风箱果(♀)×紫叶风箱果(♂)杂交种光合特性对干旱胁迫的响应
2019-09-05张书博王均睿徐梓淞马松良吴绪叶张会慧孙广玉
张书博,王均睿,周 涛,徐梓淞,马松良,吴绪叶,张会慧,孙广玉
(1.东北林业大学 生命科学学院,黑龙江 哈尔滨 150040;2.东北农业大学 资源与环境学院,黑龙江 哈尔滨 150030)
植物生长发育主要依靠其光合生理过程获取物质和能量[1-2],而植物光合作用是对干旱胁迫较为敏感的过程之一。干旱胁迫常导致植物叶片失水萎蔫甚至卷曲,叶片的光能捕获有效面积降低[3],叶绿素含量降低[4],植物的生长及观赏效果降低。另外,干旱胁迫下植物为减少水分的蒸腾散失,叶片常以气孔关闭的方式来提高水分利用率[5]。除气孔因素限制外,非气孔因素中的碳同化能力及PSⅡ光化学活性的降低等因素也会导致植物的生长受到抑制[6]。光系统II(PSII)作为植物光合机构中的光反应中心,主要负责光能的吸收、水的光解和电子传递等一系列光合生理过程[7-8]。干旱胁迫下常抑制PSII光合电子传递,导致其光抑制的发生[9],光合PSII功能受到抑制后还会导致光合作用介导的活性氧爆发,进一步加剧光抑制程度的发生,甚至导致光氧化或光漂白等[10]。
由于自然干旱以及园林绿化灌溉条件等因素的限制,城市园林绿化植物常受到干旱胁迫的影响。因此,干旱胁迫下植物的光合生理响应过程研究对抗旱物种的筛选和提高植物抗旱能力具有重要的意义。风箱果Physocarpus amurensis和紫叶风箱果Physocarpus opulifolius为蔷薇科风箱果属落叶灌木。由于风箱果花粉活力较弱以及生境的人为破坏等因素导致其分布范围和种群数量日益减少[11]。紫叶风箱果为紫叶观赏植物,是我国近年来从北美引入的观赏性花灌木[12],虽然紫叶风箱果较风箱果具有形态和繁殖优势,但其抗旱、耐盐、耐弱光性较差[13-15]。杂种优势是生物界普遍存在的生物学现象,特别是在逆境条件下,众多杂交种植物的抗逆能力较亲本明显提高[16-18]。黑龙江省森林植物园郁永英等人[19-20]通过以本地的风箱果(♀)和紫叶风箱果(♂)杂交成功得到了有紫叶观赏效果的F1代杂交风箱果,并且其光合能力表现出明显的杂种优势。但是关于杂交风箱果对干旱胁迫的适应性方面的研究较少,本试验以风箱果(♀)、紫叶风箱果(♂)和杂交风箱果F1代幼苗扦插苗为试验材料,研究了3种风箱果叶片光合特性对干旱胁迫的响应,探讨了3种风箱果光合能力的抗旱差异,明确不同品种风箱果的抗旱能力,为指导风箱果属园林植物的合理引种及栽培提供一些基础数据。
1 材料与方法
1.1 试验材料与处理
试验于2015年8月在东北林业大学植物生理实验室进行,试验材料为黑龙江省森林植物园提供的3年生风箱果、紫叶风箱及其杂交种F1代的扦插苗。3种材料均于2012年进行扦插育苗,扦插成活的幼苗移植于开口直径28 cm、底直径15 cm、高20 cm的塑料花盆中,每盆定植1株,培养基质为草碳土,室外自然环境培养,常规养护管理。干旱胁迫处理于2015年8月10日进行,选择长势相对一致的3种风箱果品种各3株,即3次重复。干旱胁迫处理之前均浇足水,然后进行自然干旱处理,分别在干旱处理后的第1、4、7天和第10 天进行各风箱果品种叶片光合气体交换和叶绿素荧光参数的测定,并于干旱处理第1天和第10天对不同处理材料进行拍照,观察植株长势及表型。3种风箱果品种处理前分别测定其叶片中花色素苷含量及叶绿素含量。
1.2 测定项目和方法
光合气体交换参数的测定:各处理3种风箱果选择新生枝条上倒数第2片完全展开叶,利用Li-6400光合测定系统(Licor公司,美国)将光强PFD设定为1 000 μmol·m-2s-1,分别测定对照和干旱处理下2种风箱果叶片的净光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)和胞间CO2浓度(Ci)等参数。光合气体交换参数测定时气路用空气缓冲瓶连接大气,大气CO2浓度约为400 μL·L-1,每个处理测定5次。
叶绿素荧光参数的测定:将不同处理3种风箱果幼苗倒数第3片或第4片叶片利用暗适应夹进行0.5 h暗适应,然后采用便携式脉冲调制荧光仪FMS-2(Hansatch公司,英国)测定初始荧光(Fo)、最大荧光(Fm)、PSⅡ最大光化学效率(Fv/Fm)、电子传递速率(ETR)、光化学淬灭系数(qP)、非光化学淬灭(NPQ)等,并计算过剩光能(1-qP)/NPQ,均测定5次。
1.3 数据处理方法
运用Excel2003和SPSS22.0软件进行数据统计分析,采用单因素方差分析(One-way ANOVA)和最小显著差异法(LSD)比较不同数据组间的差异。
2 结果与分析
2.1 干旱胁迫对3种风箱果叶片光合气体交换参数的影响
由图1可以看出,随着干旱天数的增加,3种风箱果叶片的Pn均呈明显的降低趋势,在干旱第1天时,风箱果(♀)叶片的Pn稍低于紫叶风箱果(♂)和杂交风箱果,但是随着干旱天数的增加,紫叶风箱果(♂)叶片Pn的降低幅度明显大于风箱果(♀)和杂交风箱果,特别是在干旱第7天和第10天紫叶风箱果(♂)叶片的Pn均显著低于风箱果(♀)和杂交风相果,但杂交风箱果与风箱果(♀)均无显著差异。干旱胁迫下3种风箱果叶片的Gs、Tr和Ci变化趋势相似,在干旱第7天,3种风箱果叶片的Gs和Tr均急剧降低,并且Ci也有小幅度降低。在干旱第1天和第4天,3种风箱果叶片的Gs、Tr和Ci均表现为杂交风箱果>紫叶风箱果(♂)>风箱果(♀),但在干旱第7天和第10天时,紫叶风箱果(♂)叶片的Gs和Tr均低于风箱果(♀)和杂交风箱果,但三者差异均不显著,并且在干旱第10天时,风箱果(♀)叶片的Ci分别低于紫叶风箱果(♂)和杂交风箱果的17.70%(P<0.05)和18.29%(P<0.05)。
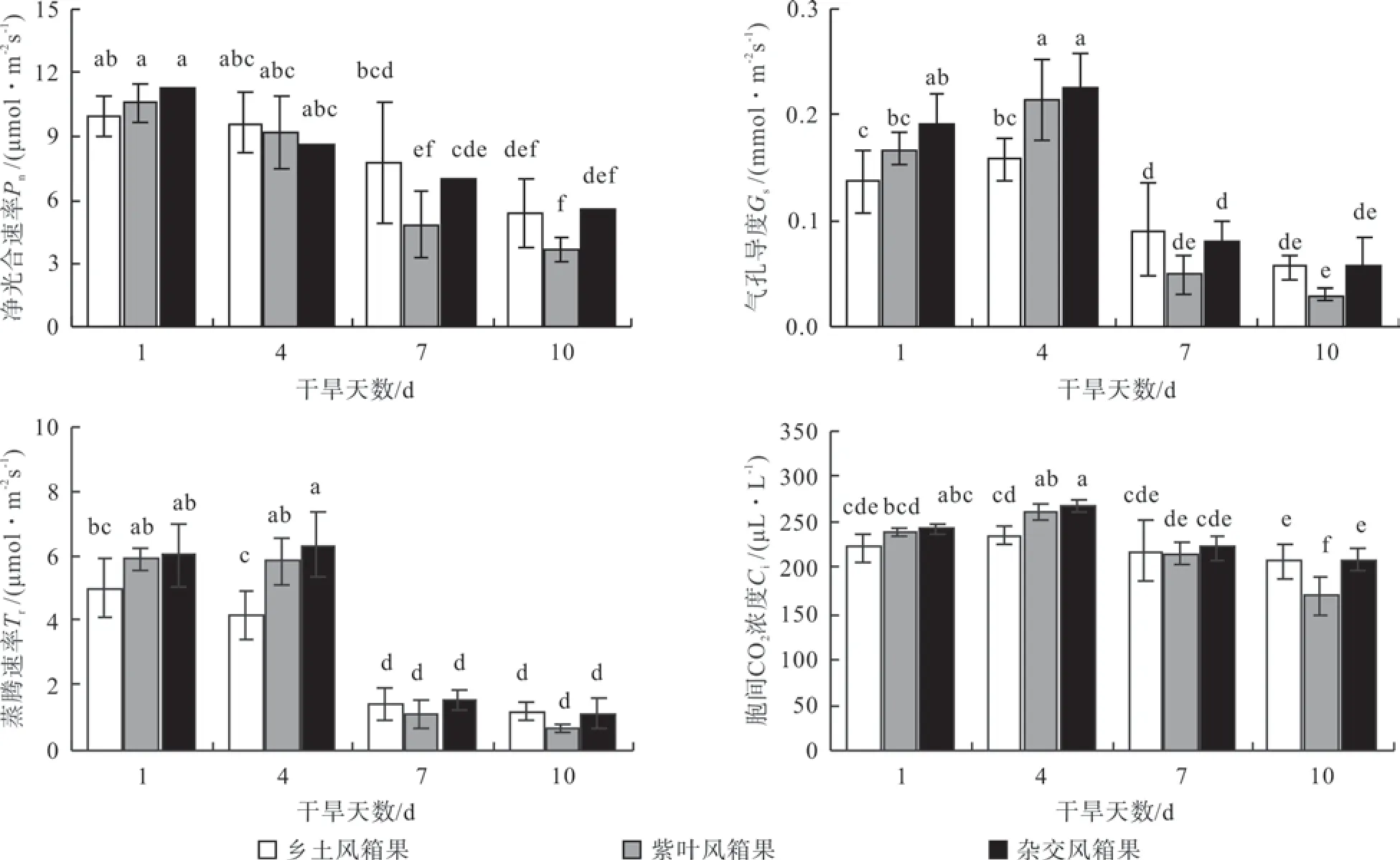
图1 干旱胁迫对3种风箱果叶片光合气体交换参数的影响Fig.1 Effects of drought stress on gas exchange parameters in leaves of three cultivars Physocarpus seedlings
2.2 干旱胁迫对3种风箱果片PSⅡ最大光化学效率(Fv/Fm)和电子传递速率(ETR)的影响
由图2可以看出,3种风箱果叶片的Fv/Fm和ETR在干旱胁迫下均表现为降低趋势,且ETR的降低幅度大于Fv/Fm。在干旱第1天和第4天时,3种风箱果叶片的Fv/Fm和ETR均无显著差异(P>0.05),但随着干旱时间的延长,紫叶风箱果(♂)叶片Fv/Fm和ETR的降低幅度明显大于风箱果(♀),在干旱第10天时,紫叶风箱果(♂)叶片的Fv/Fm显著低于风箱果(♀),而杂交风箱果却与风箱果(♀)无显著差异。并且在干旱第7和10天时,紫叶风箱果叶片(♂)的ETR均显著低于风箱果(♀),而杂交风箱果虽也稍低于风箱果(♀),但差异均不显著。
2.3 干旱胁迫对3种风箱果叶片光化学淬灭系数(qP)、非光化学淬灭(NPQ)和过剩光能(1-qP)/NPQ的影响

图2 干旱胁迫对3种风箱果叶片PSⅡ最大光化学效率(Fv/Fm)和电子传递速率(ETR)的影响Fig.2 Effects of drought stress on PSⅡ maximum photochemical efficiency (Fv/Fm) and electron transport rate (ETR) in leaves of three cultivars Physocarpus seedlings
由图3可以看出,随着干旱天数的增加,3种风箱果叶片的qP呈明显降低趋势,但是紫叶风箱果(♂)叶片的qP的降低幅度均明显大于风箱果和杂交风箱果,干旱第7天和第10天时,紫叶风箱果(♂)叶片的qP均显著低于风箱果(♀),而杂交箱果叶片qP虽也稍低于风箱果(♀),但差异均不显著。风箱果(♀)和杂交风箱果叶片的NPQ随着干旱天数的增加整体呈增加趋势,而紫叶风箱果(♂)则呈先增加后降低趋势,在干旱第10天时,紫叶风箱果(♂)叶片的NPQ较干旱第7天时降低了72.67%(P<0.05)。在干旱前7 d,3种风箱果叶片的(1-qP)/NPQ没有发生明显变化,但在干旱第10天时,紫叶风箱果(♂)叶片的(1-qP)/NPQ剧增,而风箱果(♀)和杂交风箱果仍没发生显著变化。

图3 干旱胁迫对3种风箱果叶片光化学淬灭系数(qp)、非光化学淬灭(NPQ)和过剩光能(1-qp)/NPQ的影响Fig.3 Effects of drought stress on photochemical quenching coefficient (qP),non-photochemical quenching (NPQ) and excess energy (1-qP)/NPQ in leaves of three cultivars Physocarpus seedlings
3 讨 论
光合作用是植物有机物质积累、生长发育的基础,受植物本身生理特性和环境因素的共同影响,其中土壤水分是影响光合生理过程的重要环境因素之一,植物光合能力的高低也是反映植物抗旱性强弱的重要指标[21]。气孔是植物与环境之间进行气体和水分交换的“门户”,干旱胁迫下植物叶片为保证生理代谢所需的水分,植物叶片常常通过降低气孔导度的方式以降低叶片的水分蒸腾作用,以减少水分的散失和提高水分利用效率[22],但是,在降低水分散失的同时气孔导度的降低也限制了碳同化原料-CO2的吸收,增加了CO2向叶内的扩散阻力,导致净光合速率的降低[23-24]。本试验中,随着干旱天数的增加,3种风箱果叶片的Gs和Tr均呈明显的降低趋势,即在干旱胁迫下3种风箱果均可以通过降低气孔导度的方式减少水分的蒸腾。在干旱第7天和第10天时,3种风箱果叶片的Gs和Tr迅速降低,但其Ci降低幅度却较小,根据Farqhar的气孔限制观点[25],干旱胁迫后期引起3种风箱果叶片光合能力降低的原因不仅仅是气孔因素限制,还可能与叶肉细胞对CO2的利用能力降低有关。3种风箱果比较可以看出,在非干旱条件下,紫叶风箱果(♂)叶片的Pn、Gs、Tr和Ci均显著高于风箱果(♀),说明紫叶风箱果(♂)在正常水分条件下具有相对较强的光合能力。紫叶风箱果(♂)叶片光合能力虽然较强,但对土壤水分的依赖性较强,光合碳同化过程对干旱胁迫较为敏感,随着干旱天数的增加,紫叶风箱果(♂)叶片含水率迅速降低,并且其叶片的净光合速率降低幅度远大于风箱果(♀)。但与2个亲本相比,杂交风箱果叶片的光合能力在非干旱胁迫下不仅与紫叶风箱果(♂)接近,并且在干旱胁迫下的降低幅度明显低于紫叶风箱果(♂),说明干旱胁迫下杂交风箱果叶片的光合能力较紫叶风箱果(♂)明显提高。
随着干旱天数的增加,3种风箱果叶片的Fv/Fm、ETR和qP随着干旱天数的增加均呈降低趋势,说明干旱胁迫首先会导致3种风箱叶片的电子传递速率降低,甚至还会导致光抑制的发生。干旱第7天和第10天,风箱果(♀)和杂交风箱果叶片的Fv/Fm、ETR和qP降低幅度均明显小于紫叶风箱果(♂),说明紫叶风箱果(♀)叶片PSII光化学活性和电子传递过程对干旱胁迫较为敏感,而杂交风箱果叶片的Fv/Fm、ETR和qP与风箱果(♀)均无显著差异,即杂交风箱果叶片PSII性能较紫叶风箱果(♂)明显提高。逆境下如果PSⅡ电子传递链上的电子传递受阻,PSⅡ光解H2O会形成的跨膜质子梯度会激活依赖跨膜质子梯度的叶黄素循环进行能量耗散,减轻PSII反应中心的压力[26]。非光化学淬灭NPQ与依赖叶黄素循环的能量耗散成正相关[27],因此,逆境下NPQ作为一各重要的耗散激发能途径,在保护PSⅡ反应中心免受过剩激发能和活性氧的伤害方面具有重要作用[28-29]。干旱胁迫下风箱果(♀)叶片NPQ显著增加,而紫叶风箱果(♂)叶片的NPQ则在干旱第7天增加到最大值后,干旱第10天显著降低,说明干旱胁迫下风箱果(♀)可以通过启动依赖于NPQ的能量耗散机制来提高PSⅡ的抗旱性,而紫叶风箱果(♂)在干旱第10天时这种抗旱机制受到显著抑制,从而导致其过剩激发能(1-qP)/NPQ剧增。本研究仅通过光合气体交换参数和叶绿素荧光参数探讨了风箱果(♀)×紫叶风箱果(♂)杂交种的光合能力对干旱胁迫的响应,但关于杂种优势的具体内在机理有待进一步研究。
4 结 论
干旱胁迫下3种风箱果叶片PSⅡ反应中心活性降低,光合碳同化过程明显受到抑制,特别是紫叶风箱果(♂)更易在干旱胁迫后期叶片失水严重,并且发生非气孔因素的光合限制。风箱果(♀)在干旱胁迫下叶片可以保持相对较高的水分,并且可以通过增加NPQ的方式来降低过剩下光能的产生,降低其叶片的光抑制程度,提高电子传递的稳定性。风箱果(♀)光合特性的耐旱性明显高于引种紫叶风箱果(♂),而以风箱果(♀)×紫叶风箱果(♂)杂交获得的偏父本(♂)表型的杂交风箱果不但具有紫叶风箱果(♂)的观赏效果,而且其抗旱能力明显提高,在干旱胁迫下具有较高的光合能力,表现出明显的杂种优势。
