不同减肥方式对雄性肥胖小鼠脂肪组织炎症-芳香化酶轴的影响
2019-08-21马铁赵大林李涛衣雪洁
马铁 赵大林 李涛 衣雪洁
沈阳体育学院(沈阳110102)
对我国22个省市区进行的体质监测发现,超重和肥胖人群比例达到了45.24%,中心性肥胖人数超过一半,并且男性高于女性[1]。男性肥胖后,尤其是中心性肥胖会引起其体内雄激素下降,这可能因为肥胖会抑制经典的下丘脑-垂体-性腺轴(hypothalamic-pituitary-gonadal axis,HPG轴),减少了雄激素的合成。肥胖抑制男(雄)性个体HPG 轴的原因是多方面的,其原因之一是与肥胖男(雄)性体内的高雌激素水平有关[2]。
普遍认为男(雄)性肥胖群体雌激素增加与脂肪组织有关,因为脂肪组织是雌激素主要的合成部位之一,而这种作用在雄性体内表现尤为明显[3]。脂肪组织中含有芳香化酶(Aromatase),是催化雄烯二酮和睾酮转化成雌酮和雌二醇最后一步反应的限速酶,而且芳香化酶表达量与身体脂肪含量成正比,因此男性肥胖后脂肪组织性激素转化作用增强,导致循环系统雄激素下降,雌激素升高,使用芳香化酶抑制剂可以有效降低肥胖男性体内雌激素水平[4-5]。
芳香化酶的增加与肥胖导致的脂肪组织炎症反应有关。研究显示[6],肥胖后脂肪组织促炎因子增多,这些促炎因子会引起脂肪细胞和脂肪前体细胞中编码芳香化酶的CYP19 基因转录加强,进而导致脂肪组织芳香化酶表达增强。有学者提出[7],机体存在“肥胖-炎症-芳香化酶”轴,这一调节轴已在乳腺和内脏的脂肪组织中得到了证实。在这一调节轴中,发挥主要作用的促炎因子包括肿瘤坏死因子α(tumor necrosis factor α,TNFα)和白介素6(interleukin 6,IL6)。TNFα与脂肪前体细胞上的肿瘤坏死因子受体1(tumor necrosis factor receptor 1,TNF-R1)型受体结合后促进芳香化酶表达[8]。IL6与白介素6受体α(interleukin 6 receptor α,IL6-Rα)受体结合后,激活酪氨酸激酶1(janus kinase1,JAK1)-信号传导及转录活化因子3(signal transducer and activator of transcription 3,STAT3)通路,促进脂肪组织芳香化酶的表达,并且其相对独立于TNFα途径[9-10]。
随着男性肥胖者体重的下降,其性激素紊乱得到改善[2],但对于这种变化与脂肪组织“炎症-芳香化酶”轴关系的研究较少。同时研究已经表明,单纯的中等强度有氧运动[11]或降低食物中的脂肪含量[12]都是减肥和降低肥胖所引起炎症反应的有效手段。因此,本研究从“肥胖-炎症-芳香化酶”角度,探讨单纯中等强度有氧运动或单纯降脂饮食干预两种减肥方式对雄性小鼠脂肪组织雄激素芳香化作用的影响,为预防和治疗男性肥胖提供理论与实践依据。
1 材料与方法
1.1 研究对象
3周龄雄性C57BL/6小鼠40只,购自北京维通利华实验动物技术有限公司,许可证号:SCXK(京)2014-0010。按体重随机分为2 组,正常饮食组(N)8 只和高脂饮食组(F)32 只,待小鼠肥胖模型建立后,剔除肥胖未达标8 只,将剩余高脂饮食组小鼠按体重随机分为高脂饮食对照组(FO)、高脂饮食运动组(FE)和高脂饮食转正常饮食(FN),每组8只。
1.2 动物饲养、造模及干预方式
1.2.1 饲养环境、肥胖造模
动物房温度控制在22℃~26℃,湿度在60%~80%。F 组高脂饮食,N 组正常饮食,小鼠自由摄食饮水。每天称量饮食量,每周测量小鼠体重一次。小鼠的正常饲料和高脂饲料[13](脂肪含量21%,其中蔗糖34.1 g/100 g,酪蛋白19.5 g/100 g,纤维素5.0 g/100 g,黄油21.0 g/100 g,矿物质4.9 g/100 g,小麦淀粉15.5 g/100 g)均由沈阳前民饲料厂提供。喂养12 周后,高脂饮食组小鼠体重超过正常饮食组小鼠体重20%,表示雄性小鼠肥胖模型造模成功[14]。
1.2.2 肥胖模型建立后的干预方式
1.2.3.1 8周中等强度有氧运动
肥胖造模成功后,FE组的小鼠继续高脂饲料的喂养,同时施加有氧运动干预。采用8 周跑台运动,跑速、运动时间和运动频率呈递增趋势,3 周后恒定跑速为20 m/min,单次时间60 min,每天运动一次,强度相当于70%~80%VO2max[15-16]。
1.2.3.2 8周饮食干预
肥胖造模成功后,FN 组的小鼠停止高脂饮食,转为正常饲料,但不进行运动,持续8周[16]。
1.2.3.3 8周持续高脂饮食
肥胖造模成功后,FO组单纯高脂饮食,不运动,持续8周。
1.2.4 取材及组织处理
在末次运动后36~40小时,禁食12个小时后取材[17]。称量小鼠体重,腹腔注射10%水和氯醛(350 mg/kg)麻醉。测量小鼠体长(小鼠鼻尖至肛门)后,眼球取血,室温静置凝固后,3500 转/min,离心10 min,取上清。取肾周及附睾周围脂肪、腓肠肌,称重,生理盐水冲洗后,锡纸包裹后投入液氮中,后转入-80℃冰箱保存。
1.3 指标测试
1.3.1 Lee’s指数、体脂百分比和肌肉质量指数的计算
(1)Lee’s 指数计算:Lee’s 指数=体重(g)1/3×102/体长(cm)[18]。
(2)体脂百分比(%)=肾周和附睾周围脂肪重量(g)/体重(g)×100%[19]。
(3)肌肉质量指数(%)=腓肠肌重量(g)/体重(g)×100%[20]。
1.3.2 ELISA 法测试血清、脂肪组织促炎因子(IL6、TNFα),性激素(睾酮、雌二醇)
血清和脂肪组织(脂肪组织放入预冷的PBS中,使用电动匀浆器匀浆,3000转/min,离心20 min,取上清)促炎因子以及性激素测定严格按照ELISA检测试剂盒(R&D Systems)说明书进行。在酶标仪波长450 nm处,读取光密度值,制作标准曲线,计算炎症因子和性激素浓度,每个样本做复孔。脂肪组织中炎症因子和性激素所测结果除以样本总蛋白浓度。
1.3.3 Western blot 测试脂肪组织芳香化酶、TNF受体1(TNF-R1)、IL6受体α(IL6-Rα)蛋白表达
取脂肪组织100 mg,放入1 ml 预冷裂解液中,剪碎,使用电动匀浆器匀浆。4℃,12000 转/min 离心15 min。取上清,使用BCA 法测试脂肪组织中总蛋白含量,沸水煮5 min。调整上样量,每孔上样蛋白20 μg。经SDS 凝胶电泳后(浓缩胶5%,20 min;分离胶10%,60 min),转印(湿转200 mA,60 min)至NC 膜上。5%脱脂奶粉封闭1 h后,孵育一抗(芳香化酶1︰300,TNF-R1 1︰100,IL6-Rα 1︰200,GAPDH 1︰800)4℃过夜。PBST 洗三遍后,孵育二抗(1︰10000)2 h。使用Image Studio荧光成像系统,扫描,并读取灰度值。
1.4 统计方法
结果数据采用均值± 标准差表示,使用SPSS17.0软件系统对数据进行分析,高脂饮食与正常饮食(即FO组与N组对比)比较采用独立样本t检验;不同减肥方式作用效果对比(即FE组、FN与FO组对比)采用单因素方差分析进行统计分析,事后检验采用LSD 方法。P<0.05为差异具有统计学意义。
2 结果
2.1 高脂饮食、有氧运动或饮食干预对雄性小鼠进食量及摄入热量的影响
由表1可知,FO组进食量、摄入热量高于N组(P<0.01)。FE 组进食量和摄入热量低于FO 组(P<0.01)。FN组进食量和摄入热量低于FO组(P<0.01)。

表1 各组小鼠进食量及摄入热量比较
2.2 高脂饮食、有氧运动或饮食干预对雄性小鼠体重和体成分的影响
由表2可知,FO组腓肠肌重量高于N组(P<0.05),FO组体重、腹腔脂肪重量、体脂百分比和Lee’s指数高于N 组(P<0.01)。FE 组体重低于FO 组(P<0.05),FE组腹腔脂肪重量、体脂百分比低于FO组(P<0.01)。FN组Lee’s 指数低于FO 组,肌肉质量指数高于FO 组(P<0.05),FN 组体重、腹腔脂肪重量、体脂百分比低于FO组(P<0.01)。
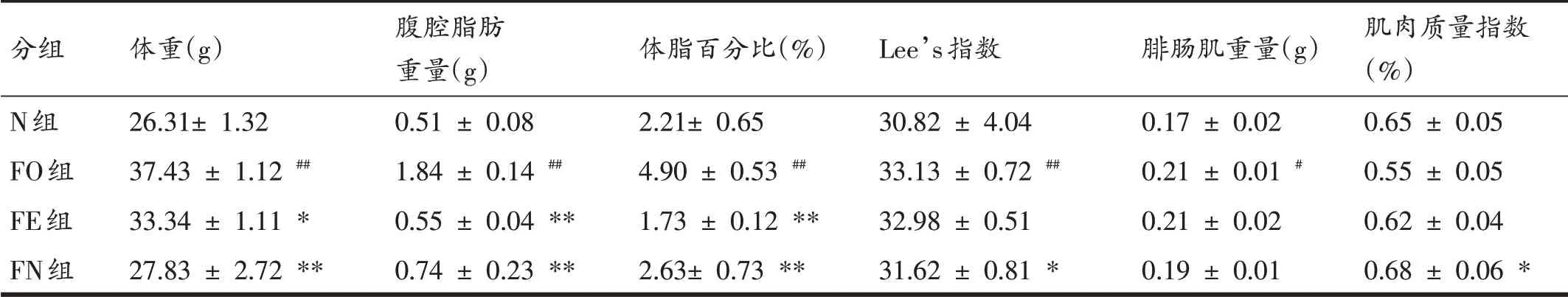
表2 各组小鼠体重和身体成分比较
2.3 高脂饮食、有氧运动或饮食干预对雄性小鼠性激素的影响
由表3可知,FO 组脂肪组织睾酮和雌二醇高于N组(P<0.05),FO组血清睾酮低于N组(P<0.05);血清雌二醇高于N 组(P<0.01)。FE 组脂肪组织睾酮、雌二醇和血清雌二醇低于FO 组(P<0.05)。FN 组血清雌二醇低于FO组(P<0.05);FN组脂肪组织雌二醇和睾酮低于FO组(P<0.01)。

表3 各组小鼠血清和脂肪组织睾酮、雌二醇比较
2.4 高脂饮食、有氧运动或饮食干预对雄性小鼠炎症因子的影响
由表4可知,FO 组血清和脂肪组织IL6 和TNFα高于N 组(P<0.05)。FE 组脂肪组织IL6 和TNFα低于FO组(P<0.05)。FN 组脂肪组织IL6 和TNFα低于FO 组(P<0.01)。FE组和FN组小鼠血清IL6和TNFα低于FO组(P>0.05)。
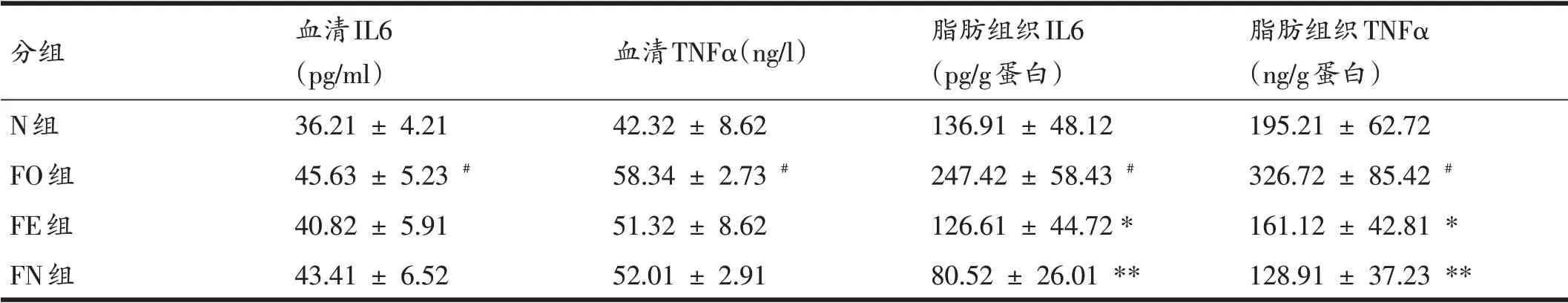
表4 各组小鼠血清和脂肪组织IL6、TNFα比较
2.5 高脂饮食、有氧运动或饮食干预对雄性小鼠脂肪芳香化酶和炎症因子受体的影响
如图1所示,FO组脂肪组织芳香化酶、TNF-R1蛋白表达高于N 组(P<0.05)。8 周有氧运动后,FE 组脂肪组织芳香化酶和TNF-R1 蛋白表达低于FO 组(P<0.05)。FN 组脂肪组织TNF-R1 蛋白表达低于FO 组(P<0.05)。 FE 组和FN 组脂肪组织IL6-Rα受体蛋白表达低于FO组(P>0.05)。

图1 各组脂肪组织芳香化酶、TNF-R1和IL6-Rα蛋白表达量
3 讨论
3.1 高脂饮食对雄性小鼠脂肪组织炎症状态和性激素水平的影响
本研究显示,持续的高脂饮食使肥胖组小鼠体重明显高于正常饮食组,并且内脏脂肪质量、Lee’s 指数和体脂百分比均高于正常饮食组,但肌肉质量指数低于对照组,表明高脂饮食会导致肥胖进行性发展,并且主要是由脂肪含量增加导致的。同时,对比进食量可以看出,持续的高脂饮食使小鼠摄食量高于其他组,进而导致了摄入热量的升高,说明高脂饮食导致肥胖发生的一个重要原因是促进了食欲[19]。此外研究还发现,高脂饮食可以通过改变脂质代谢引发中心性肥胖[21],导致白色脂肪组织细胞肥大、纤维化加强,弹性下降,蛋白质/脂肪比值下降[22]。因此,可以认为高脂饮食通过改变小鼠食欲、脂质代谢和脂肪的结构等多个途径诱发肥胖。
肥胖者机体处于一种“低度慢性炎症状态”[23],表现为循环系统和脂肪组织促炎因子(如TNFα、IL6)增多[17,24],并且脂肪组织可能是炎症反应的早发器官[25]。本研究结果也显示,肥胖组小鼠脂肪组织和血清中TNFα和IL6升高。但对于肥胖导致机体低炎症状态的原因并不完全清楚。有学者认为肥胖发生后,脂肪细胞释放脂肪酸、内毒素增加,激活巨噬细胞转化为促炎症发生的M1型巨噬细胞,进而激活其核因子κB(nuclear factor-kappa B,NF-κB)途径,使得TNFα、IL6、白介素1β(interleukin 1β,IL1β)等促炎因子释放显著增多,抑炎因子减少[19,26]。
脂肪组织炎症反应加强会诱发芳香化酶表达升高,研究发现[27],60%高脂膳食喂养的雄性小鼠8 周后引起脂肪组织炎症反应加剧,芳香化酶表达的升高。本研究也显示,肥胖发生后雄性小鼠脂肪组织芳香化酶与TNF-R1、IL6-Rα表达均升高,提示肥胖引起脂肪组织炎症-芳香化酶轴作用加强。研究还显示,肥胖可以引起局部脂肪组织芳香化酶雌激素转化能力增强[28]。这可能导致了肥胖雄性个体和脂肪组织性激素含量紊乱,如研究发现43名肥胖男性中,80%血清总睾酮低于正常值,28%雌二醇水平达到高雌激素症诊断标准[29]。8 周高脂喂养后,雄性C57bL/6 小鼠血清雄激素显著降低[30]。本研究也显示,肥胖组小鼠血清雄睾酮低于正常饮食组,血清雌二醇高于正常饮食组。
关于肥胖对男(雄)性脂肪组织性激素影响的研究,结果并不一致。有研究发现,肥胖男性体内脂肪组织中雌激素水平与腰围呈正相关,而雄激素含量与腰围呈负相关[31]。但有研究发现,人体脂肪组织含有丰富的类固醇类激素(包括雄激素和雌激素),含量高于循环系统,并推测肥胖者脂肪组织睾酮和雌二醇含量升高[32]。本研究也发现,肥胖组腹腔脂肪组织的睾酮和雌二醇均升高。Gibb等[33]也认为男性血清雌激素增加与芳香化酶催化的底物即雄激素的增加有关。有研究显示[3],脂肪组织芳香化酶过表达后,会引起脂肪组织睾酮和雌二醇均升高,这进一步佐证了高脂膳食诱发肥胖后,雄(男)性脂肪组织中芳香化酶作用增强引发了脂肪组织性激素水平的升高。
3.2 8周饮食干预对雄性肥胖小鼠对脂肪组织炎症状态和性激素水平的影响
饮食干预的减肥方式中,降低食物中的脂肪含量(如采用正常饮食或低脂饮食)是一个可以肯定的减肥手段[34-36]。本研究也显示,高脂饮食转正常饮食后8周,雄性肥胖小鼠体重、腹腔脂肪重量、腹腔脂肪百分比降低。本研究还显示,改变饮食结构后,小鼠进食量和摄入热量均降低。提示,改变高脂的饮食结构可以通过降低小鼠食欲、减少热量摄入发挥减肥作用。与高脂饮食相比,脂肪含量低的饮食还可以改善脂代谢,表现为总胆固醇和低密度脂蛋白的下降[37]。
降低食物中脂肪含量在减肥的同时还可以降低肥胖者的炎症水平,如超重男性经过6周的低脂饮食后,体重降低,同时血清IL6和TNFα降低[38]。C57BL/6雄性小鼠在14周高脂饮食后,进行5周的低脂饮食(脂类占比10%),脂肪组织巨噬细胞浸润减少[39]。10%低脂饮食可以使脂肪组织促炎因子TNFα、单核细胞趋化因子1(monocyte chemoattractant protein-1,MCP-1)降低,并且低脂饮食组的效果要优于高脂运动组[35]。本研究采用8周降脂饮食干预降低了肥胖雄性小鼠脂肪组织促炎因子的水平,并且其效果也要优于有氧运动。但二者差异的原因并不明确,有待于进一步的研究。
有关饮食干预对芳香化酶影响的报道相对较少,而且相关研究以限制热量摄入居多。有研究显示,限制热量摄入(减少30%)的高脂饮食可以有效逆转因前期高脂饮食导致的雌性小鼠乳腺脂肪组织芳香化酶mRNA的增多和活性的加强[40]。限制食物热量摄入(减少40%)可以缓解因年龄增加而导致的雄性大鼠体重增加和睾丸组织芳香化酶降低[41]。而本研究显示,对雄性肥胖小鼠进行饮食干预后脂肪组织芳香化酶蛋白表达出现降低趋势的同时还伴有热量摄入的减少,因此,可以认为减少肥胖个体热量的摄入可以改变其体内芳香化酶表达,但可能存在着器官差异。
但有研究发现[42],高脂饮食转为正常饮食后,在周累计饮食热量无显著下降的情况下,仍可以降低腹腔脂肪蓄积。说明饮食结构改变所导致脂肪含量下降并不单纯由降低热量摄入造成,饮食构成的改变也是一个重要的因素。这提示,改变饮食结构本身也可能会改变肥胖者芳香化酶的表达,但还需要进一步的证实。
本研究显示,降低食物中脂肪含量后雄性肥胖小鼠脂肪组织中促炎因子(TNFα、IL6)及其受体(TNFR1)降低,性激素水平降低,但芳香化酶下降并没有统计学意义。提示降脂饮食干预对脂肪组织性激素的影响并不是主要通过炎症-芳香化酶途径实现的。
本研究显示8 周降脂饮食干预后,肥胖雄性小鼠血清雌二醇水平下降,但血清睾酮并未升高。说明饮食干预对于循环系统睾酮的提高效应较慢。人体研究发现,虽然低脂饮食使肥胖男性的游离睾酮、雄激素结合蛋白升高,但睾酮升高表现出一定的延迟性,其原因并不十分清楚[36]。
3.3 8周有氧运动对雄性肥胖小鼠对脂肪组织炎症状态和性激素水平的影响
无论人体试验[43-44]还是动物试验[45]均表明,不控制饮食的单纯有氧运动可以很好地降低男性(雄性)肥胖者体重、体重指数(body mass index,BMI)、脂肪含量、体脂百分比和血脂等。本研究也显示,8周的中等强度有氧运动,可以有效降低肥胖雄性小鼠的体重、腹腔脂肪含量、体脂百分比和Lee’s 指数,获得了较好减肥效果。
有研究认为单纯的有氧运动有利于肥胖者炎症状态的改善,6周中等强度跑台运动使雄性C57BL/6高脂饮食小鼠血清TNFα、IL6 下降,同时伴有脂肪组织TNFα、IL6 mRNA降低,认为有氧运动可能通过改善脂肪组织缺氧来减少高脂饮食小鼠脂肪组织巨噬细胞数量、抑制巨噬细胞向M1 型转化,进而降低脂肪组织炎症[46]。本研究得出了相似的结果,8周有氧运动可以降低肥胖小鼠脂肪组织和血清的促炎因子水平。
本研究发现,有氧运动可显著降低雄性肥胖小鼠芳香化酶表达。运动干预对肥胖雄性个体脂肪组织芳香化酶影响的研究较少,但有研究发现6 周的自由转轮跑后,肥胖大鼠性腺组织(睾丸)及非性腺组织(肝脏、肾脏)芳香化酶表达显著降低[47]。这从侧面表明,运动减肥后可能对全身各器官的芳香化酶表达有普遍的抑制作用。同时,本研究还发现,运动干预使得脂肪组织中TNF-R1蛋白表达降低,性激素水平降低,提示运动干预可能通过TNFα-TNFR1途径降低了雄性肥胖小鼠脂肪组织芳香化酶的表达,进而抑制了脂肪组织对雄激素的摄取与代谢作用[48]。
从脂肪组织性激素水平来看,运动干预后肥胖小鼠脂肪组织性激素水平均下降明显,而且脂肪组织雌激素/雄激素比值呈现出升高的趋势,并接近正常饮食组。因此可以认为,中等强度有氧运动的减肥方式不仅降低了雄性肥胖小鼠脂肪组织芳香化酶的表达,而且有助于其脂肪组织的雄激素芳香化作用恢复平衡,这会更有利于体内脂肪含量的降低和脂肪细胞代谢的改善。有研究显示[49],降低脂肪组织内雌激素/雄激素比值会加重腹腔脂肪的积累。而睾酮也需要转化为雌激素才能够有效地降低男性体内脂肪含量,这一作用可能与雌激素刺激生长素分泌有关[50],或通过影响局部脂肪组织代谢来实现[51]。但也有研究显示,芳香化酶基因敲除的雄性小鼠,体重升高、脂肪组织增多,伴随着脂代谢紊乱,腹腔脂肪细胞数量和体积的增加[52]。提示保证脂肪组织芳香化酶作用的平衡对于维持男(雄)性体内脂肪含量稳定有着重要作用,但有氧运动促脂肪组织芳香化作用恢复平衡与炎症-芳香化酶轴的作用机制还需要进一步明晰。
本研究结果显示,运动通过炎症-芳香化酶途径有效降低了脂肪组织和循环系统雌二醇水平,但循环系统睾酮并没有发生明显的升高。这可能因为雌激素的减少需要通过削弱其对下丘脑-垂体-性腺轴的抑制作用来发挥促睾酮合成效应[5]。
4 结论
4.1 高脂饮食诱发肥胖会引起雄性小鼠脂肪组织“炎症-芳香化”轴作用增强,进而使机体雌激素水平升高。
4.2 有氧运动可能主要通过TNFα-TNFR1 途径降低雄性肥胖小鼠脂肪组织芳香化酶的表达,并且有助于脂肪组织雄激素芳香化作用恢复平衡。
4.3 降脂饮食干预可能并不是主要通过脂肪组织炎症-芳香化酶途径影响雄性肥胖小鼠的性激素水平。
