镉胁迫对龙葵叶绿素荧光和光合生化特性的影响
2019-09-05唐星林金洪平刘光正王丽艳
唐星林,金洪平,周 晨,刘光正,王丽艳
(江西省林业科学院,江西 南昌 330000)
由于矿产资源的不合理开采、工业废水的排放等原因,镉(Cd)已成为土壤的主要污染物之一。土壤Cd可以被植物根系吸收并积累在植物体内。当植物体内Cd浓度达到一定水平,Cd会对植物产生毒害作用[1]。Pahlsson等[2]发现植物叶片积累3~10 μg·g-1Cd就会对植物产生毒害。有研究表明:Cd过量积累会抑制植物根和茎的生长,导致叶片卷曲[3],影响氮、铁等营养元素的吸收[4],产生氧化胁迫[5],损害植物光合机构,抑制光合作用,导致植株生物量下降[6]。Cd毒害可能通过不同机制来抑制叶片光合作用。有研究表明,Cd胁迫会抑制黄菖蒲[7]和烤烟[8]叶绿素的合成。Dalla等[9]发现Cd胁迫会破坏Elodea canadensis叶绿体类囊体膜结构,抑制叶绿体的增殖。大量研究表明,Cd胁迫会导致植物叶片PSII最大光化学效率 (Fv/Fm)、PSII光化学效率(ΦPSII)、光化学淬灭系数(qP)和PSII的电子传递速率(J)等光合参数下降[8,10-11],但也有研究表明Cd胁迫对叶片Fv/Fm、ΦPSII和qP等参数无显著影响[12-13]。Krantev等[14]发现Cd胁迫抑制玉米叶片核酮糖-二磷酸羧化酶(Rubisco酶)活性,这可能导致Rubisco酶最大羧化速率(Vcmax)下降[10],进而限制光合作用。有研究表明Cd胁迫会导致植物叶片气孔密度[13]和气孔大小[10]显著降低,并导致气孔开放数减少[15],进而引起气孔导度(gs)下降。人们认为气孔限制、叶肉限制和光合生化限制是导致叶片光合作用下降的3个主要原因[16]。Tang等[13]发现Cd胁迫对滇苦菜光合作用的限制主要来自于气孔限制和叶肉限制,而生化限制的作用较小。由于植物种类、Cd浓度和胁迫时间的差异,Cd胁迫对叶片光合生理生化特性的影响并不一致。
龙葵Solanum nigrum是一种Cd超积累植物,具有耐Cd毒害和富集Cd的能力,是实施植物修复的优良材料。郭智等[17]发现Cd胁迫会抑制龙葵叶片光合作用,但Cd胁迫下龙葵光合生理响应及其主要光合限速因子还不清楚。鉴于此,本研究通过研究Cd胁迫对龙葵叶绿素荧光特性、光合光响应和光合CO2响应的影响,分析Cd胁迫对龙葵叶片光合生化特性的影响,明确Cd胁迫下龙葵光合作用的主要限制因子,深入了解Cd毒害的光合生化机制,旨在为超积累植物耐性机理研究提供理论依据。
1 材料与方法
1.1 材料与处理
2018年3月12 号,选取饱满的龙葵种子,10 g·L-1KNO3浸泡4 h,用超纯水冲洗干净,播种于盛放播种基质的穴盘中,放置于江西省林业科学院科研大棚。待幼苗长出4片真叶后,用自来水将幼苗根系附着的基质冲洗干净,并进行营养液栽培,每4 d更换一次营养液。营养液含 1.99 mmol·L-1K2SO4、4 mmol·L-1CaCl2、1.01 mmol·L-1KH2PO4、6.25 mmol·L-1NH4NO3、2 mmol·L-1MgSO4·7H2O、100.32 μmol·L-1H3BO3、131.95 μmol·L-1MnSO4·H2O、29.97 μmol·L-1ZnSO4·7H2O、5 μmol·L-1KI、0.1 μmol·L-1CuSO4·5H2O、0.11 μmol·L-1CoCl2·6H2O、1.03 μmol·L-1Na2MoO4·2H2O、100 μmol·L-1FeSO4·7H2O2、100.2 μmol·L-1EDTANa2。栽培30 d后进行如下处理:对照(CK),不加任何药品;镉胁迫(Cd),添加40 μmol·L-1CdCl2。处理30 d后进行实验测定。
1.2 试验方法
1.2.1 叶绿素相对含量和叶绿素荧光参数的测定
叶片叶绿素相对含量采用SPAD-502 叶绿素仪测定,每株植物随机选取生长旺盛的叶片8片,测定其叶绿素值,取平均值。植物叶片进行暗处理30 min后,采用便携式脉冲调制叶绿素荧光仪(FluorPen FP 100,Czech Republic)测定初始荧光(Fo)、最大荧光(Fm)、Fv/Fm、qP和非光化学淬灭系数(NPQ)。每个处理重复测定5株。
1.2.2 光响应曲线和CO2响应曲线的测定
采用美国LiCor公司的便携式光合作用仪(Li-6400XT)进行测量。实验均在晴天进行,时间段为9:00 —16:30。植物叶片先在近饱和光合有效辐射(PAR)1 200 μmol·m-2s-1、样本室CO2浓度400 μmol·mol-1、温度为(26±2)℃、空气相对湿度60% ~ 70%条件下,诱导30 min以上,直到光合速率和气孔导度相对稳定,采用仪器内置自动测量系统把PAR设定为1 200、1 000、800、600、400、300、200、150、100、50、20、0 μmol·m-2s-1,测定净光合速率数据,每个PAR下平衡时间为2 ~ 3 min。每个处理重复测定5片叶。植物叶片在PAR1 200 μmol·m-2s-1、参比室CO2浓度400 μmol·mol-1、温度(26±2)℃、空气相对湿度60% ~ 70%条件下,诱导30 min以上,直到光合速率和气孔导度相对稳定,利用Li-6400-01液化钢瓶控制参比室中的CO2浓度依次为400、300、200、100、50、450、600、800、1 000、1 200 μmol·mol-1并测定净光合速率。每种植物重复测量5片叶。
1.2.3 植株生物量和比叶重的测定
CO2响应曲线测定结束后立即采集叶片并测定其比叶重。植物收获后,将其分为根、茎和叶3部分。根系用去离子水冲洗干净。植物样品经105 ℃杀青30 min,70 ℃下烘至恒质量,并测定其生物量。
1.2.4 数据分析
采用直角双曲线修正模型[18]拟合光响应曲线,获得最大净光合速率(Amax)、表观量子效率(AQY)、光补偿点(LCP)、饱和光强(LSP)和光下暗呼吸速率(Rd)等光合光响应参数。直角双曲线修正模型的描述如下:
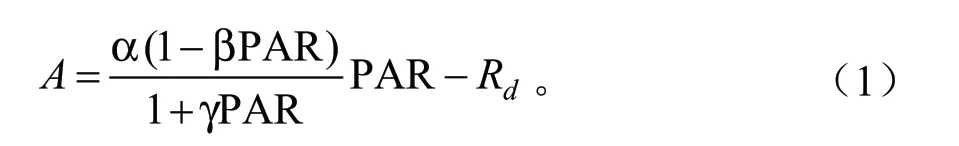
式中:α为初始量子效率;β、γ为模型系数。
令A=0,计算LCP。LSP为:

Amax为:

采用FvCB光合模型拟合CO2响应曲线,获得Vcmax、Jmax、Rubico酶活性限制阶段与RuBP再生速率限制阶段的临界胞间CO2浓度(Ci_CJ)等光合生化参数。Rd采用光响应曲线的拟合值作为输入值,拟合算法参考Gu等[19]的方法。FvCB光合模型表达式为:

式中:A为净光合速率;Ac为Rubisco酶活性限制阶段的净光合速率;Aj为RuBP再生速率限制阶段的净光合速率。
当光合作用受Rubisco酶活性限制时,净光合速率(Ac)为:

式中:Kc为RuBP酶羧化反应米氏常数(27.238 Pa);Ko为RuBP酶氧化反应米氏常数(16 582 Pa);Ci为胞间CO2浓度;Γ*为缺乏暗呼吸的叶绿体CO2补偿点(3.743 Pa);O为氧气浓度(21 220 Pa);Rd为光下暗呼吸速率。
当光合作用RuBP再生速率限制时,净光合速率(Aj)为:

式中:J为PSII电子传递速率;J与Jmax的关系常用经验公式进行描述[14]。
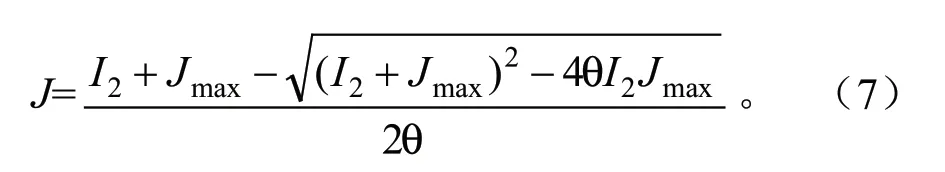
式中:θ为非直角双曲线函数曲率(经验值为1);I2为PSII吸收的有效光辐射。

式中:PAR为光合有效辐射;α1为叶片的光吸收系数(经验值为0.85);fⅡ为光能在光系统I(PSI)和光系统II(PSⅡ)中分配的比例(经验值为0.5);ΦPSIImax为最大PSII光化学效率(经验值为1)。
从CO2响应曲线中获取1 200 μmol·mol-1CO2下的净光合速率(A)、CO2气孔导度(gsc)和胞间CO2浓度(Ci)等光合参数进行分析。在1 200 μmol·mol-1CO2浓度下龙葵叶片光合作用受RuBP再生速率限制,参考Wilson的方法[20],利用A、gsc和Jmax参数来量化气孔限制(Stomatal limitation,SL)和非气孔限制(Nonstomatal limitation,NSL)的大小。本研究以对照组的光合参数作为最大参考值来进行标准化,对获得的数据进行如下分析:采用Dixon法去除异常值,进行Kolmogrov-Smirnov正态性检验,采用独立样本t检验对获得的参数进行大小比较(P<0.05);对于不服从正态分布或方差非齐性的参数,采用wilcoxon秩和检验进行大小比较(P<0.05)。对于服从正态分布的参数,进行Pearson积矩相关分析;不服从正态分布的参数,进行Spearman秩相关分析。利用Microsoft Excel 2016和R语言(R-3.5.1)进行数据整理、分析和作图。
2 结果与分析
2.1 镉胁迫对龙葵叶绿素荧光参数的影响
研究结果表明,Fo不服从正态分布,Fv/Fm方差表现为非齐性,采用wilcoxon秩和检验进行大小比较。qP、NPQ和SPAD值均服从正态分布且方差表现为齐性,采用独立样本t检验进行大小比较。由表1可知,Cd处理下龙葵的Fo显著大于CK(增大了42.7%;P<0.05),Fv/Fm显著小于CK;Cd胁迫下的SPAD值显著小于CK(减小了22.9%;P<0.05);不同处理下,龙葵叶片qP和NPQ均无显著差异(P>0.05)。

表1 镉胁迫对龙葵SPAD值和叶绿素荧光参数的影响 (平均值±标准误差)Table 1 Effects of Cd stress on SPAD and chlorophyll a fluorescence parameters of Solanum nigrum (mean±SE)
2.2 镉胁迫对龙葵光合-光响应的影响
在CK和Cd胁迫下,龙葵光合光响应变化呈现出相似规律(图1)。龙葵的A均随PAR的增大而逐渐增加,最后趋于平缓,达到饱和。当PAR≤ 200 μmol·m-2s-1时,净光合速率几乎呈线性增加,而且CK的净光合速率增幅明显高于Cd处理,这说明龙葵光合作用对PAR的变化非常敏感,而且Cd的敏感程度较CK的低。当PAR为200~800 μmol·m-2s-1时,A的增加变得趋缓。当PAR为800~1 200 μmol·m-2s-1时,A趋于平缓,达到饱和。
研究结果表明,所有光响应参数均服从正态分布。LCP方差表现为非齐性,AQY、Amax和Rd方差表现为齐性。部分光响应曲线拟合获得的LSP远超出阈值,故未对其进行分析。与CK相比,Cd胁迫下龙葵叶片Amax减小了24.9%(P<0.05,表1)。不同处理间龙葵叶片AQY无显著差异。Cd胁迫下龙葵叶片LCP小于CK(差异不显著,P>0.05)。Cd胁迫下龙葵叶片Rd显著大于CK(P<0.05)。
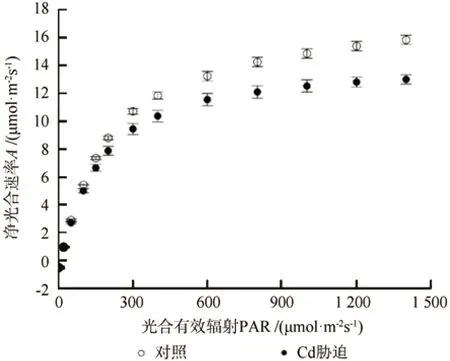
图1 镉胁迫下龙葵叶片的光响应曲线(平均值±标准误差)Fig.1 Light response curves of net photosynthetic rate (A) in Solanum nigrum under Cd stress (mean±SE)

表2 镉胁迫对龙葵光响应参数的影响(平均值±标准误差)Table 2 Effects of Cd stress on photosynthesis-light response parameters of Solanum nigrum (mean±SE)
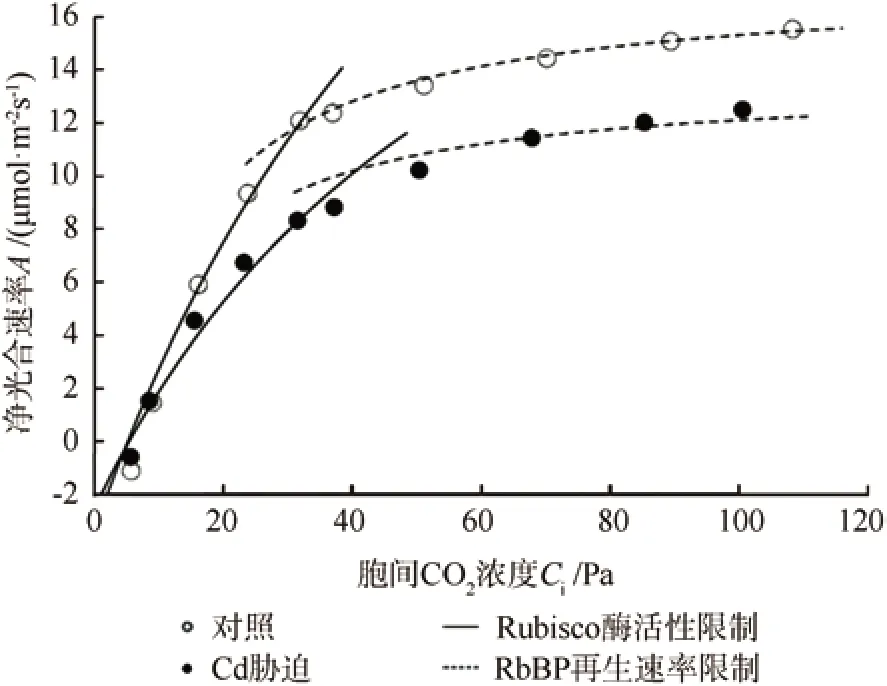
图2 镉胁迫下龙葵叶片CO2响应曲线Fig.2 CO2 response curves of net photosynthetic rate (A) in Solanum nigrum under Cd stress
2.3 镉胁迫对龙葵光合-CO2响应的影响
在CK和Cd胁迫下,龙葵的A均随Ci的增大而逐渐增加,最后趋于平缓(图2)。当Ci≤Ci_CJ时,A随Ci的增大几乎呈线性增加,这说明龙葵光合作用受Rubisco酶活性大小的限制 (实线);而且,CK的A增幅明显高于Cd处理,说明Cd胁迫下A对Ci变化的敏感程度较CK的低。当Ci≥Ci_CJ时,A的增加变得趋缓,说明光合作用受RuBP再生速率大小的限制 (虚线)。采用可决系数 (R2)对FvCB模型的拟合效果进行分析,CK和Cd胁迫CO2响应曲线拟合的R2为0.956 7 ~ 0.991 1。
表3中光合参数均服从正态分布。方差齐性检验结果表明,Ci和Ci_CJ方差表现为非齐性,其它参数的方差表现为齐性。由表3可知,Cd胁迫下龙葵叶片的A、gsc、Ci、Vcmax和Jmax均显著小于CK(P<0.05),减小百分比分别为18.3%、51.2%、4.6%、34.1%和18%。Cd胁迫下的Ci_CJ显著大于CK (增大百分比为46.9%;P<0.05)。光合限制组分量化分析结果(表4)显示,在Cd胁迫下龙葵叶片光合限速因子中,气孔限制(SL)为17.86%,非气孔限制(NSL)为82.14%。

表3 镉胁迫对龙葵叶片光合参数的影响Table 3 Effects of Cd stress on photosynthetic parameters of Solanum nigrum (mean±SE)

表4 镉胁迫下龙葵气孔限制和非气孔限制的比例Table 4 Photosynthesis limitation parameters in Solanum nigrum grown under Cd stress
2.4 镉胁迫对龙葵生物量的影响
分析结果表明,不同处理下龙葵的生物量和SLW均服从正态分布且方差表现为齐性。由表5可知,镉胁迫下龙葵单株生物量、地上部分生物量、根系生物量均显著小于CK(P<0.05),分别减小了46.8%、33.3%和49%。Cd胁迫对龙葵叶片SLW没有显著影响。Cd胁迫下龙葵根系/地上部分比例显著高于CK(高31.5%;P<0.05)。
分析结果表明,不同处理SPAD值与A不服从正态分布,采用Spearman秩相关分析;其余参数均服从正态分布,采用Pearson积矩相关分析。由表6可知,镉胁迫下龙葵单株生物量、地上部分生物量与SPAD值、A、Amax、Vcmax、Jmax呈显著相关关系。镉胁迫下根系生物量与SPAD值、Amax、Vcmax、Jmax呈显著相关关系。

表5 镉胁迫对龙葵生物量和比叶重的影响Table 5 Effects of Cd stress on biomass and specific leaf weight of Solanum nigrum (mean±SE)
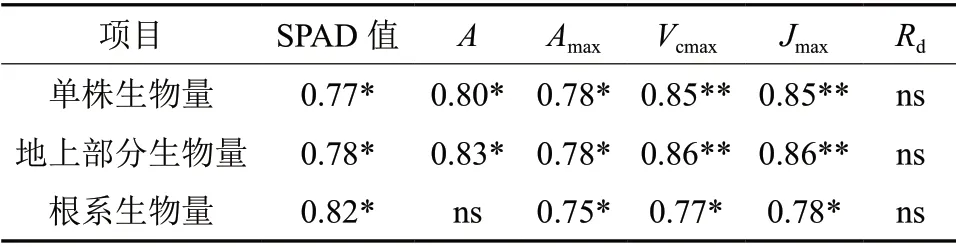
表6 镉胁迫下龙葵生物量与光合参数的相关性分析†Table 6 Correlations between biomass and photosynthetic parameters of Solanum nigrum under Cd stress
由表7可知,龙葵叶片Amax与SPAD、Jmax、Vcmax、Rd呈显著相关关系,A与SPAD、gsc显著相关,SPAD与Jmax、Vcmax、gsc、Rd呈显著相关关系,Vcmax与Jmax、gsc、Rd呈显著相关关系,Jmax与gsc、Rd显著相关。
3 结论与讨论
光合作用是植物重要的生理过程。本研究表明,龙葵叶片光合速率(Amax和A)与生物量呈显著相关关系,这说明光合作用于植株生物量累积密切相关。研究结果表明,Cd胁迫导致龙葵叶片最大净光合速率(Amax)显著下降,说明Cd胁迫导致龙葵叶片光合能力降低,这与Cd胁迫下垂柳[21]和烤烟[8]等的研究结果类似。大量研究表明,重金属胁迫下光合速率的降低可能与光能的吸收、能量转化、电子传递、Rubisco酶羧化反应和叶片内CO2扩散等光合过程有关[8,10,16]。龙葵叶片Amax与SPAD、Jmax、Vcmax和Rd等参数显著相关,说明龙葵光合作用的变化与叶绿素含量、光合电子传递、Rubisco羧化反应、呼吸作用等密切相关。叶绿素是光合光反应中的重要色素,可以反映叶片对光能的吸收和转化能力。Cd胁迫下龙葵叶片的SPAD值显著降低,表明其叶绿素含量下降,说明Cd胁迫可能抑制了龙葵叶片光合色素的合成,这与玉米[22]、美洲黑杨[10]的研究结果类似。这还可能与Cd胁迫下叶绿体数量的减少[23]、矿质元素吸收受阻[24]有关。Cd胁迫下叶绿素含量的下降会直接导致龙葵叶片对光能的吸收和利用能力降低。Cd胁迫下龙葵叶片Rd显著增大,说明叶片可能通过增加呼吸作用以获得更多的能量来应对镉毒害,这与Cd胁迫下玉米[24]的研究结果类似。本研究发现Cd胁迫下龙葵叶片gsc和Ci减小,说明叶片气孔阻力增大,这可能与Cd胁迫下气孔密度和开放气孔数的减少有关[13]。本研究还发现Cd胁迫导致龙葵叶片Vcmax和Jmax降低,这说明Cd毒害对叶片Rubisco酶羧化反应和电子传递过程有一定的抑制作用,这与美洲黑杨[10]的研究结果类似。Vcmax的降低可能是由Cd胁迫下Rubisco酶活性的降低[14]和Rubisco酶基因表达水平的下降[25]而引起的。光合限制组分分析结果表明Cd胁迫下光合速率的限制主要来自于非气孔限制(82.14%),这说明扩散限制与生化限制的作用较大,这与滇苦菜[13]的研究结果类似。

表7 镉胁迫下龙葵光合参数间相关性分析Table 7 Correlations between different photosynthetic parameters of Solanum nigrum under Cd stress
叶绿素荧光参数可以反映重金属对光合机构的胁迫程度。Fo可以反映PSII受到的损害程度。Fv/Fm为PSII最大光能转化效率,反映叶片的光能转化效率。研究结果表明,Cd胁迫下,龙葵叶片Fo显著增大,而Fv/Fm显著下降,说明Cd胁迫导致龙葵叶片PSII受到损伤而降低了对光能的捕获效率,这与Cd胁迫下滇润楠[26]、烤烟[8]和美洲黑杨[10]等植物的研究结果类似。qP可以反映叶片吸收的光能,用于光合CO2固定和光呼吸等光化学过程的能量。NPQ则是反映不被用于光化学反映,而以热能的形式释放的能量,能防止过剩光能对光合机构的伤害。本研究表明,Cd胁迫下龙葵叶片qP、NPQ与对照均无显著差异,说明Cd胁迫对龙葵叶片吸收光能的分配机制没有影响,这与大麦、玉米[12]和滇苦菜[13]等植物的研究结果类似。另一方面,LCP可以反映植物叶片对弱光的利用能力,AQY反映了叶片在弱光环境下的光合能力。惠俊爱等[27]发现Cd胁迫导致玉米叶片LCP升高、AQY降低,但本研究表明Cd胁迫对龙葵叶片LCP和AQY没有显著影响,这说明Cd胁迫下龙葵叶片对弱光的利用能力没有改变。
重金属胁迫对植物光合作用的限制可能与Rubisco酶含量及活性[14]、细胞色素f(cyt f)、ATP合酶等生理指标有关。另一方面,重金属胁迫下植物光合作用的变化还可能与气孔大小及密度[16]、单位面积叶肉细胞暴露在胞间的面积和单位面积叶绿体暴露在胞间的面积等结构特点[28]有关。本研究仅从叶绿素荧光和光合生化特性方面来研究Cd胁迫对龙葵光合作用的影响,未能从生理特性和叶片结构特点等多个方面来阐释植物Cd污染的光合生理机制,未来拟结合生理指标测定、光合生化模型模拟和叶片显微结构观察等手段来研究Cd胁迫对植物光合作用的影响,以全面揭示Cd胁迫对植物光合作用的限制机理。
