修复和抑制PCD过程(Inhi-biting PCD)等过程。由此看出,在超低温保存过程中百子莲胚性愈伤组织的氧化应激、胁迫伤害、质膜受损及PCD过程有所加剧。
2. 5 响应超低温复合逆境胁迫的差异表达蛋白
综合鉴定差异表达蛋白的GO功能注释、KEGG代谢通路及蛋白互作网络,进一步筛选得到4类与超低温复合逆境相关的96个蛋白(表3),分别参与能量代谢(Energy metabolism,44个)、氧化还原平衡(Redox homeostasis,18个)、蛋白质合成与降解(Protein synthesis and degradation,17个)及信号转导过程(Si-gnal transduction,17个)。
3 讨论
3. 1 能量代谢相关蛋白
糖代谢是细胞产生和获取能量的主要方式,在植物抵御非生物胁迫时发挥重要作用。本研究发现大量与糖代谢相关的上调表达蛋白,包括参与糖酵解过程的磷酸丙糖异构酶、6-磷酸果糖激酶、丙酮酸激酶和葡糖磷酸酶,参与TCA循环的琥珀酰-CoA合成酶、苹果酸合成酶、酮戊二酸脱氢酶及参与磷酸戊糖途径的转酮醇酶等,其中以参与糖酵解活动的差异表达蛋白居多,说明百子莲愈伤组织在超低温保存过程中主要依赖糖酵解途径产生更多的能量以维持细胞在胁迫环境下正常的生理机能。另外,蔗糖积累也是一种植物对逆境响应常见的保护性反馈方式,在香蕉分生组织及马铃薯茎尖组织的低温冻存中均发现蔗糖含量升高现象(Carpenier et al.,2007;Folgado et al.,2014)。本研究中发现一类上调表达的蔗糖合成酶,可促进细胞在逆境胁迫下蔗糖的积累,对调节细胞的渗透压水平和维持細胞质膜结构的完整性起保护作用。
3. 2 氧化还原平衡相关蛋白
在逆境胁迫下细胞体内会产生过量的ROS,不断积累导致膜质过氧化,对细胞结构及重要蛋白造成不可逆的伤害(Mittler et al.,2004)。植物自身存在一套完整的抗氧化系统,以维持细胞体内的ROS平衡。前期对百子莲超低温保存的逆境生理研究发现,过量ROS诱导的氧化胁迫伤害是影响细胞功能活性的关键因素之一(陈冠群,2014)。本研究发现2种超氧化物歧化酶和3种过氧化物酶呈下调表达,L-抗坏血酸过氧化物酶3和谷胱甘肽过氧化物酶2显著上调表达,前几类下调表达蛋白主要参与抗氧化系统的酶促反应,而后两类上调表达蛋白主要参与谷胱甘肽和抗坏血酸等非酶促抗氧化过程。通过这些差异蛋白的表达变化判断,在超低温保存过程中非酶促抗氧化过程对于清除过量的ROS、缓解细胞的氧化还原平衡状态发挥了重要作用,而高浓度冷冻保护剂可能使得胞内蛋白酶活性下降,进而影响酶促反应的程度。此外,二硫键的氧化还原状态对维持蛋白的正确折叠及高级结构具有重要作用,过量的ROS会影响蛋白间二硫键的平衡状态,导致蛋白的结构改变与失活(王秋霞,2008)。硫氧还蛋白(Thioredoxin,TRX)含有具氧化还原活性的二硫键,可通过硫醇—二硫键的可逆转换对细胞进行氧化还原调节,稳定蛋白的结构与功能(张亚芳,2011)。本研究还发现一类H型TRX,在超低温保存的四个阶段均上调表达,对超低温复合逆境中胞内蛋白结构与酶活性起重要的保护和调控作用。
3. 3 蛋白合成与降解相关蛋白
热激蛋白(Heat shock protein,HSP)是一类重要的抗逆保护蛋白,具有分子伴侣的特性,主要通过蛋白的合成与降解、稳定蛋白结构、细胞定位等多种生物学过程来维持植物体的正常运转(栗振义等,2016)。本研究发现1种上调表达和2种下调表达的HSP,表明不同类型的HSP以多种功能形式参与植物细胞的复合逆境响应。60S核糖体蛋白参与蛋白的翻译与合成,本研究发现四类60S核糖体蛋白呈现不同的表达模式,说明蛋白的合成活动在超低温保存过程中受到一定影响;二硫键异构酶和肽酰—脯氨酰顺反式异构酶在细胞内参与蛋白的折叠、装配及运输等生物学过程(Price et al.,1994),本研究还发现二硫键异构酶2前体和2种肽酰—脯氨酰顺反式异构酶均呈下调表达,表明超低温复合逆境胁迫对胞内蛋白正常的加工与修饰过程具有一定阻碍作用,降低了蛋白的功能活性与结构稳定性。前期对百子莲胚性愈伤组织的超低温保存研究发现,过量H2O2诱导的氧化胁迫与细胞程序性死亡(PCD)是影响细胞超低温保存冻后活性的主要因素(Zhang et al.,2015)。半胱氨酸蛋白酶是一类与植物PCD密切相关的蛋白酶,在番茄、拟南芥和百日菊中参与受损细胞降解与清除的PCD过程(史刚荣,2002);烟草天冬氨酸蛋白酶是在非生物胁迫下诱导细胞发生PCD所必须的蛋白(Chichkova et al.,2010;Vartapetian et al.,2011),枯草杆菌酶的蛋白前体也表现出类半胱天冬酶的活性,同样参与植物的PCD过程(Hatsugai et al.,2009;Tran et al.,2014)。本研究发现这几类重要的蛋白酶多在玻璃化脱水及恢复培养阶段上调表达,推测其参与了由高浓度玻璃化溶液及超低温冻后环境引起的PCD下游蛋白降解过程。
3. 4 信号转导相关蛋白
钙离子是植物体内重要的第二信使,其浓度变化影响着细胞的代谢活动(Bush et al.,1995)。在拟南芥中,钙依赖蛋白激酶(Calcium-dependent protein kinases,CDPKs)和钙调素结合蛋白(Calmodulin-binding protein,CaMBP)可显著提高植物的抗逆性(Xu et al.,2011;Zhao et al.,2013)。在拟南芥幼苗超低温保存的转录组研究中发现,钙结合基因calcium-binding protein表达量显著上调,是ROS信号转导和清除网络的重要因子(Ren et al.,2015)。本研究发现4种CDPKs和CaMBP在蛋白质组学层面均呈下调表达,可能源于植物幼苗与胚性愈伤材料在超低温复合逆境下具有不同的应激响应模式,或CDPKs和CaMBP在超低温复合逆境的转录调控与蛋白表达水平上存在负反馈调节机制。磷酸酶和蛋白激酶作为胞内信号分子可通过调节磷酸化程度控制其他酶类或蛋白质的活性,影响细胞的代谢活动。本研究发现多种类受体蛋白激酶和磷酸酶下调表达,说明在超低温保存过程中,细胞内通过磷酸化和去磷酸化的信号调控方式受到抑制。14-3-3蛋白是一类高度保守的逆境信号响应蛋白(Oecking and Jaspert,2009),在胁迫条件下的甜菜、番茄和柑橘等植物中参与调节离子通道和酶活性(Robert et al.,2002;肖强和郑海雷,2005;Denison et al.,2011)。本研究发现2种上调表达的14-3-3蛋白,推测其在超低温过程中通过参与调节离子通道的活性和数量,从而维持细胞渗透压水平,稳定质膜结构。肌醇单磷酸酶(Inositol monophosphatase,IMP)是磷酸肌醇信号通道的关键酶,在鹰嘴豆(Saxena et al.,2013)、南瓜(王安君等,2016)等植物中发现其在抵抗非生物胁迫和病害方面发挥重要作用。本研究发现1种肌醇单磷酸酶3在超低温过程中上调表达,说明其可能通过调节肌醇的代谢水平对细胞质膜结构发挥重要的渗透保护作用。
4 结论
在百子莲超低温保存过程中,细胞内能量代谢、氧化应激及PCD有所加剧,ROS清除和胁迫防御活动明显减弱,百子莲胚性愈伤组织在超低温复合逆境下形成以糖代谢和抗氧化系统为主的抗逆保护机制。
参考文献:
陈冠群. 2014. 基于拟南芥抗氧化机制优化百子莲胚性愈伤组织超低温保存体系[D]. 上海:上海交通大学. [Chen G Q. 2014. Optimization of Agapanthus embryogenic callus cryopreservation based on the Arabidopsis antioxidant mechanism[D]. Shanghai:Shanghai Jiaotong University.]
何廣深,张荻,申晓辉. 2014. 百子莲胚性愈伤组织在玻璃化法超低温保存4个关键步骤中Ca2+的分布变化[J]. 上海交通大学学报(农业科学版),32(6):14-22. [He G S,Zhang D,Shen X H. 2014. Distribution change of Ca2+ in embryogenic callus of Arabidopsis praecox during the 4 key steps of vitrification cryopreservation[J]. Journal of Shanghai Jiaotong University(Agricultural Science),32(6):14-22.]
李晓丹. 2013. 百子莲胚性愈伤组织玻璃化法超低温保存技术的研究[D]. 哈尔滨:东北林业大学. [Li X D. 2013. Vitrification cryopreservation of embryonic callus of Agapanthus praecox[D]. Harbin:Northeast Forest University.]
栗振义,龙瑞才,张铁军,杨青川,康俊梅. 2016. 植物热激蛋白研究进展[J]. 生物技术通报,32(2):7-13. [Li Z Y,Long R C,Zhang T J,Yang Q C,Kang J M. 2016. Research progress on plant heat shock protein[J]. Biotechnology Bu-lletin,32(2):7-13.]
任丽. 2014. 拟南芥幼苗对玻璃化法超低温保存的逆境响应机制[D]. 上海:上海交通大学. [Ren L. 2014. The regulatory mechanism of Arabidopsis seedlings response to stress from vitrification-based cryopreservation[D]. Shanghai:Shanghai Jiaotong University.]
史刚荣. 2002. 植物细胞程序性死亡的诱导和调控[J]. 生物学教学,27(12):8-9. [Shi G R. 2002. Induction and regulation of programmed cell death in plants[J]. Biology Teac-hing,27(12):8-9.]
王安君,李玉丹,黄河勋,罗少波,吴廷全,李任强,钟玉娟. 2016. 南瓜肌醇单磷酸酶基因CmIMP1和CmIMP2的克隆与表达分析[J]. 园艺学报,43(7):1315-1325. [Wang A J,Li Y D,Huang H X,Luo S B,Wu T Q,Li R Q,Zhong Y J. 2016. Cloning and characterization of two genes encoding myo-inositol monophosphatase 1(CmIMP1)and 2(CmIMP2)from pumpkin[J]. Acta Horticulturae Sinica,43(7):1315-1325.]
王秋霞. 2008. 鹽芥和拟南芥硫氧还蛋白在氧化胁迫中的功能研究[D]. 济南:山东师范大学. [Wang Q X. 2008. The functional research of Thellungiella halophila TRXh and Arbidopsis thaliana TRXh[D]. Jinan:Shandong Normal University.]
肖强,郑海雷. 2005. 14-3-3蛋白与植物细胞信号转导[J]. 细胞生物学学报,27(4):417-422. [Xiao Q,Zheng H L. 2005. The 14-3-3 proteins and plant signal transduction[J]. Chinese Journal of Cell Biology,27(4):417-422.]
张亚芳. 2011. 水稻硫氧还蛋白OsTRXh1结合蛋白的鉴定及功能研究[D]. 石家庄:河北师范大学. [Zhang Y F. 2011. Identification and functional analysis of rice thioredoxin OsTRXh1 interaction proteins[D]. Shijiazhuang:Hebei Normal University.]
Bush D S. 1995. Calcium regulation in plant cells and its role in signaling[J]. Annual Review of Plant Biology & Plant Molecular Biology,46:95-122.
Carpentier S C,Witters E,Laukens K,van Onckelen H,Swennen R,Panis B. 2007. Banana.(Musa spp.) as a model to study the meristem proteome:Acclimation to osmotic stress[J]. Proteomics,7(1):92-105.
Chichkova N V,Shaw J,Galiullina R A,Drury G E,Tuzhikov A I,Kim S H,Kalkum M,Hong T B,Gorshkova E N,Torrance L,Vartapetian A B,Taliansky M. 2010. Phytaspase,a relocalisable cell death promoting plant protease with caspase specificity[J]. The EMBO Journal,29(6):1149-1161.
Denison F C,Paul A L,Zupanska A K,Ferl R J. 2011. 14-3-3 proteins in plant physiology[J]. Seminars in Cell & Development Biology,22(7):720-727.
Folgado R,Sergeant K,Renaut J,Swennen R,Hausman J F,Panis B. 2014. Changes in sugar content and proteome of potato in response to cold and dehydration stress and their implications for cryopreservation[J]. Journal of Proteomics,98(4):99-111.
Forni C,Braglia R,Beninati S,Lentini A,Ronci M,Urbani A,Provenzano B,Frattarelli A,Tabolacci C,Damiano C. 2010. Polyamine concentration,transglutaminase activity and changes in protein synthesis during cryopreservation of shoot tips of apple variety annurca[J]. Cryo Letters,31(5):413-425.
Hatsugai N,Iwasaki S,Tamura K,Kondo M,Fuji K,Ogasa-wara K,Nishimura M,Hara N I. 2009. A novel membrane fusion-mediated plant immunity against bacterial pathogens[J]. Genes & Development,23(21):2496-2506.
Mittler R,Vanderauwera S,Gollery M,Breusegem F V. 2004. Reactive oxygen gene network of plants[J]. Trends in Plant Science,9(10):490-498.
Oecking C,Jaspert N. 2009. Plant 14-3-3 proteins catch up with their mammalian orthologs[J]. Current Opinion in Plant Biology,12(6):760-765.
Price E R,Jin M,Lim D,Pati S,Walsh C T,Mckeon F D. 1994. Cyclophilin B trafficking through the secretory pathway is altered by binding of cyclosporin A[J]. Procee-dings of the National Academy of Sciences,91(9):3931-3935.
Ren L,Zhang D,Jiang X N,Gai Y,Wang M W,Reed B M,Shen X H. 2013. Peroxidation due to cryoprotectant treatment is a vital factor for cell survival in Arabidopsis cryopreservation[J]. Plant Science,212:37-47.
Ren L,Zhang D,Chen G Q,Reed B M,Shen X H,Chen H Y. 2015. Transcriptomic profiling revealed the regulatory mechanism of Arabidopsis seedlings response to oxidative stress from cryopreservation[J]. Plant Cell Reports,34(12):2161-2178.
Robert M R,Salinas J,Collinge D B. 2002. 14-3-3 proteins and the response to abiotic and biotic stress[J]. Plant Molecular Biology,50(6):1031-1039.
Saxena S C,Salvi P,Kaur H,Verma P,Petla B P,Rao V,Kamble N,Majee M. 2013. Differentially expressed myo-inositol monophosphatase gene(CaIMP) in chickpea(Cicer arietinum L.) encodes a lithium-sensitive phosphatase enzyme with broad substrate specificity and improves seed germination and seedling growth under abiotic stresses[J]. Journal of Experimental Botany,64(18):5623-5639.
Tran V,Weier D,Radchuk R,Thiel J,Radchuk V. 2014. Caspase-like activities accompany programmed cell death events in developing barley grains[J]. PLoS One,9(10):e109426.
Vartapetian A B,Tuzhikov A I,Chichkova N V,Taliansky M,Wolpert T J. 2011. A plant alternative to animal caspases:Subtilisin-like proteases[J]. Cell Death & Differentiation,18(8):1289-1297.
Xu G Y,Rocha P S,Wang M L,Xu M L,Cui Y C,Zhu Y X,Xia X . 2011. A novel rice calmodulin-like gene,OsMSR2,enhances drought and salt tolerance and increases ABA sensitivity in Arabidopsis[J]. Planta,234(1):47-59.
Zhang D,Ren L,Chen G Q,Zhang J,Reed B M,Shen X H. 2015. ROS-induced oxidative stress and apoptosis-like event directly affect the cell viability of cryopreserved embryogenic callus in Agapanthus praecox[J]. Plant Cell Reports,34(9):1499-1513.
Zhao Q,Zhang H,Wang T,Chen S X,Dai S J. 2013. Proteomics-based investigation of salt-responsive mechanisms in plant roots[J]. Journal of Proteomics,82:230-253.
(責任编辑 王 晖)
