铅锌胁迫对山苍子幼苗生理生化特性的影响
2019-09-05胡文俐李培旺李昌珠蒋丽娟张良波陈景震马英姿
胡文俐,李培旺,李昌珠,蒋丽娟,张良波,陈景震,马英姿
(1.中南林业科技大学,湖南 长沙 410004;2.湖南省环境资源植物开发与利用工程技术研究中心,湖南 长沙 410004;3.湖南省林业科学院,湖南 长沙 410004)
Pb、Zn重金属土壤污染在我国较为严重,当Pb2+、Zn2+质量分数超过阈值时,就会对植物产生毒害[1]。铅作为植物生长非必需元素,对植物产生毒害作用主要通过抑制植物种子萌发、根系和幼苗生长以及叶片的光合作用[2-3]。锌是植物生长的必需元素,低质量分数的锌对植物生长产生促进作用,过量的锌不仅抑制种子萌发和根系生长,影响光合速率,使植物叶片变黄,甚至会改变细胞膜的结构,导致细胞失活[4]。在自然环境中,铅、锌普遍共存,因而容易形成铅锌以及其它重金属元素的复合污染[5]。目前,对超累积或富集植物修复重金属污染做了较多筛选研究,但这些植物多因生物量小、生长速度慢,在实际应用中受到限制,导致修复复合重金属污染效果不理想[6]。
山苍子Litsea cubeba(Lour.) Pers.是樟科木姜子属落叶灌木或小乔木,为我国南方重要生物质能源和香料树种,具有生长快、生物量大、适应性强、地理分布广的特点[7]。本课题组通过前期调查发现山苍子在铅锌尾矿区生长良好,这表明其对铅锌具有一定的耐受性,具备作为重金属污染地植物修复材料的基础。本文中研究了铅锌胁迫对山苍子幼苗生理生化特征的影响,以期为山苍子修复铅锌污染土壤提供理论依据。
1 材料与方法
1.1 材料处理
供试材料为山苍子1年生实生苗,采自湖南省浏阳苗圃,移栽前用去离子水将植株根部洗净。将长势基本相同的幼苗预培养20 d,再将其移至加入铅锌重金属的盆中,铅和锌分别以Pb(NO3)2、ZnSO4·7H2O形态加入。每盆栽种5株幼苗,3个重复。以未经重金属处理的个体为对照(CK),试验质量分数设计见表1,质量分数梯度设计依据我国土壤环境质量标准设计[8]。
1.2 测定方法
处理后第21天取植株相同部位的叶片,测定其叶绿素、酶活性等生理生化指标。叶绿素含量、超氧化物歧化酶(SOD)、过氧化物酶(POD)、过氧化氢酶(CAT)活性、丙二醛(MDA)、可溶性糖、可溶性蛋白、脯氨酸含量的测定参照张志良[9]的方法。
1.3 数据处理
使用WPS2016整理数据并作图,采用SPSS 13.0进行ANONA方差显著性分析和LSD检验。
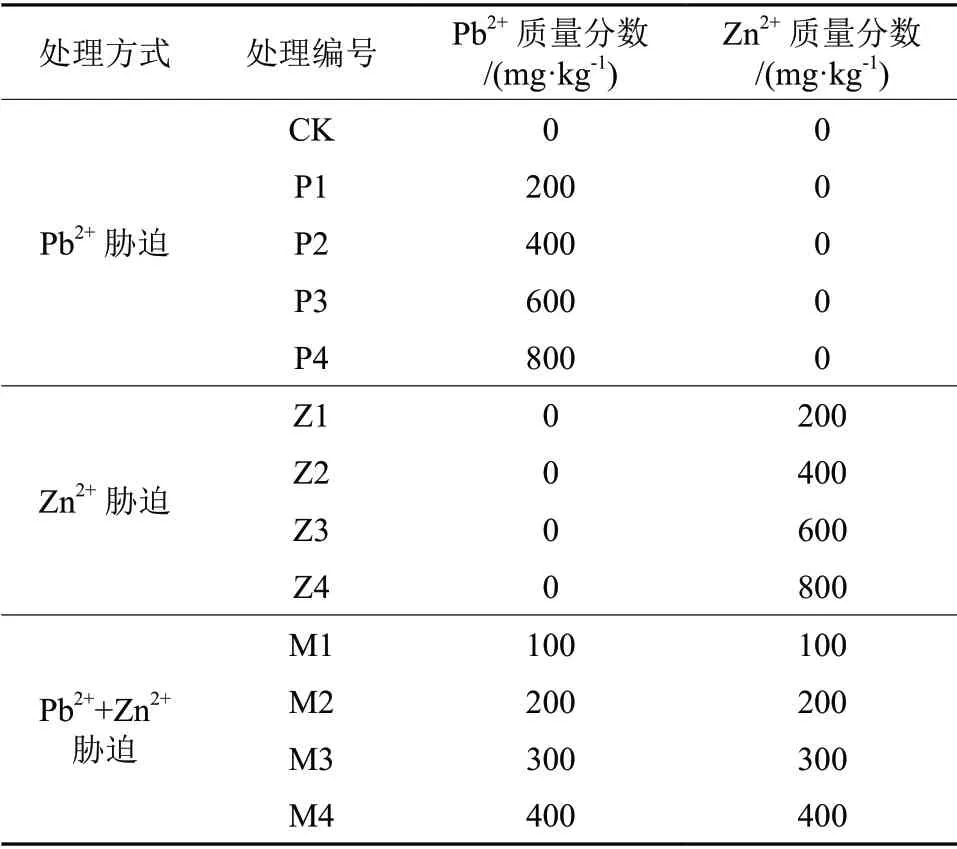
表1 试验设计Table 1 Experiment design
2 结果与分析
2.1 铅锌胁迫对山苍子幼苗叶绿素的影响
在铅锌处理下,山苍子的叶绿素a(Ca)、叶绿素b(Cb)、叶绿素a+b(Ct)、叶绿素a/b (Ca/Cb)的变化趋势如图1所示。在单一Pb2+处理下,当Pb2+为200 mg/kg(P1)时,Cb、Ct含量达到最大值,较对照分别增加了36.38%、6.95%。当Pb2+质量分数≤600 mg/kg时,Ca、Ct含量较对照无显著差异(P>0.05)。当Pb2+为800 mg/kg时,Ca、Ct显著低于对照(P<0.05),较对照分别减少了25.10%、20.21%。在Zn2+处理下,Z1(Zn2+为200 mg/kg)胁迫下Cb、Ct均达到最大值,较对照分别增加了20.95%、2.63%。当Zn2+质量分数≤400 mg/kg时,Ca、Cb、Ct与对照差异均不显著(P>0.05)。当Zn2+质量分数≥600 mg/kg(Z3、Z4)时,Ca、Ct均显著低于对照(P<0.05),较对照分别下降了20.81%、28.70%、15.91%、22.98%。在单一Pb2+、Zn2+处理下,Ca均逐渐下降,Cb、Ct均呈先增后减的趋势。
在Pb2++Zn2+胁迫下,Ca、Cb、Ct均表现出先增后减的趋势,当复合质量分数≤600 mg/kg时,Ca、Cb、Ct值均高于对照,且高于相同离子质量分数的Pb2+、Zn2+单一处理下的各值。在M2处理下,Ca、Cb、Ct达到最大值,最大值分别为1.29、0.57、2.87 mg/g。在M4处 理 下,Ca、Ct显著低于对照(P<0.05),Cb与对照差异不明显(P>0.05)。另外,Ca/Cb值在3种胁迫下均逐渐下降,在P4、Z4、M4处理下较对照组下降显著(P<0.05)。

图1 铅锌胁迫下山苍子幼苗叶绿素值的变化Fig.1 Changes of chlorophyll value of Litsea cubeba seedling under stress of Pb2+ and Zn2+
2.2 铅锌胁迫对山苍子幼苗抗氧化酶活性和丙二醛(MDA)含量的影响
图2显示,叶片内SOD、POD、CAT活性随着处理质量分数的增加呈先上升后下降的趋势,SOD酶活性峰值在Pb2+、Zn2+、Pb2++Zn2+处理下均出现在质量分数为400 mg/kg时,且Pb2++Zn2+处理下的SOD峰值最大,达740.40 U/g。POD、CAT酶活性在Pb2+处理下峰值均出现在P2处理,分别是对照的2.36、2.77倍。在Zn2+、Pb2++Zn2+处理下,POD、CAT酶活性峰值均出现在Z3和M3处理下,分别是对照的3.39、5.48和2.72、2.89倍。3种胁迫方式下,抗氧化酶活性均在800 mg/kg时达到最小值。这表明低质量分数的铅锌对SOD、POD、CAT酶活性表现出不同程度的激活作用。但在高质量分数Zn2+、Pb2++Zn2+(≥600 mg/kg)胁迫下,叶片内的POD、CAT酶活性比同质量分数单一Pb2+胁迫下更强,植物受到的损害更严重。随着胁迫质量分数的升高,山苍子体内的抗氧化酶平衡系统遭到破坏,从而导致酶活性快速下降。
图2表明,3种胁迫下,山苍子叶片内MDA含量均处于上升趋势,在P4、Z4、M4处理下分别达到最大值。这表明随着处理质量分数的增加,山苍子体内MDA的含量有所上升,幼苗受到的伤害逐渐加大。幼苗叶片内MDA含量在低质量分数Pb2++Zn2+(≤400 mg/kg)处理下比相同质量分数Pb2+、Zn2+单一处理下低,在高质量分数Pb2++Zn2+(>400 mg/kg)胁迫下MDA含量比相同质量分数Pb2+、Zn2+单一胁迫下高,说明植物在低质量分数Pb2++Zn2+处理下细胞膜质受损程度相对较轻,在高质量分数Pb2++Zn2+处理下细胞膜质损害相对严重。

图2 铅锌胁迫下山苍子幼苗叶片SOD、POD、CAT、MDA含量的变化Fig.2 Changes of SOD,POD,CAT,MDA contents of Litsea cubeba seedling leaf under stress of Pb and Zn
2.3 铅锌胁迫对山苍子幼苗可溶性糖、可溶性蛋白和脯氨酸含量的影响
由图3可知,在Pb2+、Zn2+及Pb2++Zn2+处理下,叶片可溶性糖、可溶性蛋白含量随着处理质量分数增大均呈先增后减的趋势,可溶性糖分别在P2、Z1、M2处理下达到最大值,分别为8.98、7.52、11.67 mg/g。可溶性蛋白含量分别在P2、Z1、M1处理下达到最大值,较对照分别增加了32.20%、12.58%、41.74%。当胁迫质量分数>400 mg/kg时,随着胁迫程度的加大,糖、蛋白含量在3种胁迫下均显著下降(P<0.05),但下降趋势各不相同。在单一Zn2+处理下,当Zn2+质量分数>200 mg/kg时,山苍子叶片中可溶性糖、蛋白开始下降;在Z4处理下,糖和蛋白含量为所有处理组中的最小值。
图3还显示,叶片内脯氨酸含量随着Pb2+、Zn2+、Pb2++Zn2+处理质量分数增大持续上升,且均在胁迫质量分数为800 mg/kg时达到最大值。当胁迫质量分数为600、800 mg/kg时,Pb2++Zn2+胁迫下脯氨酸含量均高于相同离子质量分数的Pb2+、Zn2+单一胁迫下含量。
2.4 各生理指标与3种处理之间的相关性
由相关性分析可知,叶绿素a与铅锌胁迫之间的相关系数均大于叶绿素b与其相关系数,说明叶绿素a对铅锌胁迫比叶绿素b更敏感。铅锌3种胁迫与叶绿素值的相关性大小为Zn2+>Pb2+>Pb2++Zn2+。SOD与3种胁迫的相关性大小为Zn2+>Pb2+>Pb2++Zn2+;POD与其相关性大小为Zn2+>Pb2++Zn2+>Pb2+;CAT与其相关性大小为Pb2++Zn2+>Zn2+>Pb2+。说明Zn2+和Pb2++Zn2+胁迫与抗氧化酶活相关性较大。可溶性糖与Pb2+、Zn2+处理均为负相关,与Pb2++Zn2+处理呈正相关,可见一定质量分数的Pb2++Zn2+对可溶性糖的形成具有促进作用。可溶性蛋白含量与Zn2+处理之间相关性达显著水平,说明Zn2+对蛋白含量影响较大。山苍子叶片内脯氨酸含量与3种处理均达到极显著正相关,相关性大小为:Zn2+>Pb2++Zn2+>Pb2+。
图3 铅锌胁迫下山苍子幼苗叶片可溶性糖、可溶性蛋白和脯氨酸含量的变化Fig.3 Changes of soluble sugar,soluble protein and free proline content of Litsea cubeba seedling leaf under stress of Pb and Zn
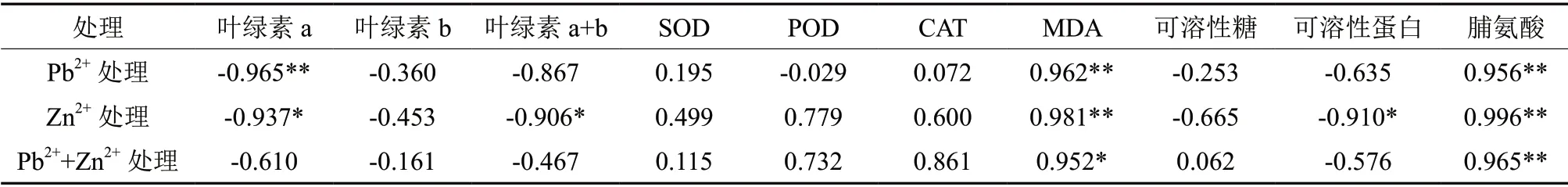
表2 铅锌质量分数与山苍子幼苗生理生化指标之间的相关性分析†Table 2 Correlation analysis between the concentration of Pb,Zn and the physiological and biochemical indices of Litsea cubeba seedlings
3 讨论与结论
叶绿素是植物进行光合作用的主要色素,其含量的高低直接反映植物光合作用和生长能力的强弱,它也用于指示逆境下植物组织、器官的衰老情况[10-12]。本研究结果表明,低质量分数Pb2+、Zn2+(离子质量分数≤400 mg/kg)对山苍子叶绿素的合成具有轻微的促进作用,高质量分数Pb2+、Zn2+(离子质量分数≥600 mg/kg)明显抑制叶绿素的合成,说明高质量分数重金属对植物的损害较大[13]。Pb2++Zn2+复合胁迫在离子质量分数≤400 mg/kg时增加了叶绿素a、b、a+b的含量,这可能是Pb2+、Zn2+复合作用使山苍子对其中一种或两种重金属元素的吸收减少所致,但其影响较为复杂,具体作用机理还有待进一步的研究[14]。叶绿素各值的变化趋势还表明,叶绿素b含量受铅锌影响最小,叶片光合色素对铅锌的敏感程度为叶绿素a>叶绿素b。
SOD、POD、CAT 3种酶在植物体内形成抗氧化系统,该系统可以有效清除活性氧自由基。抗氧化酶活性上升和下降与植物的抗性强弱有关,抗性强的植物在逆境条件下能使酶活性维持在较高水平[15-16]。山苍子体内抗氧化酶活性在低质量分数Pb2+、Zn2+及Pb2++Zn2+胁迫下逐渐上升,在高质量分数(离子质量分数>600 mg/kg)胁迫下体内SOD、POD、CAT酶活性下降,但仍保持在较高水平。说明在低质量分数重金属胁迫下,植株体内的防御机能被激发,抗氧化酶活性迅速上升以应对重金属引起的活性氧增加所带来的危害。但随着胁迫质量分数的增大,重金属在植物体内累积,使得植株生理代谢紊乱,抗氧化酶活性受到抑制并开始下降[17]。该研究表明抗氧化酶在山苍子应对铅锌胁迫的过程中发挥了积极作用。因此,山苍子可通过自身代谢降低铅锌重金属的毒害,增强对铅锌的耐受性。
植物在逆境下,机体细胞一般会发生膜脂过氧化,丙二醛(MDA)是膜脂过氧化的最终产物,其含量的多少可以反映植物遭受逆境伤害的程度大小[18]。山苍子幼苗MDA含量在3种胁迫下均随着胁迫质量分数增大而逐渐上升,叶片内MDA含量在铅锌质量分数为800 mg/kg时均达到最大值,此时植株内细胞膜质的过氧化程度最高,细胞膜受损程度最为严重[19]。这与铅、镉胁迫下中华结缕草和皱叶酸模内MDA含量以及铜锌复合胁迫下芦竹内MDA含量变化结果一致[20-21]。
可溶性糖、可溶性蛋白和脯氨酸作为细胞膜渗透性保护物质,其含量可反映植物在遭受重金属毒害时对外界环境的适应能力大小[22]。本研究发现,在Pb2+、Zn2+及Pb2++Zn2+处理下,山苍子可溶性糖、可溶性蛋白含量均表现出先上升后下降的趋势,这说明山苍子在受到重金属胁迫时,通过调节碳、氮代谢产物的平衡关系,提高了植物体对铅锌胁迫的耐性[23]。这与张静等[24]的研究结果一致。低质量分数的铅锌胁迫促进山苍子幼苗体内可溶性糖和蛋白的合成,使得山苍子在逆境下的抗性增强。随着胁迫质量分数的升高,高质量分数铅锌胁迫使得细胞结构遭到破坏,导致植物渗透调节失衡,阻碍了可溶性糖、可溶性蛋白的合成,从而对幼苗产生毒害[25]。机体内脯氨酸的含量与植物抗逆性的能力紧密相关[26],山苍子叶片中脯氨酸的含量在3种胁迫下持续上升,产生这种情况的原因可能是当铅锌含量较低时,对幼苗毒害作用相对较小,但随着处理质量分数的增大,山苍子体内的正常代谢和酶系统遭到破坏,迫使植物机体做出更为复杂的应急反应,导致大量脯氨酸累积[27]。
在低质量分数复合处理下,山苍子叶绿素各值变化趋势与Pb2+、Zn2+单一处理下相同,但高质量分数Pb2++Zn2+胁迫(离子质量分数>600 mg/kg)较同质量分数Pb2+、Zn2+单一胁迫对叶绿素含量具有更显著的抑制作用。另外,本研究中的相关性分析表明,重金属Zn2+对山苍子的生长影响较大。在高质量分数Zn2+、Pb2++Zn2+(质量分数≥600 mg/kg)胁迫下,叶片内的POD、CAT酶活性比同质量分数单一Pb2+胁迫下更强,植物损坏程度更严重,这与易心钰[28]研究的蓖麻对铅锌胁迫的响应及其机制的研究结果基本一致。可能是因为重金属Zn2+作为植物生长的必需元素,当其含量超过植物所需时,山苍子体内的酶系统遭受的破坏更严重,对植物的生长影响更大[5]。而且在高质量分数Pb2++Zn2+胁迫下幼苗抗氧化酶活性、脯氨酸、丙二醛含量均高于相同质量分数Pb2+、Zn2+单一胁迫。出现此现象的原因可能是在高质量分数Pb2++Zn2+复合胁迫下,Pb2+与Zn2+表现为协同作用,对植株毒害作用增强,这与杨玲等[29]研究的Pb、Zn胁迫对狗牙根和早熟禾的研究结果相同。不过复合污染体系中重金属的复合作用是一个复杂的问题,它与重金属的种类和质量分数组合、植物所处生命阶段、植物种类、胁迫时间及胁迫方式等诸多因素有关[30]。
通过盆栽试验,初步证明了山苍子对铅锌具有较强的耐受性,且山苍子为南方地区的优势植物,可用于修复铅锌污染土壤,并可有效避免外来物种入侵,维护当地生态系统的平衡[31-32]。但在自然环境中铅锌污染要比盆栽试验复杂得多,植物对重金属吸收的影响还受土壤性质、土壤微生物及根际环境的影响,以后的研究可放在不同土壤类型的大田中进行[33],而且可进一步从组织化学和分子水平上探讨山苍子的耐性机理,以确定重金属对山苍子的作用机制。另外,作为经济作物的山苍子,其果实中重金属的含量必须确定是否超标,以防通过食物链进入人体,对人体造成危害[34]。
