温度波动对冷却猪肉品质的影响
2019-08-28魏里朋何承云康壮丽马汉军王正荣赵圣明朱明明
魏里朋,何承云,康壮丽,鲁 飞,马汉军,王正荣,赵圣明,朱明明
(河南科技学院食品学院,河南新乡 453003)
生猪屠宰后在常温下存放极易腐败,将屠宰后的猪胴体经过检疫合格后24 h内快速降温到0~4 ℃,并且在后续处理环节保持该温度的鲜肉称为冷却肉[1]。与冷冻肉相比,冷却肉不经过低温和形成冰晶,内部组织结构不会受到严重的破坏,所以冷却肉的汁液和营养物质不会过多流失,口感和风味受到的影响较小[2];与热鲜肉相比,经过冷却处理后,微生物的生长可以得到很好的控制,从而延长保质期,因此,冷却肉以其新鲜、味美、营养、卫生等优点受到消费者的喜爱[3-4]。
消费者在购买过程中最直接的判断标准就是肉色,而色泽与pH、剪切力、储藏损失等指标密切相关[5]。冷藏温度在运输和保藏过程中会发生波动,影响冷却肉品质。目前对于冷却肉的保藏主要从微生物、保鲜剂、包装技术、冷藏温度等方面进行研究[6]。而温度波动对于冷却肉的色泽、pH等品质所产生的影响是不可忽视的,但关于温度波动对冷却肉品质影响的报道相对较少。
因此,本试验主要研究冷藏条件下温度波动对冷却肉品质和保水性的影响,为冷却肉在冷藏过程中的品质控制提供理论支持。
1 材料和方法
1.1 材料与仪器
猪背最长肌 购于众品集团。
CR-400色差计 日本美能达公司;PQ00l台式NMR分析仪 上海纽迈电子有限公司;C-LM4数显式肌肉嫩度仪 东北农业大学工程学院;电热式水浴锅 山东诸城市新旭东机械有限公司;AUY120电子天平 日本岛津公司;pH计 梅特勒-托利多仪器(上海)有限公司。
1.2 实验方法
1.2.1 猪背最长肌的冷藏 宰后冷却24 h的猪背最长肌分2个批次随机取回,共有样本120块(60头猪),使用保鲜膜包裹后放置加有冰袋的泡沫保温箱中,1 h内运回实验室。将猪背最长肌切成5 cm×5 cm×4 cm的肉块,使用托盘包装,分别在4 ℃,0和4 ℃(记0~4 ℃)各2 h,4和8 ℃(记4~8 ℃)各2 h条件下冷藏0、24、48和72 h,取样待用。
1.2.2 pH测定 取猪背最长肌5 g,将肉样剪成碎末放于小烧杯中加入45 mL蒸馏水,用匀浆机混匀后在室温静置10 min左右,测定pH。每组测定5次。
1.2.3 冷藏损失率 冷藏损失率按式(1)计算,每组测定3次。
冷藏损失率(%)=(m1-m2)×100/m1
式(1)
式中:m1为冷藏前猪背最长肌质量;m2冷藏后猪背最长肌质量。
1.2.4 蒸煮损失率 将真空包装的猪背最长肌在75 ℃水中煮制30 min至中心温度72 ℃,捞出放入流水中冷却,至中心温度降至室温。蒸煮损失率按式(2)计算,每组测定3次。
蒸煮损失率(%)=(m1-m2)×100/m1
式(2)
式中:m1为蒸煮前猪背最长肌质量;m2为蒸煮后猪背最长肌质量。
1.2.5 色差的测定方法 使用色差计对猪背最长肌表面不同部位进行测定,标准白色比色板为L*=96.86,a*=-0.15,b*=1.87。其中L*代表亮度值,a*代表红度值,b*代表黄度值,每组测定5次。
1.2.6 剪切力的测定 使用肌肉嫩度仪来测定生猪背最长肌的剪切力。先顺着肌原纤维方向切成3 cm×1.5 cm×1.5 cm的长方体肉柱。用剪切仪沿垂直于肌原纤维的方向剪切肉柱,记下剪切力值(N),每组测定5次。
1.2.7 NMR自旋-自旋驰豫时间(T2)测量 称取重量为2 g左右的生猪背最长肌放入直径为15 mm的核磁管后放入NMR分析仪中。测量温度为32 ℃,质子共振频率为22.6 MHz。参数如下:τ-值为(90 °脉冲和180 °脉冲之间的时间)为200 μs。重复扫描32次,重复间隔时间为6.5 s,得到12000个回波,每个测试至少4次。
1.3 数据处理
本实验所有处理重复4次,每个值是平均值±SD。应用软件SPSS v.18.0进行统计分析,使用单因素方差分析(ANOVA)的方法对数据进行分析,当p<0.05时,组间存在显著差异。
2 结果与分析
2.1 温度波动对冷却猪背最长肌pH的影响
pH是反映屠宰后肌糖原酵解速率的重要指标,也是判断生理正常肉或异常肉(PSE或DFD肉)的依据[7]。通常冷却肉的一级鲜度pH为5.8~6.2,二极鲜度pH为6.3~6.6,而变质肉的pH则达到6.7以上[8]。温度波动对不同冷藏时间冷却猪背最长肌pH的影响如图1所示。在冷藏初期,肌肉的组织细胞仍然进行新陈代谢,肌糖原发生酵解,ATP分解,分别产生乳酸、磷酸等酸性物质,导致肌肉的pH逐渐降低[9]。随着时间的推移,pH整体呈现上升趋势,与李苗云等[10]的研究结果一致。这是因为肉中的蛋白质在微生物和酶的作用下被分解为小分子的氨和胺类化合物等碱性物质[11-12]。但在相同冷藏时间下,0~4 ℃的pH最小,主要原因是随着温度的升高,氢键、疏水作用等化学键被破坏,蛋白质的立体结构崩塌,其酸性基团减少[13],且温度为4 ℃时,温度恒定,微生物的生长与酶活性都能稳定的发挥作用,使得蛋白质被分解为小分子的氨和胺类化合物等碱性物质[11],造成pH升高。在0~4 ℃温度波动区间时可以在一定程度上抑制微生物生长[14],使碱性物质的生成相对较少,pH的上升程度也相对较小。而在4~8 ℃这个温度波动区间中,微生物生长加快,在相同时间内碱性物质生成较多。另外在0~4 ℃的温度波动中,随着时间的增长,pH显著升高(p<0.05)。

图1 温度波动对冷却猪背最长肌pH的影响
2.2 温度波动对冷却猪背最长肌冷藏损失率的影响
由图2可知,不同温度波动对猪背最长肌冷藏损失率影响显著(p<0.05)。冷藏损失率随着温度升高而显著增大(p<0.05),随着冷藏时间延长,冷藏损失率也显著增大(p<0.05),因为肌肉中的蛋白质分子所带静电荷之间相互吸引可以将水分纳入蛋白质高分子网状结构的立体空间中,随着冷藏时间的延长,蛋白质降解,水分发生流失,肌肉产生冷藏损失。Bowker等[15]报道了随着冷藏时间的延长,肉样滴水损失显著增加。而在相同的冷藏时间中,0~4 ℃温度波动区间的冷藏损失率相比于4 ℃和4~8 ℃的波动区间的冷藏损失率较小。可能因为随着温度的增加和冷藏时间的延长,微生物繁殖速度较快,酶的活性增大[16],导致蛋白质降解的程度也逐渐增大,所能吸附的水分也相应减少,水分流失增大。
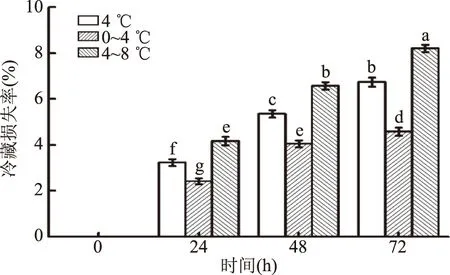
图2 温度波动对冷却猪背最长肌冷藏损失率的影响
2.3 温度波动对冷却猪背最长肌蒸煮损失率的影响
肌肉的持水性是衡量冷却肉品质的主要指标。由图3可知,蒸煮损失率随着温度波动的升高显著增加(p<0.05),同时也随冷藏时间的延长而显著增加(p<0.05)。在相同的冷藏时间下,0~4 ℃的波动区间的蒸煮损失率最小(p<0.05)。温度越高,对肌纤维的影响越大;冷藏时间越长,肌纤维的变化越明显,肌纤维横向收缩是造成汁液流失通道变宽,保水能力下降的主要原因[17]。延长冷藏时间或者升高冷藏温度,肌肉自身的蛋白质结构产生了较大的破坏,造成蒸煮损失率增加。
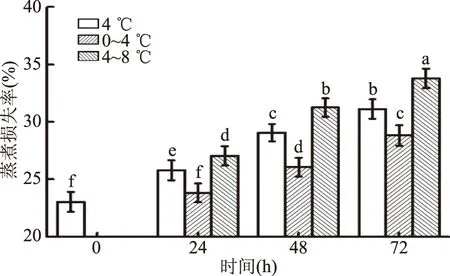
图3 温度波动对冷却猪背最长肌蒸煮损失率的影响
2.4 温度波动对冷却猪背最长肌色差的影响
由表1可知,随着时间的延长,0~4 ℃冷藏下,L*值的差异并不显著(p>0.05);4 ℃冷藏下,24 h时显著降低(p<0.05),48 h以后显著升高(p<0.05),而48和72 h时差异不显著(p>0.05);4~8 ℃冷藏,L*值在0~48 h之间的差异不显著(p>0.05),72 h时显著降低(p<0.05)。随着冷藏温度的升高,L*值变化加快,因为冷藏温度较高,肌肉氧化速度加快[18],造成L*值在4~8 ℃下72 h时显著降低(p<0.05)。0~4 ℃冷藏,a*值在24和48 h时显著升高(p<0.05),且差异不显著(p>0.05),72 h时显著降低(p<0.05),但仍显著高于0 h(p<0.05);4 ℃冷藏,24和48 h时a*值显著升高(p<0.05),但24 h显著高于48 h,72 h时显著降低(p<0.05),与0 h差异不显著(p<0.05);4~8 ℃冷藏,a*值的变化趋势与4 ℃冷藏时一致,但72 h时a*值更低。肌肉的a*值与高铁肌红蛋白含量相关[19],较高温度下,a*值变化较快,这是因为肌红蛋白与氧气结合生成氧合肌红蛋白和高铁肌红蛋白的速率加快[20-21]。胡胜杰等[22]研究表明a*值在冷藏12 h时显著(p<0.05)增大,24~36 h时变化不显著,48 h以后显著减小(p<0.05)。随着冷藏时间的延长,b*值显著升高(p<0.05)。高温加速b*值的升高,因为肌肉表面微生物代谢加速,硫化肌红蛋白含量增加,造成b*值升高,肌肉品质下降[23]。

表1 温度波动对冷却猪背最长肌色差的影响(L*,a*,b*值)
2.5 温度波动对冷却猪背最长肌剪切力的影响
不同冷藏时间和温度波动影响冷却肉的剪切力(图4)。随着冷藏时间的延长剪切力显著下降(p<0.05),0~4 ℃冷藏,剪切力在0和24 h时差异不显著(p>0.05),在48和72 h时显著降低(p<0.05),且两者之间差异不显著(p>0.05);4 ℃冷藏,剪切力在0 h最大,在24、48和72 h时显著下降(p<0.05),且24和48 h时差异不显著(p>0.05),72 h时最小;4~8 ℃冷藏,0、24、48和72 h时差异显著(p<0.05),随着时间的延长,剪切力显著下降(p<0.05)。随着冷藏温度的升高,剪切力降低速度加快。因为在较高温度下,钙激活蛋白酶活性增加,加速了肌纤维中起连接、支架作用蛋白质的水解,引起细胞结构的弱化,造成肌肉松软,促使肉嫩度的增加[24-25]。延长冷藏时间,钙激活蛋白酶作用时间增加,也能够降低肌肉的剪切力[26]。
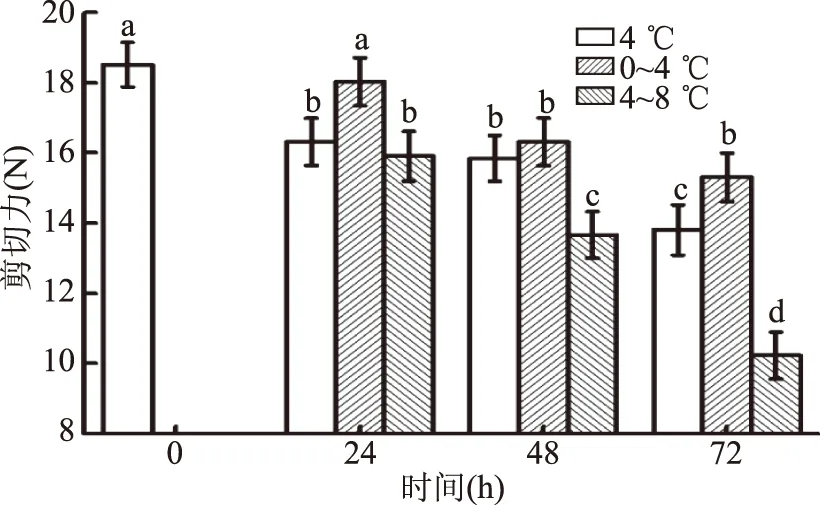
图4 温度波动对冷却猪背最长肌剪切力的影响
2.6 NMR质子弛豫分析
NMR质子自旋-自旋弛豫时间和峰面积比例能够用来反映冷却肉中水分的迁移和分布[27]。表2所示,NMR质子自旋-自旋初始弛豫时间共有3个特征峰:T2b、T21和T22。T2b为结合水,表示肌肉中与蛋白质等大分子结合的水分子和部分肌内脂肪中的水分子,在0~10 ms之间;T21为束缚水,表示肌纤维内和肌纤维间结合较紧密的水分子,20~100 ms之间;T22为自由水,表示冷却肉中能够自由流动的水分,在350~600 ms之间[28]。随着冷藏时间的延长,T2b的起始弛豫时间呈增加的趋势,0和24 h时所用处理组差异不显著(p>0.05),48 h时,4和4~8 ℃处理组显著高于0~4 ℃处理组(p<0.05),4~8 ℃处理组在72 h时最高,表明温度波动对结合水的影响较大,特别是高温波动时。T21和T22的起始弛豫时间随着波动温度的升高而显著增大(p<0.05),束缚水和自由水与肌肉的结合越来越松散[29],水分子移动增强。延长冷藏时间和温度波动能够使水分子与蛋白质等底物的结合越来越松散,加速水分的迁移。
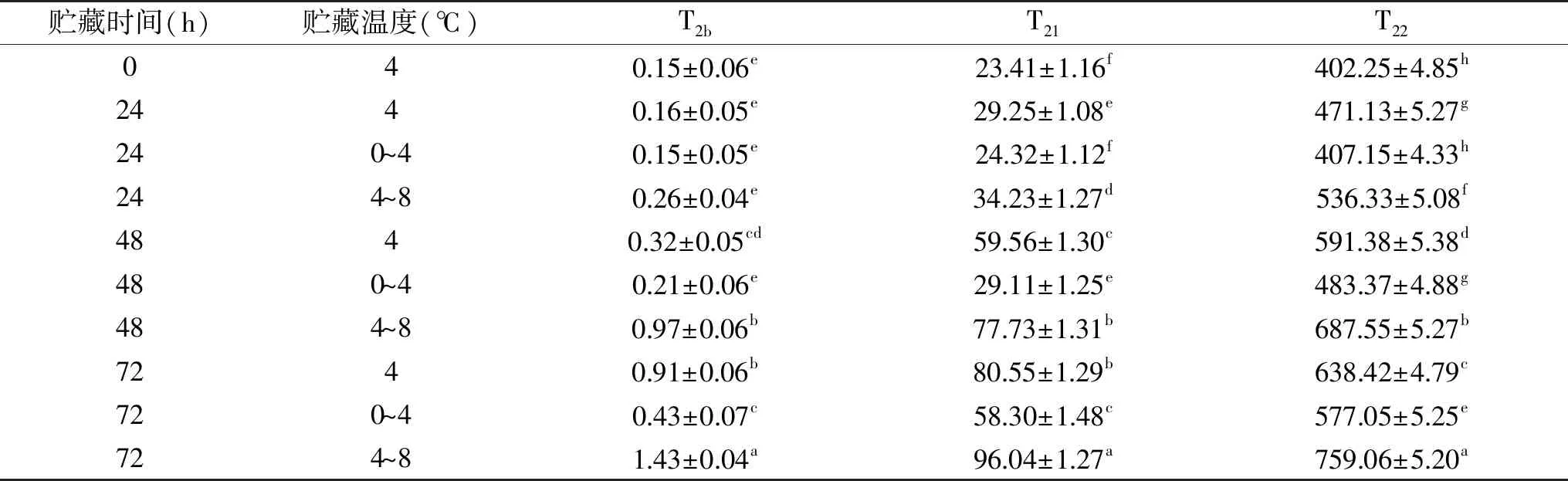
表2 温度波动对猪背最长肌的弛豫时间(ms)的影响
由表3可知,温度波动对猪背最长肌的不同状态水的峰比例影响显著(p<0.05),变化最大的主要是束缚水和自由水。随着冷藏时间的延长,所用处理组的束缚水比例降低,结合水和自由水比例增加,这与冷藏损失结果一致。因为冷藏时间增加,部分肌肉组织和结缔组织被降解,亲水基团暴露,增加了结合水的比例[30-31]。由于肌肉结构被破坏,蛋白质和水分之间的作用力、毛细管力等降低,部分水分被挤出肌肉组织,增加自由水的比例。在相同冷藏时间下,波动温度升高,结合水和自由水比例增加,束缚水比例降低。72 h时,0~4和4 ℃处理组中结合水(P21)和自由水(P22)比例差异不显著,说明低温变温冷藏和恒温冷藏能够降低水分的迁移速度,有利于冷却肉中水分的保持。以上结果表明,随着冷藏时间的增加和波动温度的升高,猪背最长肌的保水性降低。
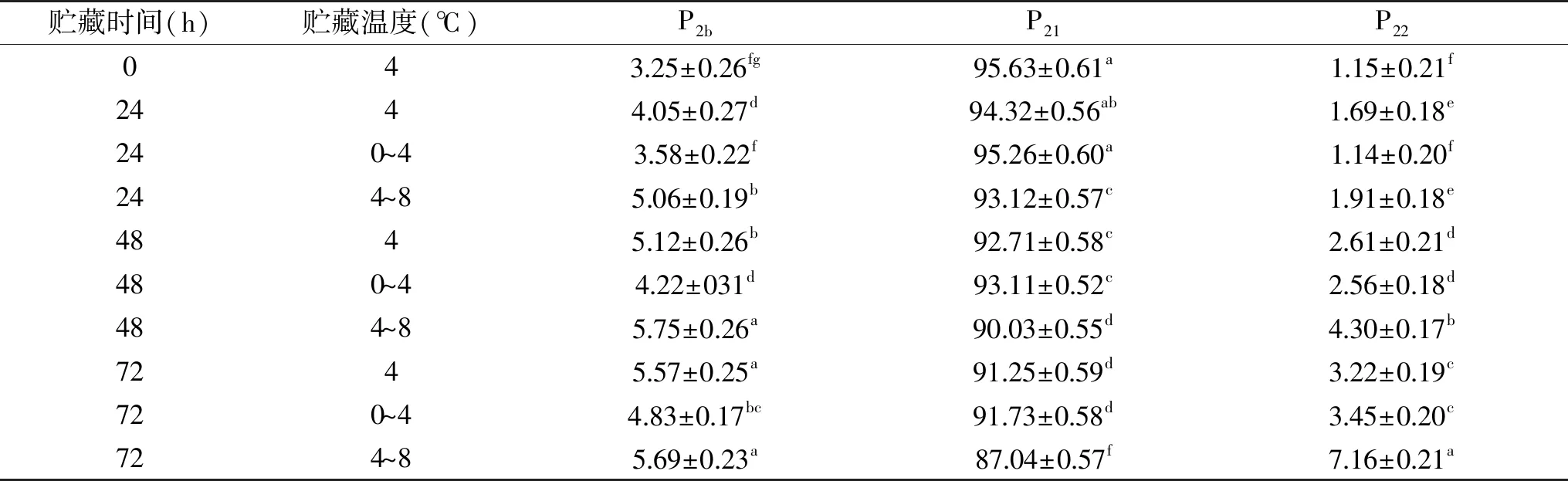
表3 温度波动对猪背最长肌的峰面积比例(%)的影响
3 结论
在4 ℃,0和4 ℃各2 h,4和8 ℃各2 h下冷藏72 h,背最长肌的品质变化显著(p<0.05),冷藏损失率和蒸煮损失率增加(p<0.05),剪切力降低(p<0.05);升高波动温度,pH、b*值、冷藏损失率和蒸煮损失率显著增加(p<0.05),剪切力显著降低(p<0.05),在4和8 ℃各2 h冷藏条件下,72 h时猪背最长肌的冷藏损失率和蒸煮损失率最大,L*值和剪切力最小。随着冷藏时间延长和冷藏温度高温波动,猪背最长肌中水分与蛋白质等底物结合有利于松散,结合水和自由水比例升高,束缚水比例下降。综上所述,冷藏温度波动在4 ℃以下有利于保持猪背最长肌的品质。
