基于下一代测序技术分析巴氏杀菌乳中残留细菌在贮藏期间的动态变化
2019-07-26丁瑞雪耿丽娟张铁华孙雪姣岳喜庆武俊瑞
丁瑞雪,耿丽娟,张铁华,孙雪姣,岳喜庆,武俊瑞,*
(1.沈阳农业大学食品学院,辽宁 沈阳 110866;2.沈阳市食品检验所,辽宁 沈阳 110136;3.吉林大学食品科学与工程学院,吉林 长春 130062)
巴氏杀菌乳是通过低温杀菌的一种乳品,具有新鲜、营养、原味、安全、环保等优势,在奶业发达国家消费中占据主导地位,如加拿大、美国、澳大利亚、日本等国巴氏杀菌乳所占比例均达90%以上[1-3]。由于巴氏牛乳的处理方法比较温和,较好地保存了牛乳的营养与天然风味,因此在国内外迅速发展,得到了消费者的认可[4-5]。但由于杀菌温度较低,冷链温度并不能总保持在推荐范围内,特别是在家用冰箱中,这增加了食品在贮存过程中发生变质和出现潜在致病性微生物生长的风险[6-7]。因而不能杀死鲜乳中全部的微生物和酶,仍有部分孢子形成菌和耐热细菌可能在这种环境下存活[8-9]。已有研究表明,巴氏杀菌乳货架期都是单纯依靠纯培养技术判断,此种方法仅能获得实际检测样本中不到1%的微生物[10],而绝大多数微生物过去都由于技术原因被忽略,由此可见当前普遍采用的依靠纯培养技术获得的乳制品贮藏条件和货架期等参数可能不准确。同时,由于这些方法耗时且劳动密集,可能无法提供微生物群落多样性的真实图景。
近年来,随着高通量测序技术的迅速发展,高通量测序技术早以应用到各个领域,包括土壤[11]、海洋[12]和肠道环境[13-14]。通过使用高通量测序技术,之前与牛奶没有关联的细菌属,如拟杆菌、粪杆菌,现已经能够在牛奶中被检测到[15]。张哲等[16]应用Pacbio SMRT测序技术分析传统发酵驼乳中细菌和真菌的多样性,发现有子囊菌门、硬壁菌门的出现。此外,Watanabe等[17]的研究结果也有类似的结果。其他研究[18]也利用高通量测序技术对当地的两家奶牛场牛奶样品的微生物组成进行了表征,通过对季节性的研究,发现冬季有假单胞菌属、链球菌属和不动杆菌属的产生。由此可见,高通量测序技术的应用对乳制品中的微生物多样性提供一个更加全面的分析[19-20],使乳制品微生物方面的研究实现了一次历史性的跨越。
因此,本研究对不同贮藏条件下巴氏杀菌乳中的微生物组成进行表征,利用宏基因组学技术方法对不同贮藏条件的巴氏杀菌乳样品中残留的耐热微生物种类和动态变化规律进行探究。
1 材料与方法
1.1 材料与试剂
采集国内大型3 家乳品企业(GZ、CG、HS)正常工艺当天生产的巴氏杀菌乳,各家66 份,共198 份成品,分别置于0、4、10 ℃的恒温箱中保存,运回实验室进行实验。
DNA提取试剂盒、引物、配制聚合酶链式反应(polymerase chain reaction,PCR)体系中的脱氧核糖核苷三磷酸、10×PCR buffer、r-Taq酶、Loading buffer、Marker DM2000、Gel-red核酸染料 上海美吉生物有限公司。
1.2 仪器与设备
Axy Prep DNA凝胶回收试剂盒 美国Axygen公司;DYY-6C电泳仪 北京六一仪器厂;凝胶成像系统美国伯乐公司;ZHJH-C1112C无菌操作台 北京瑞尔欣德科技有限公司;YCD-EL259医用冷藏冷冻箱中科美菱低温科技股份有限公司;HN-50S电热恒温培养箱 邦西仪器科技(上海)有限公司;MIR-254-PC低温恒温培养箱 日本松下健康医疗器械公司;GeneAmp®9700型PCR仪 美国ABI公司;TGL-168高速台式离心机、超低温冰箱、微量紫外-分光光度计德国Eppendorf公司。
1.3 方法
1.3.1 样品采集及前处理
GZ乳业的工艺条件:原奶验收→净乳→预巴杀→冷藏→定容检验→预热均质→巴氏杀菌→冷藏罐装→成品。
CG乳业的工艺条件:原奶验收→净乳→均质→巴氏杀菌→降温罐装→成品。
HS乳业的工艺条件:原料乳的验收→过滤与净化→标准化→均质→巴氏杀菌→冷却灌装→成品。
将采集的3 家巴氏杀菌乳混合均匀,分别置于0、4、10 ℃三种不同温度的恒温箱中进行贮藏,并分别于0、3、6、9、12、15 d进行取样,备用。
1.3.2 总DNA的提取与检测
取不同贮藏条件下的巴氏杀菌乳样品10 mL放入离心管中,离心后去除乳脂层,再次离心后收集沉淀用pH 7.4磷酸盐缓冲液(phosphate buffer saline,PBS)充分洗涤,并移入干净的离心管中,按照Soil Pure超纯土壤基因组DNA试剂盒要求进行操作,完成巴氏杀菌乳样品中细菌DNA的提取,利用1%琼脂糖凝胶电泳检测提取的基因组DNA,最后将DNA溶液于-20 ℃条件下保存,以备后续操作。
1.3.3 16S rRNA PCR扩增及荧光定量电泳检测
本研究选择的扩增区域是细菌16S rRNA的V3-V4区,利用已提取的DNA作为模板,选择特异性引物338F(5’-ACTCCTACGGGAGGCAGCAG-3’)和806R(5’-GGACTACHVGGGTWTCTAAT-3’)。PCR扩增程序为:95 ℃预变性3 min,27 个循环(95 ℃变性30 s,55 ℃退火30 s,72 ℃延伸30 s),最后72 ℃延伸10 min。扩增体系为20 μL,4 μL 5×FastPfu缓冲液,2 μL 2.5mmol/L dNTPs,0.8 μL引物(5 μmol/L),0.4 μL 5×FastPfu聚合酶;10 ng DNA模板。将每个样本扩增3 份,同一样本的PCR扩增产物通过琼脂糖凝胶电泳检测,检验其PCR扩增是否成功。再参照电泳初步定量结果,将PCR产物用Q uanti FluorTM-ST蓝色荧光定量系统进行检测定量,之后按照每个样本的测序量要求,进行相应比例的混合,将PCR产物送于上海美吉生物科技有限公司进行测序。
1.3.4 MiSeq文库的构建及测序
首先通过PCR将Illumin官方接头序列添加到目标区域的外端,按物质的量比例混合后在MiSeq测序平台进行测序。使用QIIME(quantitative insights into microbial ecology)软件去除低质量序列,再进行生物信息学分析。在97%的相似性水平上归类和确定可操作分类单元(operational taxonomic unit,OTU)数量,同时通过Shannon指数曲线评估每个样品的微生物多样性和测序结果的多样性。
2 结果与分析
2.1 不同条件巴氏杀菌乳中细菌PCR扩增结果

图1 不同条件巴氏杀菌乳中细菌16S rRNA基因PCR扩增产物琼脂糖凝胶电泳图Fig. 1 PCR amplification results of bacterial 16S rRNA genes from pasteurized milk samples at different storage temperatures for different times
将样品DNA提取后,采用细菌338F-806R通用引物对浓度、吸光度均适宜的DNA样品进行PCR扩增。扩增结果经2%的琼脂糖凝胶电泳进行检测,由图1可知,16 份样品细菌在750 bp处可见扩增条带清晰明显,完整性较好,且无拖尾、弥散现象,这表明细菌的16S rRNA基因扩增成功。
2.2 不同条件巴氏杀菌乳中微生物多样性分析
2.2.1 序列丰度分析
在97%的相似水平上进行OTU序列分类,巴氏杀菌乳中总共聚成1 887 个OTU。依据划分的OTU,统计的序列数见表1,获得的多样性指数见图2。细菌OTU数量随着贮藏时间的延长而下降,在12 d后细菌种类有所增加。4、10 ℃条件下,细菌OTU数量下降速度较块,0 ℃下降较缓慢。由此可见,巴氏杀菌乳在0 ℃贮藏微生物多样性的变化较小,品质较为稳定。
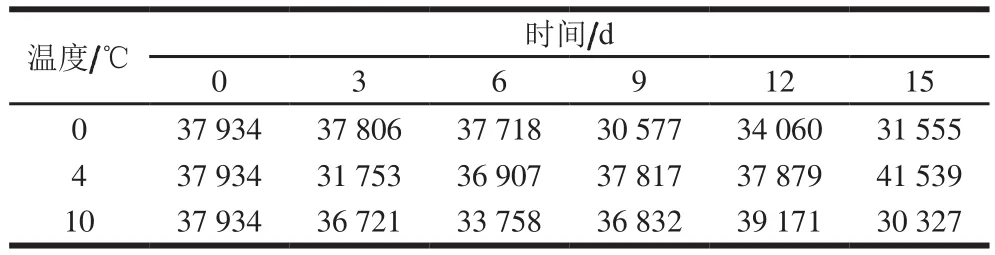
表1 不同条件巴氏杀菌乳的序列数Table 1 Sequence number of pasteurized milk at different storage temperatures for different times
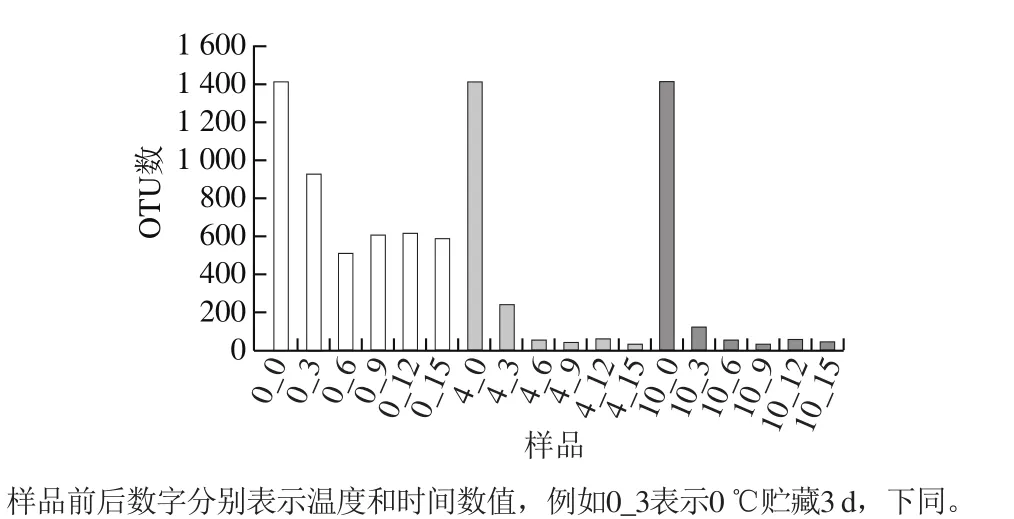
图2 不同条件巴氏杀菌乳的多样性指数Fig. 2 Diversity index of pasteurized milk at different storage temperatures for different times

图3 不同样品组巴氏杀菌乳的样本层级聚类分析Fig. 3 Cluster analysis of pasteurized milk samples in different groups
由图3可知,对不同贮藏条件的巴氏杀菌乳进行微生物层级聚类分析,发现其聚集程度各不相同。在0 ℃贮藏0~15 d内,巴氏杀菌乳的样本OTU层级聚类在一起,变化不明显,无明显差异。而在4 ℃贮藏3、6、9 d时,样本OTU层级与10 ℃贮藏不同时间条件下存在明显的交叉聚类情况,这种现象发生在3、6 d时较为明显。因此,猜测在4、10 ℃分别贮藏3、6 d时可能会影响微生物多样性发生一定的变化,值得进一步探讨和分析。

图4 不同贮藏温度、时间下巴氏杀菌乳的Shannon曲线Fig. 4 Shannon curves of pasteurized milk at different storage temperatures for different times
从图4可以看出,在低测序量时,Shannon多样性随着测序量的增加而增加;而且当测序深度达到200~300左右时,样品的Shannon多样性曲线趋势增长呈现缓慢趋势,接近水平状态,说明当前的测序量已经能够揭示出样品中的微生物组成。
2.2.2 巴氏杀菌乳门水平上细菌群落多样性分析
根据不同的贮藏温度和贮藏时间,从巴氏杀菌乳样品中共鉴定出7 个细菌门,分别为暂定螺旋菌门(Saccharibacteria)、拟杆菌门(Bacteroidetes)、放线菌门(Actinobacteria)、柔壁菌门(Tenericutes)、厚壁菌门(Firmicutes)、变形菌门(Proteobacteria)、未分类菌群(others)。相对丰度较高且样品均有分布的主要门类群有变形菌门、厚壁菌门。

图5 不同条件巴氏杀菌乳门水平细菌群落组成分析Fig. 5 Analysis of bacterial community composition at the phylum level in pasteurized milk at various storage temperatures for different times
不同的贮藏温度之间巴氏杀菌乳中细菌微生物菌群明显不同(图5),其中0 ℃贮藏的巴氏杀菌乳在门水平上细菌群落多样性较为丰富,几乎7 种细菌门皆有提及。随着贮藏时间的延长也可看出其各菌门的丰度有着明显波动,但并无明显变化。证明在0 ℃贮藏的巴氏杀菌乳在0~15 d内其门水平多样性无明显变化,此时贮藏较为稳定。4 ℃贮藏3 d时可看出暂定螺旋菌门消失,菌门的丰富度显著下降。在4、10 ℃贮藏6 d之后,在巴氏杀菌乳中,只剩下变形菌门、厚壁菌门两类门水平微生物,并以变形菌门为此时的主要优势菌群。
根据主成分分析以及巴氏杀菌乳门水平细菌群落组成的分析,发现0 ℃贮藏0~15 d内巴氏杀菌乳的微生物多样性的变化不明显,无明显差异。4、10 ℃其微生物多样性的变化有明显差异。由此,将进一步对贮藏温度0、4、10 ℃的巴氏杀菌乳细菌菌落属水平进行分析,从菌属的程度判断和分析贮藏条件对巴氏杀菌乳的品质影响。
2.2.3 巴氏杀菌乳属水平上细菌群落多样性分析

图6 0 ℃贮藏巴氏杀菌乳属水平细菌菌落分析Fig. 6 Analysis of bacterial community composition at the genus level in pasteurized milk stored at 0 ℃ for different times
如图6所示,在0 ℃不同贮藏时间条件下,巴氏杀菌乳样品中共鉴定出34 类细菌菌属,分别为支原菌属(Mycoplasma)、草酸杆菌属(Oxalobacteraceae)、假单胞菌属(Pseudomonas)、不动细菌属(Actinetobacter)、链球菌(Streptococcus)、Terrisporobacter、微球菌(Micrococcus)、Turicibacter、拟杆菌目S24-7组(norank_f_Bacteroidales_S24-7-group)、真细菌([Eubacterium]_rectale_grou)、肠球菌(Enterococcus)、Peptoclostridium、Faecalibacterium、寡养单胞菌属(Stenotrophomonas)、芽孢杆菌(Bacillus)、丛毛单胞菌科(unclassified_f_Comamonadaceae)、类芽孢杆菌(Paenibacillus)、疣微菌科UCG-005(Ruminococcaceae)、巨型球菌(Macrococcus)、鞘脂菌属(Sphingobium)、norank_p_Saccharibacteria、Paeniclostridium、Blautia、鞘氨醇单胞菌(Sphingomonas)、Peptostreptococcaceae、短波单胞菌属(Brevundimonas)、Clostriduium、嗜酸菌属(Acidovorax)、乳杆菌属(Lactobacillus)、金黄杆菌属(Chryseobacterium)、乳球菌属(Lactococcus)、Solobacterium、伯克霍尔德菌(Burkholderia)、Collinsella以及其他未分类菌属等。随着贮藏时间的延长可以看出支原菌属先缓慢下降,而后又呈现上升状态直到趋于稳定。但从整体来看,0 ℃贮藏的巴氏杀菌乳在0~15 d内其细菌菌落多样性及各细菌属丰度有明显的波动,但并无明显变化。

图7 4 ℃贮藏巴氏杀菌乳属水平细菌菌落分析Fig. 7 Analysis of bacterial community composition at the genus level in pasteurized milk stored at 4 ℃ for different times
与0 ℃巴氏杀菌乳细菌菌落种属多样性34 种(图6)相比,4 ℃贮藏巴氏杀菌乳种属多样性(图7)下降为21 种。大部分的菌属都已消失,在贮藏3 d时其优势菌群也由支原菌属、草酸杆菌属变成了假单胞菌属。同时此时还发现有新的菌属气单胞菌属(Aeromonas)的产生,并且其在6 d后开始发生显著的上升现象。随后,在贮藏9 d时发现有类芽孢杆菌属(Paenibacillus)、沙雷氏菌属(Serratia)出现的情况,并在贮藏12 d以后,其类芽孢杆菌属丰度显著上升。根据之前研究的理化结果[21],发现在此贮藏条件下的巴氏杀菌乳质地逐渐变黏稠,这可能是发生了腐败变质的现象。由此,多样性结果也进一步在此基础上判断出在3、 9 d有新的菌属的产生,这可能是其品质下降的关键点,推测假单胞菌属、气单胞菌属、芽孢杆菌属、沙雷氏菌属或许是导致巴氏杀菌乳品质衰败的关键因素。

图8 10 ℃贮藏巴氏杀菌乳属水平细菌菌落水平Fig. 8 Analysis of bacterial community composition at the genus level in pasteurized milk stored at 10 ℃ for different times
巴氏杀菌乳10 ℃贮藏3 d时细菌菌落属水平上多样性出现显著下降现象(图8),其变化与4 ℃贮藏大致相同(图7)。贮藏3 d时假单胞菌属丰度显著上升,并在此后随着贮藏时间的延长而下降,同时也发现了气单胞菌属的产生,其丰度在贮藏6 d开始大幅度上升。在贮藏9 d后也同样发现了之前几天未检测出的芽孢杆菌属、沙雷氏菌属以及变形菌属(unclassified_c_Gammaproteobacteria)的产生。结果表明,在10 ℃贮藏6 d时有新菌属的产生,结合之前理化的研究结果[21],推测此贮藏条件可能是影响品质腐败的关键裂变点此结果与4 ℃贮藏结果大致相同。故此,进一步推断在贮藏期间新产生的假单胞菌属、气单胞菌属、以及芽孢杆菌属、沙雷氏菌属可能是影响乳品发生腐败变质的潜在因素,今后也将在此基础上进一步研究和探讨,以此来证明实验结果的完整性和准确性。
3 讨 论
巴氏杀菌乳在我国的普及销售是国际形式的大势所趋,对巴氏杀菌乳贮藏条件的探索也是目前的主要研究方向[22]。有研究发现,在巴氏灭菌奶中存在潜在的腐坏微生物,这些微生物可能会在贮藏过程中影响产品质量[23-24]。本研究利用宏基因组测序技术对不同贮藏条件下巴氏杀菌乳的微生物多样性进行分析。研究表明,变形菌门、厚壁菌门这两类门水平微生物是巴氏杀菌乳贮藏期间的主要优势菌群。但是随着贮藏时间和温度的延长及升高,巴氏杀菌乳中的细菌多样性也在随之改变。目前,我国的巴氏杀菌奶产品货架期只有3~7 d。Elwell等[25]在2006年报道,商品巴氏杀菌奶在6 ℃的条件下可以贮藏16 d,而在0 ℃的条件下可以贮藏66 d。降低贮藏温度主要是增加了细菌生长的迟滞期和延缓了细菌的对数生长期,此结论与陈庆华等[26]的研究结果相同,随着贮存时间的延长微生物数量明显增加。本研究在此基础上还发现存在一些类芽孢菌属以及沙雷氏菌属,会导致乳品贮藏期间的腐败变质。研究结果表明,如若不能进行良好的贮藏,巴氏杀菌乳中残留的嗜冷菌、嗜热菌会导致乳品的腐败变质,进而影响产品的品质。
研究表明,耐热性细菌及嗜冷菌的巴氏杀菌后污染是限制货架期延长的主要原因[27]。大多数耐热性细菌不能在冷藏条件下生长,芽孢杆菌和类芽孢杆菌的菌株被认为是限制牛奶保质期常见的耐热嗜冷菌[27]。芽孢菌属在自然界中分布广泛,与包括牛奶在内的食品的污染和变质有关[28]。这些细菌可能在牛奶运输过程中以及在乳制品工厂通过加工设备而被污染[29]。芽孢杆菌形成内生孢子的能力给乳制品行业带来了挑战,如果环境条件允许,孢子可能会萌发成增殖细菌[30]。因此,有必要对在不同贮藏温度下存在的芽孢杆菌进行进一步的鉴定。本研究发现,随着温度的升高,芽孢杆菌属的OTU数量明显增加。相关研究也发现,巴氏灭菌后某些OTU的增加可能与梭菌目、芽孢杆菌目和乳酸菌目中的几个热成菌存活密切相关。同样,其他研究[31-32]也证实牛奶样品的成分差异也适用于芽孢杆菌属。
嗜冷菌同样也是影响原料乳及其制品质量和安全的关键微生物。假单胞菌是对原料乳危害最大的嗜冷菌之一,极易在冷藏条件下成为优势菌群[33]。在冷藏温度下,可能会增加假单胞菌和不动杆菌[12]的数量。本研究发现,巴氏杀菌乳在4、10 ℃条件下贮藏3 d时发现其优势菌群逐渐由支原菌属、草酸杆菌属变成了假单胞菌属,与此同时还出现了气单胞菌属,在6 d以后显著增加,这与贮藏的温度密切相关。嗜冷菌在生长代谢过程中,分泌的耐热性蛋白酶和脂酶,极易导致原料乳在高温灭菌后发生凝胶、沉淀、变苦和酸败现象[34]。由此可见,巴氏杀菌乳在不当的贮藏条件下会产生对人体致病的菌株。此外,研究还发现在4、10 ℃贮藏9 d发现有沙雷氏菌菌株的出现,该细菌有能力在冷藏条件下生长,同样也是导致牛奶发生变质的主要原因。随着基因和生物信息技术的进步,新型检测方法对嗜冷菌在原料乳生产、贮藏和加工中的危害研究更加深入细致。开发快速、灵敏并能评估嗜冷菌以及胞外酶的危害性的检测方法,仍然是需要解决的问题。
4 结 论
本研究利用宏基因组学高通量测序的方法对不同贮藏温度下巴氏杀菌乳的多样性指数和群落结构进行了分析研究,共检测到7 个细菌门,34 类细菌菌属。在0 ℃贮藏的巴氏杀菌乳微生物多样性保持最完整;而4、10 ℃贮藏时是微生物多样性显著变化的关键点,对其巴氏杀菌乳的微生物多样性及营养品质都有着较大影响,菌群构成及优势菌群都发生了变化,贮藏温度越高影响越大。同时,在4 ℃贮藏3、9 d,10 ℃贮藏6 d下巴氏杀菌乳品质腐败的关键时间点都出现了之前未被检测出的类芽孢菌属、沙雷氏菌属,推测此两种菌属是导致巴氏杀菌乳品质腐败的关键决定因素。
