卵丘细胞对猪卵母细胞成熟及发育的影响
2019-08-12王庆凯崔茂盛赵子墨付永利唐佩娟李千军
王庆凯,崔茂盛,杨 凌,赵子墨,付永利,张 丹,唐佩娟,李千军*
(1.天津市畜牧兽医研究所,天津300381;2.河北工程大学生命科学与食品工程学院,河北邯郸 056038;3.天津市畜牧总站,天津 100125;4.天津市动物疫病预防控制中心,天津 100125)
获取高质量的体外培养卵母细胞是成功实现动物体外胚胎生产和克隆的关键因素。长期以来,卵母细胞体外培养的研究重点主要集中在提高卵母细胞的成熟效果,在卵丘细胞方面上研究较少。事实上,卵丘细胞对于卵母细胞的成熟起到很大的作用。一些研究证明,与体内成熟的卵母细胞相比,体外成熟的卵母细胞经历体外受精后的着床前胚胎发育能力显著降低[1]。目前认为,体外培养胚胎发育能力降低可能是由于体外培养的卵母细胞胞质成熟不充分造成[2]。此外,一些学者也证明了体外成熟裸卵的发育能力差是由胞质成熟不完善造成[3]。报道称用卵母细胞与卵丘细胞进行共培养可以促进卵母细胞的成熟和后期胚胎发育能力[4],但是目前其作用机制研究不多。因此,研究卵丘细胞在体外培养过程中对卵母细胞成熟效果的影响及作用机制显得十分重要。本实验旨在研究卵丘细胞对卵母细胞体外成熟效果和对孤雌胚胎发育潜能的影响及卵丘细胞对卵母细胞谷胱甘肽(GSH)合成相关基因的影响。
1 材料与方法
1.1 试剂和药品 Medium 199(美国Gibco 公司);微量 RNA 提取试剂盒(德国Qiagen 公司);反转录试剂盒、qRT-PCR 试剂盒(美国GeneCopoeia 公司);在无特殊说明的情况下,细胞培养所需其他试剂均购自Sigma 公司。
1.2 卵巢来源 猪卵巢取自天津市西青区屠宰场,屠宰后将卵巢置于37℃含有双抗的生理盐水中,2~4 h 内运回实验室。
1.3 卵母细胞采集 将带回的卵巢用剪刀去除周边结缔组织,然后用含有双抗的37℃生理盐水清洗3~5 次,然后用10 mL 注射器抽取直径为2~8 mm 的卵泡放入离心管中,待其沉淀后,洗卵,并挑出卵丘在3 层及以上、卵母细胞胞质黑色均一的卵丘-卵母细胞复合体(COCs)用于实验。卵泡分级如图1 所示。
图1 猪卵巢中的卵泡分级
1.4 卵丘细胞的制备 参考黄承俊等[5]的研究,将收集的COCs 放到0.1%透明质酸酶中捶打,脱去卵丘细胞,然后将裸卵移出,剩余液体离心,再用成熟液离心洗涤2 次,加入成熟液使其终密度在1×106~2×106个/mL。
1.5 卵母细胞体外成熟和孤雌激活
1.5.1 COCs 的成熟培养 COCs 采用微滴培养法进行培养,制作100 μL 成熟培养滴,盖上石蜡油,提前5 h放入培养箱平衡,每滴中放20~30 枚卵母细胞。
1.5.2 卵丘细胞与裸卵共培养 取之前制备好的卵丘细胞50 μL,加入到平衡过的50 μL 成熟培养滴中,继续平衡5 min,然后放入裸卵。
1.5.3 裸卵培养 将COCs 放入到透明质酸酶中捶打,待卵丘细胞脱落后,获得裸卵,然后用成熟洗滴洗2次,然后放入到成熟微滴中,将成熟盘放入到39℃、5%CO2、饱和湿度的CO2培养箱中培养。
1.5.4 孤雌激活 体外培养成熟后,将卵母细胞放入透明质酸酶中吹打以至于卵丘细胞脱落,然后将卵母细胞过激活液洗涤3 次,放入融合槽内,1.2 kv/cm、30 μs、一次脉冲。移出后在胚胎发育液(PZM-3)中洗涤3 次,然后转入至胚胎发育盘中,每滴15~20 枚卵母细胞。
1.6 GSH 合成相关基因检测 每组收集100 枚GV 期卵母细胞,利用微量RNA 提取试剂盒提取总RNA,按照反转录试剂盒操作说明获得cDNA,将所得cDNA按照荧光定量PCR 试剂盒说明加入反应体系,反应参数:95℃预变性 3 min,95℃变性15 s;56℃退火30 s;72℃延伸30 s(40 个循环),最后72℃延伸5 min,每个样品每次设3 个复孔。引物序列见表1。

表1 谷胱甘肽合成相关基因序列
1.7 实验设计 实验分COCs 组、卵丘细胞与卵母细胞共培养组、裸卵组。将各组卵母细胞经体外培养42 h后统计各组的存活数及极体数,并检测各组中GSH 代谢相关基因的表达,将各组卵母细胞经电激活后放入PZM-3 中培养,48 h 后检查卵裂率,第5 天检查囊胚率。
1.8 统计分析 采用2-△△Ct法计算目的基因相对表达量,应用SPSS 11.5 统计软件对数据进行单因素ANOVA 分析,使用LSD 法进行多重比较,所有数据以平均值±标准误表示。P<0.05 表示差异显著,P>0.05 表示差异不显著。
2 结果与分析
2.1 卵丘细胞对猪卵母细胞存活率的影响 由表2 可知,COCs 组存活率高于裸卵组(P<0.05),COCs 组和共培养组间差异不显著。
2.2 卵丘细胞对猪卵母细胞成熟率的影响 由表3 可知,COCs 组第一极体率高于共培养组和裸卵组(P<0.05),共培养组第一极体率高于裸卵组(P>0.05)。
2.3 卵丘细胞对猪孤雌激活效果的影响 由表4 可知,COCs 组卵裂率和囊胚率高于共培养组和裸卵组(P<0.05),共培养组又高于裸卵组(P<0.05)。
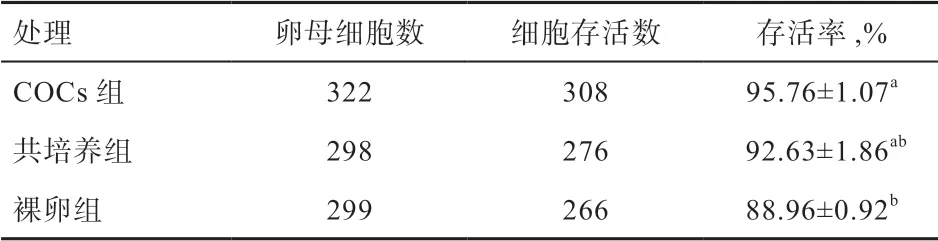
表2 卵丘细胞对猪卵母细胞存活率的影响

表3 卵丘细胞对猪卵母细胞成熟率的影响

图2 极体染色

表4 卵丘细胞对猪孤雌激活效果的影响
2.4 卵丘细胞对卵母细胞GSH 合成相关基因的影响如图3 所示,与COCs 组相比,裸卵组显著下调了GCLM、GCLC 的表达,共培养组中GCLM 的表达与COCs 组相比差异不显著。
3 讨 论
缝隙连接存在于卵丘细胞与卵母细胞之间,其完整与否影响着卵母细胞的成熟[6]。鲁金花[7]研究表明,脱卵丘操作严重损伤了兔卵母细胞,导致卵母细胞不成熟,但脱卵丘操作不影响小鼠卵母细胞。本实验通过透明质酸酶来脱除卵丘细胞获得裸卵,缝隙连接已经受到损坏,卵丘细胞与卵母细胞间的连接机制出现问题[8],通过后期裸卵的发育能力强弱可表明缝隙连接完整的重要性。卵母细胞通过分泌有效的生长因子介导卵丘细胞分化,卵丘细胞则通过缝隙连接传递信号并在卵泡发育的后期阶段支持卵母细胞的生长,促进卵母细胞成熟和受精[9]。卵丘细胞可以提高卵母细胞冻融后的存活率和受精率并支持卵母细胞的体外成熟[10],当两者间的缝隙连接受到破坏时,两者间的协调就会中断,从而影响卵母细胞的整体发育情况[11]。

图3 卵丘细胞对卵母细胞GSH 合成代谢相关基因的影响
本研究结果表明,COCs 组的存活率、第一极体率、成熟率及后期发育中卵裂率、囊胚率均显著高于裸卵组。由此说明,在卵母细胞整个发育阶段中卵丘细胞与卵母细胞的连接机制扮演了重要角色。Tanghe 等[12]研究表明,卵丘细胞在维持卵母细胞减数分裂恢复、促进卵母细胞胞质成熟等方面具有重要作用。郝元斌[13]也证实了去除卵丘细胞后的卵母细胞整个发育指标显著低于未去除卵丘细胞的卵母细胞。本实验中,将卵丘细胞和卵母细胞共培养后,相关成熟指标和发育指标低于COCs组,因为卵丘细胞与卵母细胞共培养系统中,卵丘细胞与卵母细胞间的缝隙连接可能已经受到破坏;但共培养组的极体率和胚胎后期发育指标高于裸卵组。表明在成熟液中添加分散的卵丘细胞时,会提升裸卵的发育潜能,并且旁分泌机制作用在卵母细胞体外培养过程中发挥了重要作用。Hashimoto 等[14]研究也表明,在成熟液中添加卵丘细胞能提高卵母细胞的发育潜能。任文娟等[15]也研究表明,与卵丘颗粒细胞复合体共培养可以提高ICSI 治疗周期中所获未成熟卵的体外成熟率和优质胚胎率。
卵母细胞合成GSH 是其成熟的关键部分,GSH 主要参与抗氧化及抵抗活性氧(ROS),可消除氧化自由基对细胞造成氧化损伤[16]。本实验在成熟液添加了半胱氨酸,半胱氨酸是卵母细胞体外成熟过程中进行GSH 合成所必需的外源底物[17]。Li 等[18]研究发现,在牛体外成熟过程中,卵丘细胞有利于卵母细胞摄取半胱氨酸;且Ozawa 等[19]研究发现,人为去除卵丘细胞可降低胞内GSH 含量,推测卵母细胞外层的卵丘细胞可能通过缝隙链接(GJCs)促进卵母细胞积累GSH。
Takahashi 等[20]研究表明,胚胎能够利用卵丘颗粒细胞代谢胱氨酸后向培养液中释放的物质来进行GSH合成。本实验结果表明,裸卵组中GCLC、GCLM 的表达显著低于COCs 组,并且共培养组中GSH 合成相关基因GCLM 的表达显著高于裸卵组。由此可推断,随着卵丘细胞的减少和缝隙连接的破坏,会造成GSH 合成相关基因的表达降低,从而影响卵母细胞内的GSH合成。
4 结 论
在成熟液中添加分散的卵丘细胞时,会提升裸卵的发育潜能;随着卵丘细胞的减少和缝隙连接的破坏,会造成GSH 合成基因表达降低,从而影响卵母细胞内的GSH 合成。
