不同水位下莼菜叶片气孔及光合特性的相关性分析
2019-08-30张依南管东旭
张依南 张 蔚 田 昆 孙 梅 戴 强 管东旭 李 晖
( 1. 西南林业大学湿地学院,国家高原湿地研究中心,云南 昆明 650233;2. 腾冲北海湿地自然保护区管理局北海管护站,云南 保山 679100)
水生植物通过光合作用和分解作用,在湿地碳循环中扮演着重要角色[1-2]。近年来,由全球气候变化及人为干扰活动等带来的水位变化严重影响着水生植物的生境[3-4]而作为水生生态系统中特殊的初级生产者,水生浮叶植物常在叶片的光合作用及其组织结构上表现出极高的适应性[5-6]。植物叶片是植物进行光合作用的主要场所[7],对植物的健康生长具有重要影响。在环境变化过程中,植物可通过自身表型可塑性适应环境的异质性[8],而植物叶片的可塑性大小则可作为重要的衡量依据之一[9]。植物叶片的气孔和叶脉是其重要的失水和供水系统,二者的相互作用关系影响着植物光合作用能力的大小[10-11]。有研究指出,气孔是植物叶片与外界进行气体交换的重要通道,影响着植物的光合作用、蒸腾作用以及呼吸作用[12-13]。气孔密度、气孔大小等都是衡量蒸腾速率的重要指标[14-15]。双子叶植物气孔复合体是由两个保卫细胞和中间的微孔组成的[16-17]。蒲公英(Taraxacum mongolicum)、火绒草(Leontopodium leontopodioides)和美丽风毛菊(Saussurea pulchra)等菊科植物在长期的进化过程中,气孔密度随着温度增加而呈现增加趋势[18],研究指出,水葱的气孔对CO2浓度的变化敏感[19-20],海拔差异显著,但水生浮叶植物对水位变化的气孔适应性研究仍然十分缺乏。叶脉是为植物叶片运输水分的重要通道,是植物重要的供水系统,它与蒸腾作用的失水形成鲜明的对比,是植物叶片生长的重要功能性结构。张亚等[21]研究指出,基部被子植物气孔密度与叶脉密度成显著正相关关系,二者属于关联进化,与陆生植物不同的是,水生浮叶植物的叶片漂浮于水面,其特殊生境将如何决定叶脉密度与气孔密度之间的关系,仍有待进一步研究。
莼菜(Brasenia schreberi)属水生浮叶植物,1999年被列为国家Ⅰ级重点保护野生植物[22]。因其有性繁殖能力弱,对土壤及水环境要求高,其原生种群较少[23]而极度濒危。莼菜在国内主要分布于长江中下游地区的江浙一带[24],在生物多样性丰富的云南,野外仅在腾冲北海湿地发现其天然分布[25]。由于莼菜富含莼菜多糖,具有重要的药用价值,其嫩叶可做水生蔬菜食用,具有较高的经济利用价值[26]。在湿地保护与利用矛盾的平衡中,莼菜的利用成为了湿地资源合理利用的范例,如何有效保护莼菜这一珍稀濒危湿地植物,科学恢复其生境是许多学者关注的科学问题[27]。但国内对莼菜的保护利用研究多注重其药用价值及经济价值,近年来,由于气候变化及人为干扰的影响,水位及水质的变化严重影响到莼菜原生种群资源的保护[28]。本研究从浮叶植物莼菜的叶片结构性状及其对水位变化的适应性机制开展研究,以期为濒危种莼菜原生种群的保护和繁育提供科学参考,同时也为其经济栽培利用及湿地生态补水工程的实施提供参考依据。
1 研究区概况
研究地位于云南省保山市腾冲县的北海湿地自然保护区,地处东经 98°30′30″~98°34′50″,北纬 25°6′42″~25°8′38″,属高黎贡山西坡的浅切割中山地貌,为典型的亚热带火山熔岩堰塞湖湿地,是伊洛瓦底江水系上游的重要湖泊,其主要补给水源为大气降水和四周汇水面山带来的汇集水源。境内年平均气温14 ℃,多年平均降雨量1 740 mm[29],降水主要集中在5—9月。经筑坝补水后,水域内平均水深约为6 m,境内地表径流面积为16.29 km2。
2 材料与方法
2.1 研究材料
研究材料采自腾冲北海湿地自然保护区境内,在野外选择3种水位生境,对照水位(CK)为1.4 m、低水位为(LWL)1.0 m和深水位为(DWL)1.8 m进行研究,以立体空间变化代替时间演变的方法,探索大气降水减少带来的水位下降和人工补水带来的水位上升所引起的水位变化对莼菜的影响。
2.2 研究方法
2.2.1 光合气体交换参数测定方法
于2018年7—9月中旬,选取晴朗天气,在9:00—11:00,使用LI-6400便携式光合仪(Li-COR,USA)测定莼菜健康植株叶片的净光合速率(Pn)、气孔导度(Gs)、胞间 CO2浓度(Ci)、蒸腾速率(Tr)每个指标重复10次。光合参数测定前,将叶片提出水面,擦净叶片上下表面,用光合测定仪的探测头夹住叶片一端,将另一端保持与水面接触,尽可能维持其原位生境,然后用1 200 μmol/(m2·s)及 1 800 μmol/(m2·s)的光强进行光诱导,以使其具有稳定的最大气孔导度。维持光强为 1 000 μmol/(m2·s),并使叶片适应3~5 min后,记录叶片的光合生理参数。测定时,为保证测定数据的稳定性与准确性,维持叶室内的温度、相对湿度、压强和CO2浓度分别为(31.79±2.33)℃、67.61%±6.99%、(82.76±0.01)kPa和(384.69±8.89)μmol/mol。按公式(1)~(2)计算叶片瞬时水分利用效率(WUE)和光能利用效率(LUE)。

采用光合—光响应曲线自动测量程序分别记
录光照强度为2 200、2 000、1 800、1 600、1 400、1 200、1 000、600、200、100、50、0 μmol/(m2·s)下的光合速率。测定前,叶片在1 800 μmol/(m2·s)光强下进行充分的光诱导,每个光强测定时间为120~180 s。测定时,叶室内的温度、相对湿度、压强和CO2浓度分别为(33.45±2.42)℃、57.29%±7.30%、(82.05±0.19)kPa和(388.13±7.58)μmol/(m2·s)。根据光合—光响应曲线拟合的非直角双曲线模型[30-31]求得光补偿点(LCP)、光饱和点(LSP)、暗呼吸速率(Rd)及最大净光合速率(An)。
测量结束后将植株整株捞起,将叶片保鲜密封后迅速带回验室进行室内分析。
2.2.2 气孔特征观测方法
气孔特征参考指甲油涂抹印迹法[32]进行测量。将带回的叶片清理干净后,在上下表皮分别涂抹上一层透明指甲油,待晾干后,用镊子轻轻剥取指甲油印迹,将其置放在备好的载玻片上,制成临时玻片,在莱卡光学显微镜(Leica Corp,Germany)下观察,在40倍物镜下用显微镜装备的照相机拍取照片,照片大小为0.31 mm×0.24 mm,保存为JPEG格式(如图1)。用Image J软件分别测量气孔的长度、宽度、面积及气孔孔隙长,并统计每张印迹照片中的气孔个数,计算气孔密度(SD)。气孔密度用单位叶面积内气孔的个数来计算。从每个玻片对角线选取10个视野,每个视野3张照片,每片叶片共90张照片。

图 1 气孔制片图Fig. 1 Stomatal density picture
2.2.3 叶脉特征观测方法
选取野外带回的健康叶片,洗净后在每个叶片剪取适宜大小的叶片,用质量分数为5%的氢氧化钠溶液浸泡7 d,每天更换1次溶液。待叶片用蒸馏水清洗3遍,并浸泡0.5 h,加入比例为1∶1的次氯酸钠溶液,浸泡12 h,清水去除,浸泡0.5 h后依次用体积分数为95%、70%、50%、20%、10%共5个浓度梯度的乙醇浸泡,每个浓度梯度浸泡0.5 h,然后用清水浸泡待测。将处理后的叶片吸至载玻片上,用配置好的质量分数为1%甲基蓝溶液进行染色,用吸水纸从玻片的一侧吸去多余的水和浮色,制成临时玻片,将其置于莱卡光学显微镜20倍物镜下观察并用显微镜装备的照相机拍取照片,照片大小为0.63 mm×0.47 mm,保存为JPEG格式(如图2)。用Image J软件测量叶脉的总长度,计算叶脉密度(VD)。叶脉密度用单位面积内叶脉的长度来表示。从每个玻片对角线选取10个视野,每个视野3张照片,每片叶片共90张照片。

图 2 叶脉制片图Fig. 2 Vein density picture
2.3 数据处理与统计分析
利用SPSS 16.0软件,以水位梯度为单因素,对各指标进行单因素方差分析,然后对不同水位梯度上的叶片结构指标(SD、VD)与光合指标(Pn、Gs、Ci、Tr)之间以及SD与VD之间进行Pearson相关性分析,阐明叶片结构对光合作用的影响,探讨水生浮叶植物对光合的适应策略。
3 结果与分析
3.1 不同水位梯度下莼菜叶片解剖学指标的适应特征
由表1可知,在不同水位梯度下,莼菜叶片的各解剖学指标间均具有显著差异(P<0.05)。除SD外,LWL莼菜气孔各项指标均大于对照(CK)水位,而DWL的各项气孔特征均小于CK,气孔长(SL)、气孔宽(SW)、气孔面积(SA)和气孔孔隙长(SPL)均呈现在DWL具有更小的数值。而SD和VD则刚好相反,最高值出现在LWL,最大值出现在DWL。

表 1 不同水位梯度下莼菜解剖特征值及差异比较Table 1 Comparison of anatomical characteristics and differences of B. schreberi under different water level gradients
3.2 不同水位梯度下莼菜的光合生理适应特征
由表2可知,在不同水位梯度下,莼菜叶片的各光合气体交换参数间均具有显著差异(P<0.05)。LWL光强值高,这对莼菜的Pn、Tr和LUE起到了促进作用,而DWL光强较弱,莼菜的WUE较高。而不同水位条件下,莼菜的Gs和Ci均处于较低水平。

表 2 不同水位梯度下莼菜光合气体交换参数值及差异比较Table 2 Comparison of photosynthetic gas exchange parameter values and differences of B. schreberi under different water level gradient
光响应曲线是反映物种对光合有效辐射的利用能力的曲线表现方式。在不同水位下,即使是同一个物种,也同样会具有不同的表现形式。本研究通过对不同水位莼菜测定光响应曲线,探究其对光合有效辐射的利用能力。
由图3可知,不同水位莼菜对光合有效辐射的利用能力产生了显著差异(P<0.05)。且不同水位的莼菜LCP和LSP均有所不同,LWL、CK、DWL的 LSP 分别为 1 432.93、1 701.33、1 044.56 μmol/(m2·s),LCP 分别为 41.07、38.13、70.40 μmol/( m2·s),Pn分 别 为 15.93、 15.11、 10.77 μmol/(m2·s)。由此可见,Pn在DWL水平上最小,表明DWL的莼菜光合能力潜力较小,LSP降低在DWL水平上呈现最小值,则说明DWL莼菜对强光的利用能力最弱。

图 3 不同水位下莼菜的光响应曲线Fig. 3 Pn-PAR curves of B. schreberi under different water levels
3.3 不同水位梯度下莼菜的解剖学特征与光合作用的相关性分析
叶片是莼菜进行光合作用的主要部位,而气孔和叶脉是植物进行水分输送的重要器官,气孔和叶脉对水位变化的适应性影响着莼菜的光合能力和养分积累。本研究对莼菜的气孔特征及叶脉密度等与光合气体交换参数进行相关性分析(表3~5),结果表明CK与DWL莼菜SD与Pn、Gs及Tr间相关性较好,呈显著线性负相关(P<0.05),而LWL莼菜SD与各光合气体参数间相关性较差,水位梯度与Ci间不具有线性相关性。SD的增加会增加水分蒸腾,而水分蒸腾过多又会抑制植物的气孔导度,Gs的下降则又会抑制植物的光合作用。

表 3 CK水位下气孔性状、叶脉密度及合气体交换参数值间的相关性Table 3 The correlations between the stomatal traits, vein denisy and the gas exchange parameters of CK

表 4 低水位下气孔性状、叶脉密度及合气体交换参数值间的相关性Table 4 The correlations between the stomatal traits, vein denisy and the gas exchange parameters of LWL
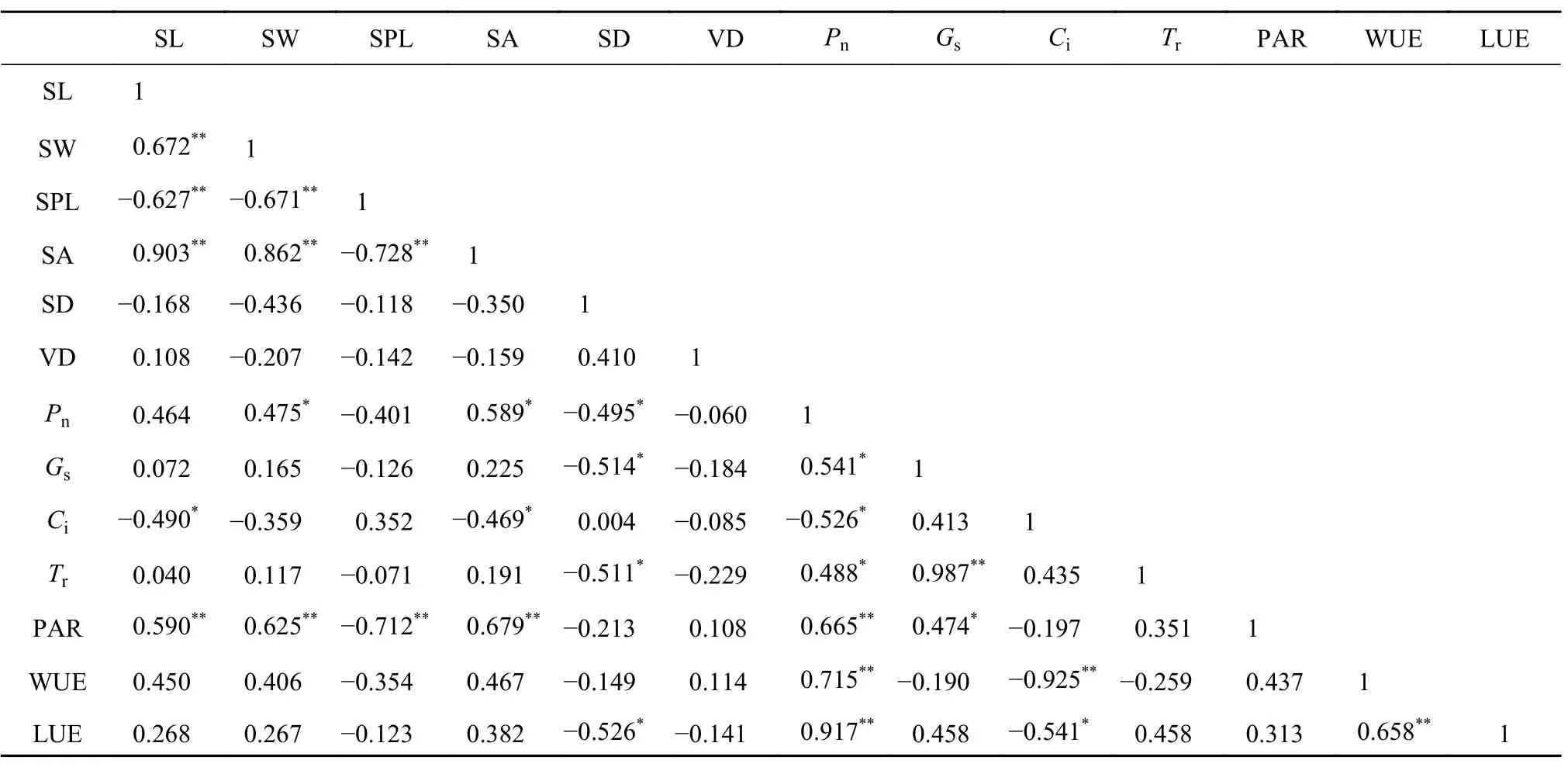
表 5 深水位下气孔性状、叶脉密度及合气体交换参数值间的相关性Table 5 The correlations between the stomatal traits, vein denisy and the gas exchange parameters of DWL
由表3~5可知,水生浮叶植物莼菜的VD与光合气体交换参数间均不存在显著相关性,与水位变化无较大影响,莼菜的VD对其叶片光合能力的影响较小。VD和气孔特征间不存在协同变异。经综合分析发现,莼菜的VD和SD间存在不显著正相关关系,而与SPL和SA间存在不显著负相关关系。但从不同水位上莼菜VD与气孔特征间的线性拟合关系上看,均无显著相关性,说明水位变化对二者间相关性关系的影响较小。
4 结论与讨论
4.1 水生植物叶片结构性状与光合作用之间的关系
莼菜属浮叶根生的浮叶植物,叶片是其进行光合作用的重要部位。本研究发现,莼菜的气孔结构仅存在于其叶片上表皮,对于全植株唯一与空气接触的重要部位,莼菜叶片气孔对水位变化的适应性将对植株的光合作用及其健康生长产生巨大影响。莼菜叶片的气孔特征与光合作用密切相关。作为植物的失水系统,气孔对植物的光合作用能力起到重要作用[33-34]。植物气孔特征对环境的适应,主要通过SD和气孔大小的协调变化实现。SD增加,总SA增加,从而提高了植物失水速度,对植物的光合速率起到了抑制作用。对于水生浮叶植物而言,气孔多分布在植物进行气体交换的上表皮,气孔特征的变化,影响着浮叶植物的气体交换速率,从而影响植物的光合作用及其适应环境的能力,若水位上涨淹没漂浮于水面的叶片,导致其气孔遭到水体堵塞,将破坏植物的这种协调机制,影响湿地植物的健康生长。同时,DWL植物叶片面积较小,叶片气孔采取减小SA和SL,增加SD,从而增加气孔灵活性,适应深水位环境以维持自身生长。LWL莼菜的SD与光合气体交换参数间不具有线性关系,可能与LWL莼菜数量较多,单片叶片的光合作用并不影响整株植物的光合物质积累有关。这可能与叶片面积的变化有关。研究发现,低水位莼菜拥有更宽的叶片伸展空间,而深水位莼菜将更多的光合物质投入到茎的伸长从而限制了叶片面积的扩增。
在封闭半封闭的湖泊湿地中,水位的变化将对植物的生长具有重要的影响。LWL水体的光照强度更强,对植物的光合作用有可能会起到促进作用,而DWL植物接收的光强较小,有可能会限制其光合作用。本研究材料莼菜属于湿地浮叶植物,水位的变化对其光合生理适应将产生重要影响。
叶脉特征的变化能在一定程度上反映植物的适应能力大小[18]。同样是湿地植物的芨芨草,在土壤水分变化的过程中,通过自身叶脉特征的变化合理进行资源的优化配置,从而更好地应对生境中资源的异质性分布[35]。秦茜等[36]研究发现,耐旱性强的甘蔗品种具有更高的VD,而甘蔗VD与Pn呈显著正相关关系,这与本研究结果不同,表明叶脉特征对光合的影响作用在陆生植物和水生植物之间存在着显著差异。水生浮叶植物莼菜的VD与光合气体交换参数间不存在相关性关系。这可能与浮叶植物的叶片特性有关,浮叶植物莼菜的下表皮具有较厚的胶质,并且在其茎和叶片下表皮均具有根毛,具有供水功能,因此削弱了叶脉对叶片供水作用,而更加突出其抵抗水流带来的风浪及支撑叶片伸展的作用。
4.2 水生植物的水位变化适应策略
叶脉是植物叶片光合作用的主要供水系统,VD的增加,能够促进植物叶片的水分和无机盐运输,并增强植物叶片的抗风险能力[37]。段贝贝等[38]研究发现,刺槐VD与SD呈显著正相关关系。莼菜VD与SD呈现不显著的相关性,反映了莼菜具有与刺槐不同的环境适应策略,通过保持植物叶含水量的动态平衡,从而更合理分配和利用叶性状间的资源。宋丽清等[39]研究发现,高粱叶片较紫苏具有更大的VD,导水能力更强,从而具有更高的Tr,但SD却较低。其研究结果显示VD与SD可能不存在严格的协同变异关系,本研究结果与其一致,莼菜VD与SD间也不存在严格的协同变异关系,这可能与莼菜的气孔分布有关,高粱和紫苏均为陆生植物,叶片的上下表皮均有气孔分布,所以气孔的密度与叶脉的密度不存在协同变异关系,而水生浮叶植物莼菜的气孔只分布于叶片的上表皮,但莼菜下表皮的根毛结构能够有效的促进浮叶植物进行水分吸收,从而支持光合作用和植物生长所需的水分,以此来降低叶脉的导水压力,这就导致莼菜SD和VD之间不能进行协同变异。莼菜的VD的增加促进了莼菜叶片的伸展性生长,增加了叶片的整体面积,从而影响了SD与VD之间的协同变异关系。但从水位变化上来看,水位的加深或者变浅均未能使得SD与VD之间的相关性得到增强,说明水位变化对莼菜SD与VD之间的相关性影响较小。
水生植物莼菜在光合作用过程中的气孔特征与光合之间的协同变异能较好的促进植物的健康生长;但同时,也存在着因某一因素生长受限,对另一因素起到协同限制的风险。原生莼菜种群有性繁衍能力低,抗风险能力弱,LWL为浮叶植物提供了更多的光照资源,促使其根茎横向生长,叶片数量大量增加,而DWL则促使其根茎垂直生长,若根茎生长未能跟上水位变化的速率,则可能会致使浮叶植物上表皮气孔受到水体压力过强,造成堵塞现象,限制其气体交换速率,从而影响其光合作用,导致植物叶片的不健康生长,甚至会影响到整株植物的存亡。
因此,对于恢复湿地面积的生态补水应更多地考虑其补水策略,通过多次水位调节的科学补水,使浮叶植物逐渐适应其变化,既起到恢复湿地面积的作用又可更好地保护水生植物,做到真正恢复湿地生态系统功能。湿地植物是湿地生态系统的第一生产者,具有重要的固碳、吸附污染物等多种作用,湿地补水应更多的考虑湿地植物群落的影响,而湿地浮叶植物群落对水位的变化更加敏感,因此在湿地补水前,应更深层次的考虑湿地浮叶植物群落的耐受性,将其作为补水策略中重要的因素去考虑。
[ 参 考 文 献 ]
[1]田昆, 徐写秋. 高原湿地的碳循环 [J]. 生命世界,2015(2): 38-41.
[2]郭绪虎, 田昆, 肖德荣, 等. 滇西北高原纳帕海湿地湖滨带优势植物凋落物分解特征研究 [J]. 生态科学,2013, 32(2): 200-205.
[3]Yu L F, Yu D. Responses of the threatened aquatic plant Ottelia alismoides to water level fluctuations [J].Fundamental and Applied Limnology, 2009, 174(4):295-300.
[4]Davies P. Algal beds and threatened aquatic fauna in Great Lake: Current status, responses to lake level and management recommendations [J]. Library Home,2001.
[5]Jiang M, Kadono Y. Seasonal growth and reproductive ecology of two threatened aquatic macrophytes, Blyxa aubertii and B. echinosperma (Hydrocharitaceae), in irrigation ponds of south-western Japan [J]. Ecological Research, 2001, 16(2): 249-256.
[6]Leyer I. Predicting plant species’ responses to river regulation: the role of water level fluctuations [J].Journal of Applied Ecology, 2005, 42(2): 239-250.
[7]程建峰, 陈根云, 沈允钢. 植物叶片特征与光合性能的关系 [J]. 中国生态农业学报, 2012, 20(4): 466-473.
[8]蔡志全, 齐欣, 曹坤芳. 7种热带雨林树苗叶片气孔特征及其可塑性对不同光照强度的响应 [J]. 应用生态学报, 2004, 15(2): 201-204.
[9]何春霞, 李吉跃, 孟平, 等. 4种高大树木的叶片性状及WUE随树高的变化 [J]. 生态学报, 2013, 33(18):5644-5654.
[10]单提波, 徐正进. 不同气孔密度水稻叶片光合及叶绿素荧光参数的日变化特征 [J]. 沈阳农业大学学报,2015, 46(2): 129-134.
[11]彭莉霞, 黄东兵, 吴德, 等. 竹节树幼苗的光响应特性及其曲线最适模型选择 [J]. 西南林业大学学报(自然科学), 2018, 38(4): 1-5.
[12]龚容, 高琼. 叶片结构的水力学特性对植物生理功能影响的研究进展 [J]. 植物生态学报, 2015, 39(3):300-308.
[13]潘莹萍, 陈亚鹏. 叶片水力性状研究进展 [J]. 生态学杂志, 2014, 33(10): 2834-2841.
[14]孙梅, 田昆, 张贇, 等. 植物叶片功能性状及其环境适应研究 [J]. 植物科学学报, 2017, 35(6): 940-949.
[15]冯春慧, 何照棚, 田昆, 等. 不同海拔生长的水葱功能适应性对比研究 [J]. 西南林业大学学报(自然科学),2019, 39(1): 167-172.
[16]Xu Z, Zhou G. Responses of leaf stomatal density to water status and its relationship with photosynthesis in a grass [J]. Journal of Experimental Botany, 2008,59(12): 3317-3325.
[17]孙会婷, 江莎, 刘婧敏, 等. 青藏高原不同海拔3种菊科植物叶片结构变化及其生态适应性 [J]. 生态学报,2016, 36(6): 1559-1570.
[18]王宏亮, 郭思义, 王棚涛, 等. 植物气孔发育机制研究进展 [J]. 植物学报, 2018, 53(2): 164-174.
[19]刘国栋. 气候变化对高原湿地优势植物生理生态学特性的影响研究[D]. 重庆: 重庆大学, 2016.
[20]许俊萍, 田昆, 孙梅, 等. 水葱构件生长对大气CO2浓度升高的响应 [J]. 西南林业大学学报, 2016, 36(5):84-88.
[21]张亚, 杨石建, 孙梅, 等. 基部被子植物气孔性状与叶脉密度的关联进化 [J]. 植物科学学报, 2014, 32(4):320-328.
[22]方元平, 刘胜祥, 汪正祥, 等. 七姊妹山自然保护区野生保护植物优先保护定量研究 [J]. 西北植物学报,2007, 27(2): 348-355.
[23]董元火, 雷刚, 刘红艳. 利川绝迹莼菜种群原因分析 [J]. 安徽农业科学, 2010, 38(11): 6047-6048.
[24]胡美华, 金昌林, 杨新琴, 等. 浙江省水生蔬菜产业现状及发展对策 [J]. 浙江农业科学, 2012, 53(3):269-272, 275.
[25]刘晓达. 腾冲北海湿地湿生植物资源与可持续性研究[D]. 北京: 中央民族大学, 2017.
[26]吴会会, 刘乐承. 湖北利川莼菜资源的开发利用与保护 [J]. 长江蔬菜, 2013(18): 15-17.
[27]朱红莲, 柯卫东, 刘玉平, 等. 四川雷波马湖莼菜生产及加工情况考察 [J]. 长江蔬菜, 2013(6): 28-29.
[28]王昌全, 李焕秀, 彭国华, 等. 土壤及水质条件与莼菜生长的关系 [J]. 四川农业大学学报, 2000, 18(3):265-268.
[29]冯春红. 云南省腾冲北海湿地径流区水文特性分析 [J]. 中国农村水利水电, 2013(10): 46-48.
[30]Thornley J H M. Mathematical Models in Plant Physiology[M]. London: Academic Press, 1976.
[31]贺燕燕, 王朝英, 袁中勋, 等. 三峡库区消落带不同水淹强度下池杉与落羽杉的光合生理特性 [J]. 生态学报, 2018, 38(8): 2722-2731.
[32]叶春秀, 庄振刚, 李有忠, 等. 棉花叶片气孔制片方法比较及改良 [J]. 分子植物育种, 2014, 12(3): 543-546.
[33]周丽, 陈诗, 蔡年辉, 等. 云南松不同种群4年生苗木光合特性比较 [J]. 西南林业大学学报(自然科学),2018, 38(3): 71-79.
[34]方怡然, 蔡金桓, 李洁, 等. 加勒比松纯林混交改造阔叶树种叶片的养分和光合特性研究 [J]. 西南林业大学学报(自然科学), 2018, 38(2): 30-36.
[35]韩玲, 赵成章, 徐婷, 等. 不同土壤水分条件下洪泛平原湿地芨芨草叶片厚度与叶脉性状的关系 [J]. 植物生态学报, 2017, 41(5): 529-538.
[36]秦茜, 朱俊杰, 关心怡, 等. 7个甘蔗品种叶片解剖结构特征与光合能力和耐旱性的关联 [J]. 植物生理学报, 2017, 53(4): 705-712.
[37]刘希凤. 植物叶片拓扑结构与其力学性能关系研究[D]. 广州: 华南理工大学, 2010.
[38]段贝贝, 赵成章, 徐婷, 等. 兰州北山不同坡向刺槐叶脉密度与气孔性状的关联性分析 [J]. 植物生态学报,2016, 40(12): 1289-1297.
[39]宋丽清, 胡春梅, 侯喜林, 等. 高粱、紫苏叶脉密度与光合特性的关系 [J]. 植物学报, 2015, 50(1): 100-106.
