吉林珲春保护区哺乳动物种间冷暖季空间共存关系的研究
2019-08-19郎建民潘海峰姜广顺
王 铎 郎建民 潘海峰 姜广顺
(1.国家林业和草原局猫科动物研究中心,东北林业大学野生动物资源学院,哈尔滨,150040;2.吉林珲春东北虎国家级自然保护区管理局,珲春,133300;3.吉林省通榆县林业局,通榆,137200)
生物群落是一个特定地区或生境中具有一定次序的,由多个种群共同组成的集合体[1]。影响群落结构和种群动态的包括非生物和生物因素,非生物因素有温度、光照等,生物因素主要是种间关系:竞争和捕食。同时,通常认为生物之间的相互作用是群落结构和动态的关键因素[2]。局域尺度上的物种共存问题是群落构建和生物多样性维持机制研究的重要组成部分[3],因此,通常以较小尺度上的群落为物种共存的研究对象,侧重于群落内相互作用的物种之间的关系[4]。大多数生态学家认为,捕食作用(predation)、竞争作用(competition)及物理环境是解释群落共存格局所必需的3种因子。季节变化是环境上的综合性影响,季节的交替同时意味着降水量、光照时间和强度等的变化,进而影响初级生产力。这些变化会在食物链中传递,于空间层面上产生种群间的同步效应[5]。吉林珲春东北虎国家级自然保护区位于东北地区,有着明显的冷暖季变化,特别是冬季寒冷且持续时间较长,导致植物生长缓慢,生长周期较长,而动物也会因环境特点有不同的生存策略,空间共存关系随之发生改变。
占域模型(occupancy model)最早由Mackenzie 等[6-7]提出,可根据物种出现数据估算物种空间占有率及动态变化。占域模型可以在动物不是每次都被探测到的情况下,通过重复调查,对目标物种的探测概率进行估算,进而得出特定样点或区域该物种占有的概率[8]。而自动相机技术具有对动物干扰小、能够连续收集数据、资料便于存档检索等优点[8],是一种高效、有用、易于重复、经济高效的野生动物调查和记录技术[9-10],适合用于研究和监测陆地动物,提供了大量的动物监测数据,适合用于占域模型。
1 研究地区概况
吉林珲春东北虎国家级自然保护区(130°14′08″—131°14′44″E,42°24′40″—43°28′00″N)位于吉林省延边朝鲜族自治州珲春市东南部。保护区南北跨越珲春全境,东西呈狭长地带分布,地势北高南低北部全区最高点海拔达973 m,南部最低点海拔5 m,高差大。高山、谷地、河流、湖泊均有发育。分为低中山地貌区、低山丘陵地貌区和河谷及盆地地貌区。气候上属近海中温带季风气候区,气候特点是四季分明,冬冷夏热,雨热同季。年平均气温为5.65℃,极端最高气温为36.3℃,极端最低气温为-32.5℃。河流属图们江水系,河网密布,平均每平方公里河长超过4.3 km,有大小河流52条。多年平均径流深超过400 mm。
2 研究方法
2.1 数据采集
在珲春保护区中进行调查以了解动物活动频繁的区域,以此选定研究区域,划分成3 km×3 km的网格并设置54个相机样点,按照每个网格中2台相机的密度布设相机,每个样点在架设3个月时,更换SD卡和电池,从而完成对研究区域一整年的取样工作。相机架设地点一般选择山脊与水源附近,主要靠近动物活动路径的位置,布设相机的同时记录每个相机位点的GPS信息和周边的环境信息。
2.2 数据整理
更换SD卡时获得数据,进行人工筛选并录制表格。在表格中录入相机编号、相机点坐标、日期时间、动物物种等信息。半小时内拍到的同一物种按照拍到一次计算[11]。统计出监测到的动物物种,根据其出现频次,选出样本数量足够的动物作为研究对象。行(row)名为相机位点编号,每天为1列(column),有监测到动物的位点,在对应的相机位点和时间的单元格内填“1”,没有的用“0”表示[6]。
2.3 数据分析
因珲春地区气候条件及研究需求,将全年相机数据划分为两个时间阶段:将5月到10月划分为暖季,11月到第二年4月划分为冷季。同时相机数据按暖季和冷季分别处理,整理成数据矩阵。
Presence软件是建立占域模型的一个工具,可用于分析并估算占域率和探测率[12]。在Presence软件中对数据矩阵进行拟合,找出使方程收敛,AIC值稳定的最佳拟合天数[13]。占域模型中的物种间单季节模型(two-species single season model)可以计算两个物种同一季节的占域率[14]。通过该模型可得出使用物种互作因子(SIF,species interaction factor),可以反映物种空间共存关系:SIF=1时表明物种间空间分布相互独立;SIF>1时表明物种空间分布上趋于重合;而SIF<1时表明物种空间分布上趋向于分离[14-16]。将检测到的动物物种依据出现频次及不同季节行为差异进行筛选,且依据已有种间关系和食物组成进行组合[17-30]。使用SPSS 22.0对数据进行标准化,将标准化之后的数据矩阵导入Presence软件。
3 研究结果
3.1 自动相机监测结果
研究区域内自动相机监测时间为369 d,共拍摄到哺乳动物15种,隶属4目8科;其中国家Ⅰ级保护动物3种,包括虎(Pantheratigris)、豹(Pantherapardus)、梅花鹿(Cervusnippon);国家Ⅱ级保护动物3种,包括黑熊(Ursusthibetanus)、黄喉貂(Martesflavigula)、马鹿(Cervuselaphus);出现频次共计2 263次,其中冷季867次,暖季1 396次(表1)。
3.2 物种间单季节模型结果
在监测到的15种哺乳动物中依据已知食物组成及种间关系等筛选出12个优势动物放入模型中。其物种间单季节模型结果展示了物种间的空间共存关系(表2)。
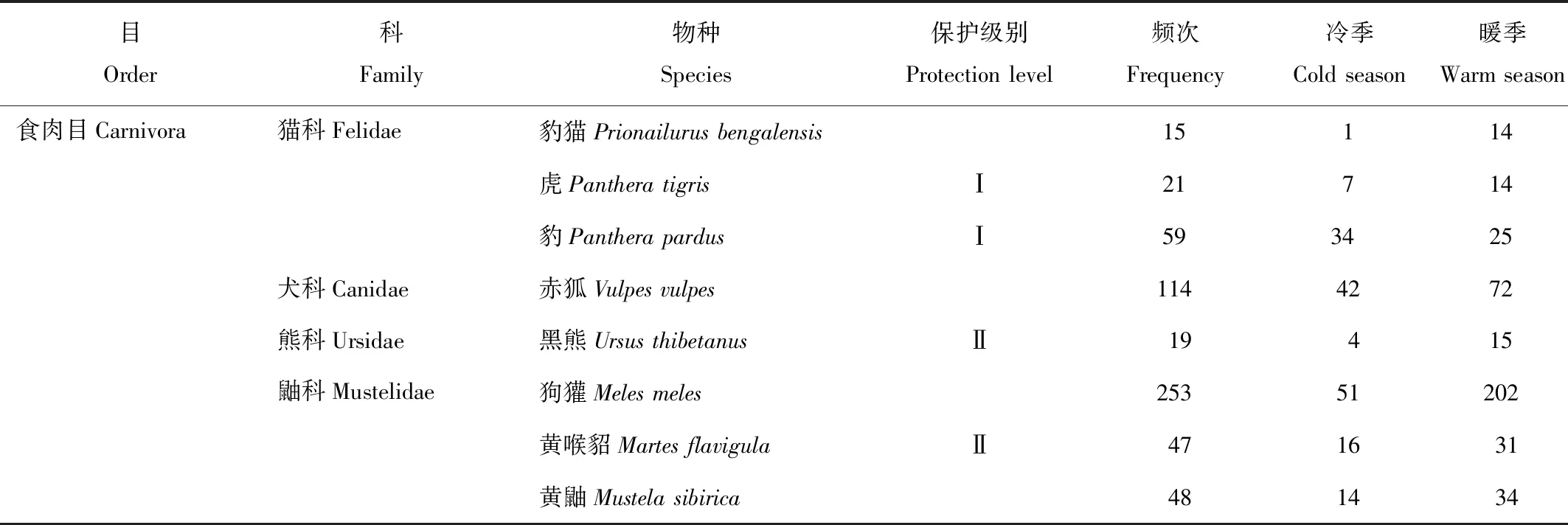
表1 自动相机哺乳类记录Tab.1 Mammals recorded by camera traps
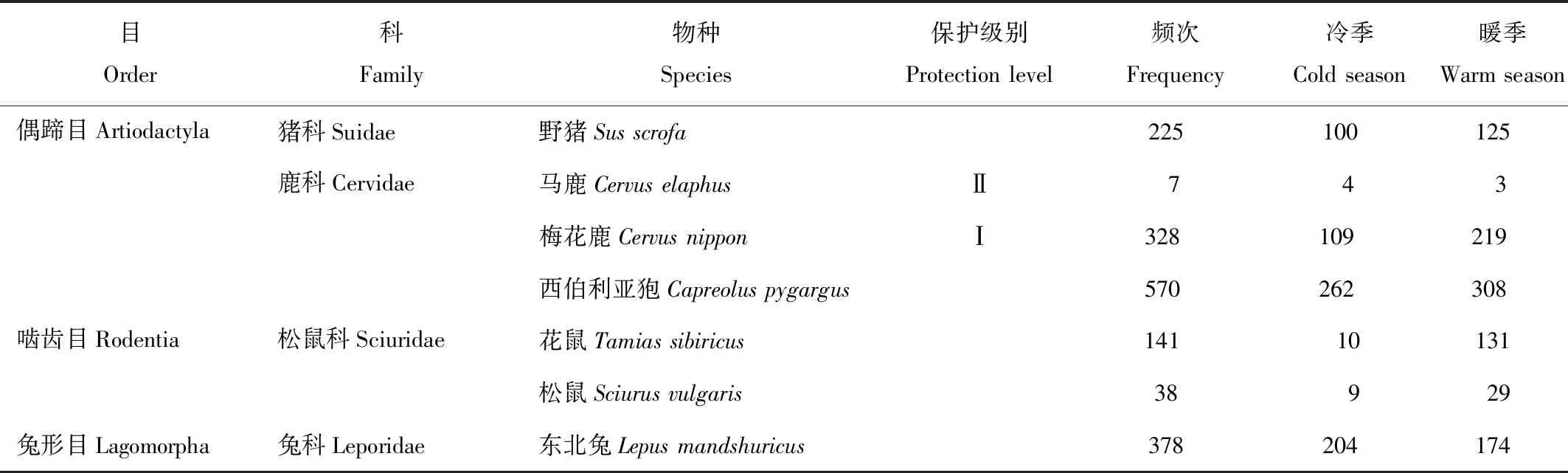
续表1

表2 珲春保护区哺乳动物空间共存关系结果Tab.2 Species interaction factors of mammals in Hunchun Nature Reserve
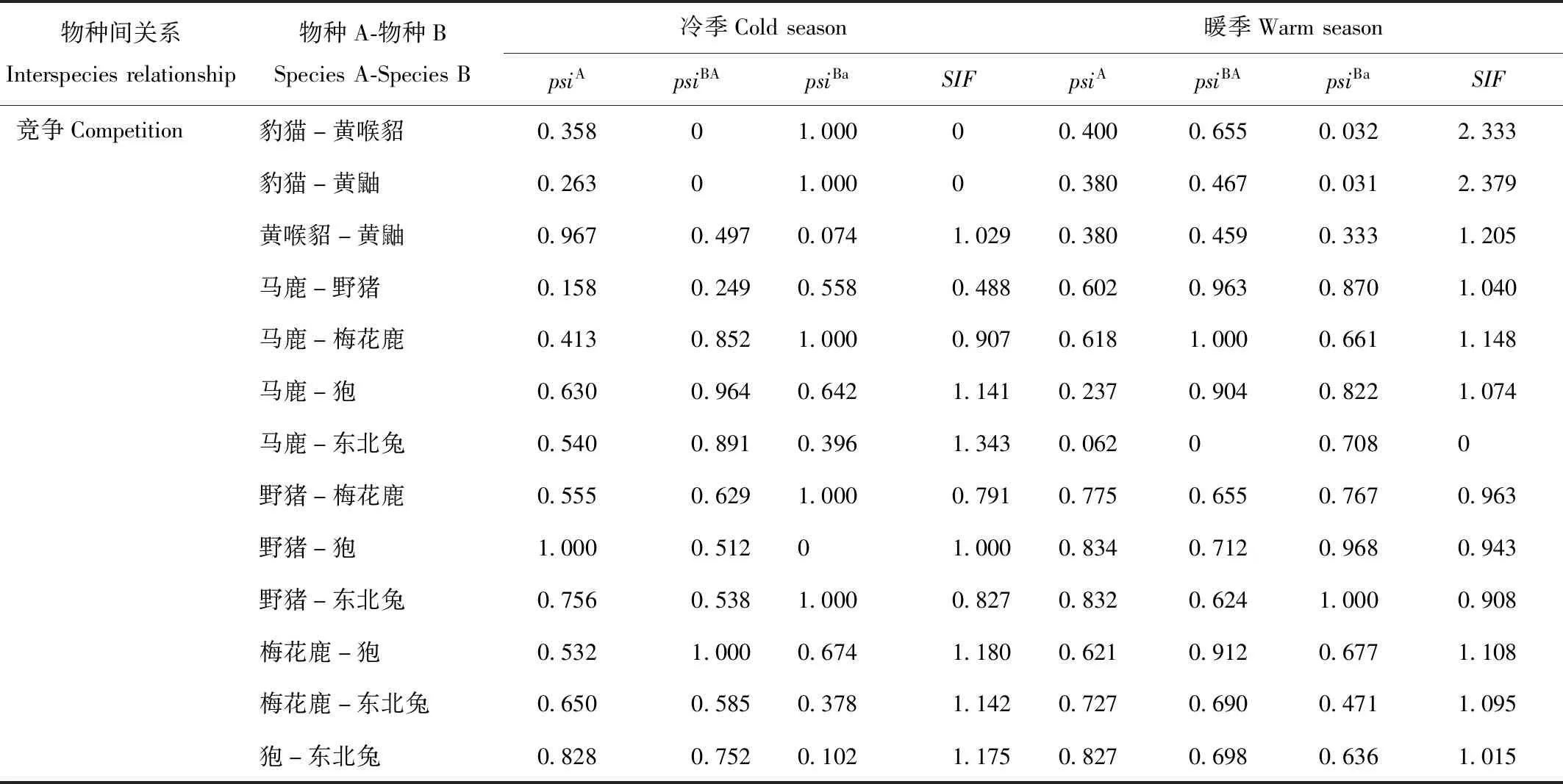
续表2
注:psiA:物种A的占有概率;psiBA:物种A存在时,物种B的占有概率(BA);psiBa:物种A不存在时,物种B的占有概率(Ba);SIF:物种相互作用因子
Note:psiA:Occupancy probability of species A.psiBA:Occupancy probability of species B when species A is present(BA).psiBa:Occupancy probability of species B when species A does not exist(Ba).SIF:Species interaction factor
3.3 捕食关系下的种间空间共存关系
物种间单季节模型结果显示(图1),在捕食关系的物种对中,暖季比冷季的SIF值显著上升的有虎与其猎物马鹿、野猪的物种对,豹与其猎物马鹿、野猪的物种对,豹猫-松鼠,黄喉貂-东北兔,黄鼬-松鼠;暖季比冷季的SIF值明显下降的有虎-狍物种对,豹与其猎物梅花鹿、狍的物种对,赤狐与其猎物东北兔、松鼠的物种对,豹猫-东北兔,黄喉貂-松鼠,黄鼬-东北兔等物种对。
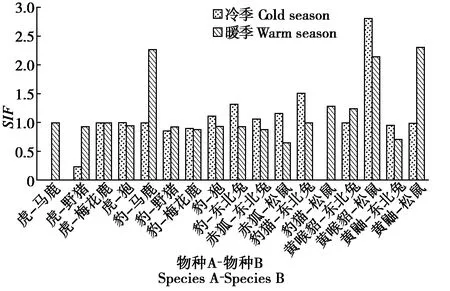
图1 冷暖季捕食者-猎物之间物种相互作用因子变化Fig.1 Changes of species interaction factors between predators and preys in the cold and warm seasons
在冷季,虎与其猎物马鹿、野猪在空间分布上趋于分离,与梅花鹿、狍呈空间分布上互相独立,互作强度均较弱;豹与猎物马鹿空间分布上互相独立,与野猪、梅花鹿在空间分布上趋于分离,互作强度均较弱,而与狍、东北兔在空间分布上趋于重合,互作强度较强;赤狐与东北兔、松鼠均在空间分布上趋于重合,互作强度强;豹猫与东北兔在空间分布上趋于重合,互作强度强,与松鼠在空间分布上趋于分离,互作强度弱;黄喉貂与东北兔空间分布上互相独立,互作强度较弱,与松鼠在空间分布上趋于重合,互作强度较强;黄鼬与东北兔在空间分布上趋于分离,互作强度弱,与松鼠呈空间分布上互相独立,互作强度较弱。在暖季,虎与其猎物马鹿、梅花鹿呈空间分布上互相独立,与野猪、狍在空间分布上趋于分离,互作强度均较弱;豹与马鹿在空间分布上趋于重合,互作强度强,与野猪、狍、东北兔均在空间分布上趋于分离,互作强度弱;赤狐与东北兔、松鼠均呈空间分布上趋于分离,互作强度弱;豹猫与东北兔呈空间分布上互相独立,互作强度较弱,与松鼠在空间分布上趋于重合,互作强度较强;黄喉貂与东北兔、松鼠均在空间分布上趋于重合,互作强度强;黄鼬与东北兔在空间分布上趋于分离,互作强度弱,与松鼠则在空间分布上趋于重合,互作强度强。
3.4 竞争关系下的肉食动物种间空间共存关系
物种间单季节模型结果显示(图2),在竞争关系的肉食动物物种对中,暖季比冷季的SIF值显著下降的有豹-豹猫、豹-黄鼬和赤狐-豹猫;其他物种对暖季的SIF值较冷季都显著上升。
在冷季,虎-豹、豹-豹猫呈空间分布上互相独立,互作强度较弱;豹-黄鼬、赤狐-豹猫、黄喉貂-黄鼬在空间分布上趋于重合,互作强度较强;其他物种对均在空间分布上趋于分离,互作强度弱。在暖季,赤狐-豹猫为空间分布上互相独立,互作强度较弱;豹-豹猫、豹-黄鼬在空间分布上趋于分离,互作强度弱;其他物种对均在空间分布上趋于重合,互作强度强。
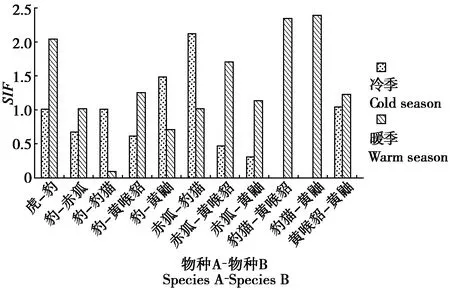
图2 冷暖季肉食动物物种相互作用因子变化Fig.2 Changes of interaction factors of carnivores in the cold and warm seasons
3.5 竞争关系下的草食动物种间空间共存关系
物种间单季节模型结果显示(图3),在竞争关系的草食动物物种对中,暖季比冷季的SIF值显著上升的有马鹿-野猪、马鹿-梅花鹿、野猪-梅花鹿和野猪-东北兔;其他物种对暖季的SIF值较冷季都会下降。
在冷季,野猪-狍为空间分布上互相独立,互作强度较弱;马鹿-野猪、马鹿-梅花鹿、野猪-梅花鹿和野猪-东北兔在空间分布上趋于分离,互作强度弱;其他物种对均在空间分布上趋于重合,互作强度强。在暖季,狍-东北兔为空间分布上互相独立,互作强度较弱;野猪-梅花鹿、野猪-狍和野猪-东北兔均在空间分布上趋于分离,互作强度弱;其他物种对均呈现空间分布上趋于重合,互作强度强。
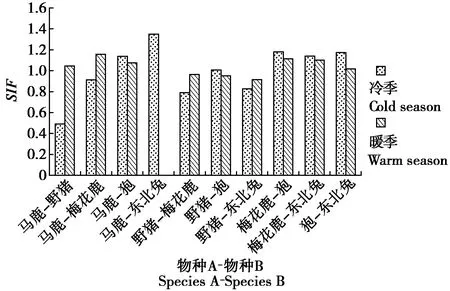
图3 冷暖季草食动物物种相互作用因子变化Fig.3 Changes of interaction factors of herbivores in the cold and warm seasons
4 讨论
依据物种间空间共存模型结果,从物种间的捕食关系来看,捕食者与猎物之间的空间共存关系在冷暖季间有着显著的变化,如豹与其猎物狍、东北兔在冷季中为空间分布上趋于重合,互作强度强,在暖季则与马鹿互作强度较强。可见不同季节下,捕食者对猎物的选择会随环境的变化而发生明显改变。捕食者捕食策略的变化也在冷暖季间竞争关系下的空间共存差异上显现出来,本次研究中的肉食动物物种对不同季节的空间共存关系均存在着明显变化。而草食动物也是如此,证明在冷暖季变化中,草食动物的空间利用及分布也随之产生了显著改变。
空间利用上的重叠同时也意味着动物对同一空间中的资源进行竞争,捕食关系下则为发生捕食行为概率较高,由此可依据空间共存关系对物种间关系进一步理解。具体分析捕食者和猎物的空间共存关系,可知豹在冷季主要追逐狍与东北兔,暖季则追逐马鹿;赤狐在冷季的活动范围与东北兔、松鼠的在空间上趋于重合,暖季则趋于分离,可能是赤狐在暖季中多以红外相机难以拍摄到的啮齿类为食;豹猫冷季多选择东北兔为追逐目标,暖季时为松鼠较多;黄喉貂捕食能力较强,与东北兔、松鼠在全年均有空间分布上趋于重合的趋势;黄鼬与松鼠在暖季呈现强烈的空间分布上分离的趋势,而与东北兔在冷暖季中都是空间分布上分离的趋势,可能是因为东北兔对于黄鼬来说捕食难度较大;虎与猎物的关系均未表现出明显的空间分布重合,这可能是由于虎的数量较少,还需要进一步的研究调查。此外,总体上来看调查的草食动物组合中冷季竞争强度多强于暖季,可能是因为冷季气候寒冷,资源短缺导致食物丰富度下降[31],动物集中于对有限的环境资源进行利用。而肉食动物则是暖季竞争强度多强于冷季,可能是由于东北地区在冷季的恶劣环境条件导致捕食难度增大,使肉食动物改变捕食目标及策略,同时猎物物种冷暖季不同的空间分布模式也会造成肉食动物竞争强度的差异。
物种空间分布是受到多种因素影响的,有来自内部的作用即物种间的相互作用,包括捕食关系、竞争关系等,同时还有外部作用,也就是环境中生境变量的影响[32-33]。季节上的变化同时也意味着环境的改变,物种间空间共存关系在不同季节的显著差异也说明了物种的空间分布不仅受物种间相互作用的影响,同时还有环境的影响。本研究显示了珲春自然保护区内各物种间的空间共存关系在冷暖季变化时基本都存在显著差异,包括捕食关系下的捕食者与猎物之间,竞争关系下肉食动物之间及草食动物之间。期望为在不同季节环境下珲春自然保护区内生境群落的保护与管理提供科学参考。
致谢:感谢研究过程中国家林业和草原局猫科动物研究中心同学们的帮助;感谢野外工作中给予支持和帮助的珲春自然保护区的工作人员。
