非细胞病原体(纳米病原体)及其致病作用
2019-08-12吴克复马小彤郑国光宋玉华
吴克复 马小彤 郑国光 宋玉华
近几个世纪的科技发展表明,不同尺度的时空遵循不同的运动规律,亦即直观世界与宏观、微观世界有不同的运行规则,近半个世纪的研究发现纳米尺度(约0.1~100.0nm)的颗粒有特殊性能,形成了新的研究领域-纳米科学和纳米技术。纳米生物学从纳米尺度研究细胞内部各种细胞器的结构和功能,研究细胞内部、细胞微环境与整个生物体的物质、能量和信息交换。但是,生命科学工作者习惯于直观世界的分析与思维,现有的生命科学理论都是按照直观世界的运行规律建立的,对于近年来科技发展揭示的纳米领域的事物感到困惑,本文从纳米生物学和细胞分子生物学的视角探讨非细胞结构的纳米病原体。
21世纪微生物学研究的突破性进展显示了许多前所未知的微观生物世界。微生物除了细菌、真菌和“经典的病毒-滤过性因子”,还有不能通过除菌滤器的“巨病毒”,它们可以寄生在原虫、细菌和真核生物体内,堪称巨分子生物或纳米生物。微生物间相互作用,形成进化网络[1~3]。地球上微生物的数量和种类惊人之多,是“天文数字”。人体内和体表也有大量的各种微生物,人类细胞与它们共生、互生,共进化博弈构成了微生态系统,堪称“超有机体”[4]。
21世纪组学技术的发展和广泛应用促进了细胞分子生物学的研究,对细胞起源的深层次探索导致众多新课题的提出,细胞生物与纳米生物间以嵌合式进化模式演化,从远古的宏进化到实时机体内外的微进化,无时无刻不在进行之中[5,6]。近年来的研究进展表明非细胞生物和细胞间的基因横向传递(horizontal gene transfer, HGT)在微进化进程中起主要作用,是生物界物质运动的形式之一,也是人类许多疾病发生、发展的基础[7,8]。多种组学技术的同时应用,阐明了许多纳米级的生命现象。
一、病毒是纳米生物与细胞生物的共进化产物
病原生物有细胞生物和非细胞生物两大类,致病性细菌、真菌、寄生虫等细胞致病生物已有专门学科深入研究,病毒是研究最多最深入的非细胞致病生物,属于纳米生物。经过一个世纪的研究形成了病毒学科,医学病毒学是病毒学的核心研究领域,后来提出了朊病毒(prion)等新课题。近10多年来随着研究方法和技术的发展,生命起源、病毒起源等生命科学基础理论课题的研究取得了大量资料,出现了许多新的概念和设想,为非细胞致病生物的研究提供了新的视角和课题。
1.病毒概念的扩展:病毒的细胞内寄生导致多年来认为病毒是细胞的退化产物假设。近年来一些新发现改变了对病毒起源的观念:病毒生态学研究表明病毒是地球上数量和种类最多的生物体;比较生物学研究资料表明病毒基因组的独特性,排除了它源自窃取细胞基因的可能性;感染古菌的病毒的发现提示病毒的起源早于细胞;真核细胞的基因组充斥了源自病毒或相关元件(质粒、转座子、反转录元件等)的基因,提示病毒与其宿主共进化过程中病毒对宿主进化的影响和作用[9]。
病毒是异质性的模糊概念,包括类病毒(感染性核酸)、阮病毒(感染性蛋白)和巨病毒等,也有作者将病毒概念延伸到可移动性元件(转座子),包括基因组的内源性反转录病毒。一般文献所述的病毒是指由核酸和蛋白构成的衣壳编码的生物,有细胞内寄生和细胞外病毒体(virion)两种形式。细胞内寄生形式有两类:virocell,即感染病毒的细胞,细胞不再分裂,但是产生病毒体;另一类称为ribovirocell,也是感染病毒的细胞,但是仍能分裂,也产生病毒体。病毒体是非细胞结构的生物,可以在细胞外和体外环境中生存。病毒在细胞内寄生时有生命的特征(如增殖、代谢等),在细胞外存在时没有生命特征,但保持感染性和结构特征,具有基因组结构。
近年来的研究发现海洋中有大量的微型浮游生物,大小不及白细胞的1‰,它们作为生态群体存在,吸取阳光和化学能及各种必需营养成分,成为食物链的始端,但是未能用培养方法分离研究。近年来用流式细胞仪测定证明它们的存在,初步阐明它们的生长规律,证实属于纳米生物。它们的宏基因组资料表明其代谢的异质性,它们的生命以群体形式构成网络而生存,这类网络由群体和局部的理化环境组成,推测可能与病毒的形成有关,正在研究中[10]。
按照细胞起源的研究假设,所有的细胞生物有一个共同的祖先(last universal common ancestor,LUCA),是核糖体编码的有机体的共同祖先。以LUCA为根画出的进化系统树包含了所有现在的细胞生物。病毒的进化画不出进化系统树,因为它们是多源的,没有共同的祖先,不同类别的病毒有不同的起源[11]。病毒与细胞生物共进化,呈嵌合式进化,即病毒在机体内、群体内微进化过程中有基因的横向传播和纵向传递,在病毒获得机体细胞基因过程中,易于基因突变成为致病基因[6]。
2.基因横向播散的医学意义:基因的纵向传递是种族繁衍的基础,基因的横向播散提供了变异-适应环境变化的基础,二者都是生物进化的基础,异常变化则可以导致疾病的发生、发展。遗传性疾病是异常基因纵向传递的结果,异常基因的横向播散导致各种传染病和一些少见病的发生。病毒是研究最多的引起基因横向传递的生物因子,质粒、转座子等作为生物工程的工具已应用多年,但是在疾病发生、发展中的作用和意义的研究尚在起始阶段。
可移动元件(transposable element, TE)是指有从基因组的一个位点移位到另一个位点能力的核酸片断,又称转座子或“跳跃基因”,在自然界广泛分布,真核细胞基因组序列中都有转座子,约占人类基因组的45%。按它们转座的介质是DNA或RNA分为两类:①DNA转座子,其DNA序列通过由该元件编码的转座酶作用下的“切割与粘贴”机制移位,大多数处于静止状态;②逆转座子的DNA序列通过“复制和粘贴”机制转座。逆转座子可再分为含长末端序列(long terminal repeat,LTR)和不含长末端序列(non-LTR)的两亚类,LTR逆转座子即内源性反转录病毒,构成约8%的人类基因。non-LTR逆转座子在人类基因组中由于点突变、重排或缺失,仅少数处于活跃状态,已报道有124个能引起疾患,这类自治性的长分散元件(long interspersed element-1, LINE-1或L1)是现代人类活跃的反转座子,通过它编码的反转录酶不仅能复制自身的RNA,也能复制其它的RNA(如Alu,SVA),还能转座细胞的mRNA,结果形成伪基因[12]。逆转座子含有若干DNA调控元件:转录增强结构域,剪接信号、转录因子结合点,抑制信号等。可以在基因组内跳跃,可以引起基因组不稳定或重排,或插入性变异,剪接异构,3′或5′转导,早熟和多聚腺苷化,产生伪基因,延长无效转录,甲基化等。逆转座子的直接效应通常对细胞是有害的,甚至可以引起人类疾病,如肿瘤等,也可以导致基因组进化[13,14]。
TE在维持基因组整合性,胚胎发育和适应环境微进化中起重要作用,TE自调节和机体调节机制协同调节TE活性,失调时可以导致疾病的发生。DNA甲基化和组蛋白修饰是TE的调节机制,已有的研究表明这些表观遗传学机制可以通过激活TE起致癌作用。甲基化的完全丧失可以导致TE激活,基因组不稳定,染色体断裂,进而启动致癌机制。如直肠结肠癌、肝细胞癌和乳腺癌中就有DNA去甲基化引起的逆转座子LINE1的激活。LINE1、ALU、SVA是最常见的插入性致变异的TE。致癌基因驾驭者的活化也由肿瘤抑制基因的异常插入和调节序列的混乱介导。基因表达的失调、剪切引起的蛋白片断和基因组的不稳定性都引发致癌机制。基因组不稳定包括染色质破裂和重组都能引起插入变异[15]。在人类进化过程中插入了上万个可移动元件,过去认为转座子插入是无害的,近年来的研究资料表明插入引起的基因组改变可引起转录改变,引起有害的效应,尤其是致癌效应,如白血病、乳腺癌、卵巢癌、结肠癌等。虽然相关研究尚在起始阶段,展示了新的诊疗方向[16]。
近10多年来的研究表明,miRNA参与基因表达调节,在发育、细胞代谢、凋亡和疾病的发生、发展中起重要作用。深入研究发现多数miRNA的形成源自转座子,人类基因组近半数基因由转座子驾驭,转座子可以通过miRNA进入机体的调控网络,影响功能和微进化进程,其影响程度可以小到忽略不计,也能大到降低基因组稳定性导致疾病的发生、发展[17,18]。
二、朊病毒和蛋白折叠错误病(protein misfolding disorders,PMDs)
从1937年发现羊的搔痒症,到完全重组的朊蛋白在体外复制,近80年的研究充斥了非传统观念的提出和发展(表1)。提示这个领域与我们熟悉的以细胞为基础的医学生物学范畴有不同的基础,可能是非细胞的纳米病原体,遵循人们还不熟悉的运行规律不断地演变[19]。

表1 prion假设的研究进展[19]
朊病毒(prion)是有传染性的以自身为模版的蛋白聚集物。最早是在人和动物的海绵状脑病发现的,是一类仅有蛋白成分的感染因子,通常以可溶性形式存在,但是可以折叠组装成能自复制的prion多聚体。通常以散发性或家族遗传性形式出现。但是,也能以获得性甚至跨种属传播(如疯牛病),造成恐慌和畜牧业经济损失,引起了广泛关注。后继研究在低等的真核细胞也发现prion,对于宿主可以有害,也可以有利,在真菌可以通过细胞分裂纵向传递,也可以通过细胞接合横向传播。近期研究发现哺乳类动物细胞可以有酵母prion结构域的同系物NM产物,在培养体系中有从可溶性到聚集态到持续自复制的生活史,成为研究prion的模型。酵母朊蛋白的发现和证实提供了不通过核酸遗传信息改变,从蛋白折叠改变传递功能改变的新途径。prion假设的实验研究已经充分证明朊病毒是细胞蜕变的产物,在体内环境中可能有诸多因素导致PrP基因编码蛋白的折叠错误,PrP基因编码蛋白在折叠错误后可以有感染性,有待深入的体内研究[20]。
人和动物海绵状脑病的共同特征是在脑组织中堆积了大量折叠错误的蛋白导致神经原死亡和淀粉样变。克雅症(Cresutzfeldt-Jakob病,CJD)是人类的海绵状脑病,其发生率约为(1~2)/1000000。临床表现为快速进行性痴呆,共济失调,肌阵挛和行为改变。85%的病例是散发性的,10%~15%是家族(遗传)性的,不到1%的病例是传染的(表2)。
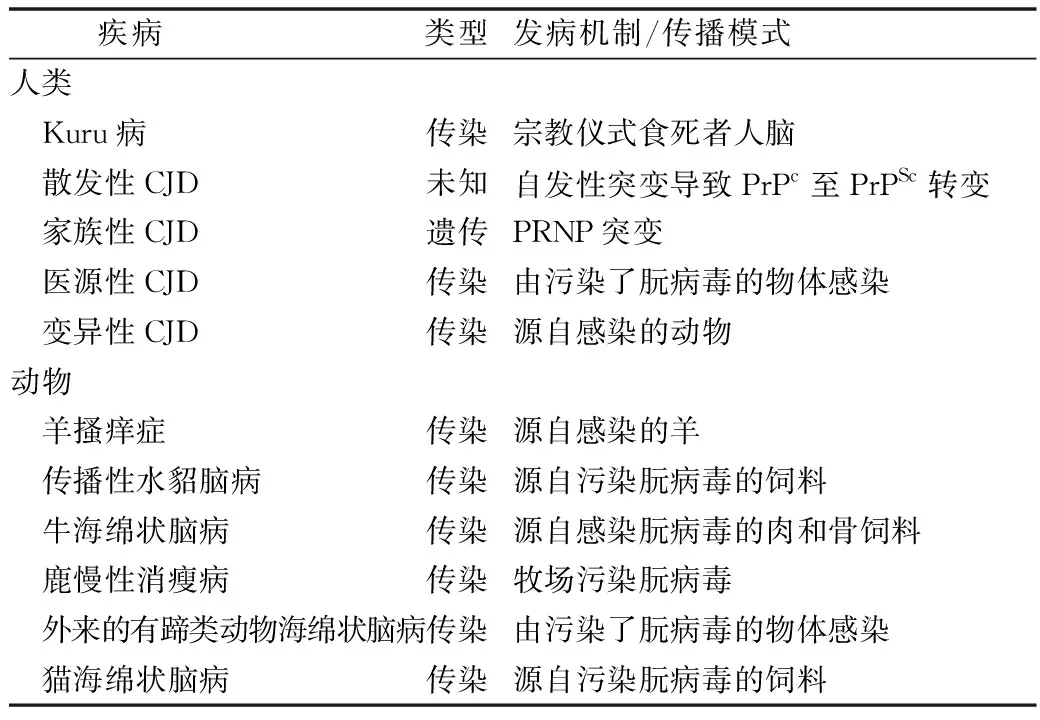
表2 人和动物的朊病毒病[21]
朊病毒蛋白(PrP)由染色体基因PRNP编码,哺乳类动物有保守的正常细胞朊病毒蛋白PrPc,主要由α螺旋构成;致病的朊病毒蛋白PrPSc由β板层构成,PrPc转变成PrPSc是致病的关键分子机制。从动物的朊病毒病观察到PrPSc的复制、堆积并耐受蛋白酶解。由于朊病毒的跨种属传播性,动物群体的prion病传播增加了人类罹患prion病的风险。PrPSc的传播通过消化道途径,然后经过淋巴组织再进入脑组织[21]。
近年来的研究结果表明:蛋白折叠错误相关疾病与朊病毒疾病有共同的分子机制,包括以自身为模版的复制和最终能播散。还有共同特征如病原体株的可变性。至少有一种蛋白或多肽折叠错误、聚集堆积在组织中,导致细胞损伤和器官功能失调,导致蛋白折叠错误病。这组疾病至少有25种病,包括阿尔茨海默病、帕金森病、亨廷顿病和播散性海绵状脑病(朊病毒病,prion病),其中prion病研究得最多。近年来的研究表明,2型糖尿病(T2DM)的胰岛细胞中有蛋白折叠错误形成淀粉样多肽的堆积,可能在胰腺损伤中起重要作用[22]。近年来的研究发现动脉粥样硬化斑块中也含有淀粉样变的蛋白纤维,与阿扑脂蛋白相关[23,24]。形成淀粉样蛋白沉淀是许多疾病的共同特征,其作用和基础正在研究中。值得注意的是淀粉样变蛋白的致病作用多数与脂质稳态相关,与脂质传递、结合至膜中或脂质代谢途径相关。亦即淀粉样变蛋白往往结合脂质,脂质通常参与这些蛋白的正常折叠。所以脂质可能在淀粉样变相关疾患的发生、发展中起重要作用[25]。
朊病毒病和阿尔茨海默病(AD)不仅有临床相似性,还都有蛋白折叠错误和沉积的病变机制。研究表明朊病毒可以通过外泌体(exosome)和细胞纳米管道传播,外泌体参与神经退行性疾患的发展,细胞朊蛋白(PrPc)在外泌体高表达。在神经退行性变疾病PrPc至少起两种作用,在朊病毒病中PrPc是致病蛋白PrPSc增殖的底物;在AD中PrPc结合有神经毒性的淀粉样β寡聚体(Aβ),后者启动和促进AD的发展,详见表3[26,27]。
三、外泌体的病原—发病学作用
从原核细胞到真核细胞的进化过程中细胞形成了多种细胞间通讯机制,包括细胞接触,分泌因子和细胞外囊泡(extracellular vesicle, EV)。EV是异质性的囊泡,按大小分为3类,即外泌体(exosome,30~150nm)、微泡(microvesicle, 100~1500nm)和凋亡体(apoptotic body,1000~5000nm)。外泌体与大囊泡的区别不仅体积小,其形成机制和内含物也不同,外泌体由质膜形成或由多囊泡体形成,能在释放细胞和接受细胞间交换所含的蛋白、核酸、脂质等细胞内容物。不同细胞可有配体-受体、融合、吞噬、内吞等多种接受外泌体的机制(表4)。近年来外泌体的病理作用受到研究者的关注,成为研究疾病发生、发展机制的新热点。
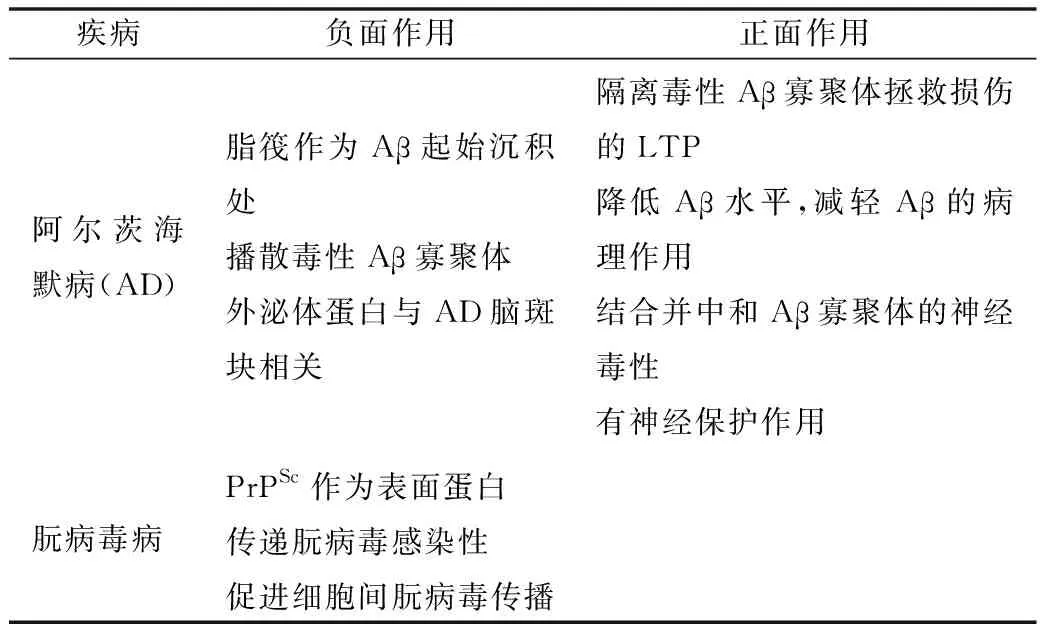
表3 外泌体PrP在不同疾病中的功能比较[26]

表4 接收外泌体信号的细胞机制
*蛋白/基因转移可引起细胞反应或转移到溶酶体降解
外泌体参与机体的正常生理机制调控,如组织分化和修复、造血干细胞增殖分化、凝血、妊娠、免疫监测等。外泌体也参与多种病理过程,已经在多种人类的疾病进行研究,如HIV-1感染、自身免疫病、帕金森病、阿尔茨海默病和多种炎症状态。许多病毒利用外泌体的性质和功能逃避机体的免疫机制,有些病毒能劫持细胞外囊泡而播散。外泌体的摄取不依赖病毒受体,也不需外膜,可能是病毒持续性感染的机制之一[28]。
近年来的研究表明,肿瘤细胞衍生的外泌体(tumor-derived exosomes,TEX)在肿瘤的发生、发展中起重要作用。TEX的内容物包括蛋白、脂质、mRNA、miRNA、DNA等,参与对病原体和肿瘤的免疫反应,以及自身免疫调节。TEX可以很快被邻近细胞内化,也可以被其他组织细胞内化或进入体液循环[29,30]。如急性白血病细胞表达的抗凋亡蛋白Bcl-2、Mcl-1、Bcl-X、Bax等通过血浆外泌体影响病程,可作为诊断指标。研究表明癌变过程中囊泡的产率增加,其内容物反映该细胞的内容,检测体液中的肿瘤外泌体可作为“无创伤活检”[31,32]。
TEX的功能可归纳为:(1)肿瘤细胞的自分泌效应,增强肿瘤的生存、增殖、转移能力;从胞质中去除药物,改变药物代谢,增强肿瘤的耐药性。(2)对肿瘤微环境的影响:促进血管新生,使肿瘤微环境中的免疫细胞向亲肿瘤极化,细胞外基质重建,骨髓重编程使之适于肿瘤转移。(3)对免疫机制的影响—抑制抗肿瘤免疫反应,使活化的抗肿瘤效应细胞凋亡;扩增和上调有免疫抑制活性的Tregs、MDSCs和Bregs细胞,干扰髓系、淋巴系和树突细胞分化;刺激肿瘤特异性免疫反应;从肿瘤细胞向正常细胞传递蛋白、脂质、核酸、突变基因,改变其表型和功能,从而干扰免疫治疗[33~36]。
研究表明慢性淋巴细胞白血病(chronic lymphocytic leukemia, CLL)藉助CLL细胞产生的TEX维持与外周微环境持续的“对话”,保持适宜于CLL细胞增殖和生存的状态,CLL的病情与TEX的分泌、组成及功能状态相关,是CLL难以治愈的原因之一[37]。
病毒颗粒与外泌体有类似的性质和功能,反转录病毒样颗粒是内源性反转录病毒的产物,在黑色素瘤、乳腺癌、浆细胞瘤和胶质母细胞瘤细胞系和牛皮癣、乳腺癌患者血清中测出,其病原学意义有待深入研究阐明。
肿瘤病毒和慢性感染的病毒对细胞外囊泡的产生和成分有影响,可能含有病毒的miRNA、蛋白甚至整个病毒颗粒,影响邻近细胞和机体对病毒的识别。HIV-1病毒与外泌体的关系已经深入研究,反转录病毒劫持外泌体用特洛伊木马策略进入CD4+T细胞,在体内繁殖播散。
综上所述,纳米生物学研究方兴未艾,有待于建立理论体系。量子力学是阐述微观世界运行规律的物理科学,量子生物学(quantum biology)运用量子力学的理论、概念和方法研究生命过程和分子动态结构,已经阐释了光合作用、磁场导航和视觉机制,在酶的作用机制研究中展现出独特的作用,随着细胞分子生物学研究的深入和发展,期望量子生物学能在纳米生物学机制的研究中开辟新的途径[38]。
