丙二醛氧化对米糠蛋白结构及功能性质的影响
2019-07-05周麟依孙玉凤
周麟依,孙玉凤,吴 非*
(东北农业大学食品学院,黑龙江 哈尔滨 150030)
我国米糠资源丰富,年产量达1 800万 t。米糠中蛋白质约占20%。米糠蛋白中可溶性组分约占七成,近似于大豆蛋白。米糠蛋白所含有的必需氨基酸组成相对完整,接近于联合国粮农组织/世界卫生组织推荐模式。米糠蛋白中的赖氨酸含量比大米胚乳、小麦面粉等谷物蛋白的含量相对要高,并且米糠蛋白的生物效价接近于乳酪蛋白,消化率高于90%,因此被认为是一种高营养价值的植物蛋白。
蛋白质的氧化是由活性氧(reactive oxygen species,ROS)直接诱导或者由氧化应激反应的次级副产物间接诱导反应。氧化过程产生的这种结构修饰使蛋白质结构发生显著变化,如主链断裂、分子交联、分子解折叠以及构象改变。目前,可诱导蛋白质氧化的促氧化剂种类较多,如分子氧受到光活化转化为激发单线态氧(1O2),或减少一个电子形成超氧阴离子自由基(O2-·)。而脂肪氧化也可促使蛋白质氧化,脂肪氧合酶会催化大豆油、米糠油等植物油氧化,在此过程中会产生大量的烷氧自由基以及醛、酮等活性次生氧化产物,这些氧化产物氧化活性较高,进一步诱导了蛋白质氧化。米糠含有活性较强的脂肪水解酶及脂肪氧化酶,它们可水解脂质产生游离脂肪酸,从而导致米糠酸败[1]。此过程中产生的游离脂肪酸容易氧化形成脂质自由基和脂质活性产物,进而诱导米糠蛋白氧化,严重影响了米糠蛋白的功能性[2-4]。
目前,脂质过氧化被定义为多不饱和脂肪酸或脂质的氧化变质。脂质过氧化是由一个自由基的复杂链反应所引起。多不饱和脂肪酸受自由基的氧化还原夺氢反应生成脂质烷基自由基,而后通过分子重排作用形成共轭二烯结构,继而共轭二烯结构在脂质烷基自由基上进一步通过氧化反应逐步形成结构稳定性相对较差的脂质氢过氧化物[5]。脂质氢过氧化物失稳后逐渐转化为活性羰基化合物[6]。对比于脂质自由基及脂质氢过氧化物,活性醛分子具有更稳定的结构,并可通过更大的分子扩散作用对蛋白产生更大程度的氧化攻击[7]。最具代表性且最易造成氧化损伤的脂质过活性产物主要是丙烯醛、4-羟基-2-壬烯醛及丙二醛(malondialdehyde,MDA),并且MDA是脂质过氧化反应最易形成的活性醛,尽管MDA的氧化反应活性弱于丙烯醛,但由于分子内存在两个羰基,可通过与蛋白质之间发生更大程度共价交联影响蛋白质结构及功能性[8-9]。当蛋白质和过氧化脂质接触反应时,过氧化脂质亦可通过疏水相互作用或氢键作用与蛋白复合生成脂蛋白复合体[10-12]。在脂质氧化反应生成的多种活性醛均可与氨基发生醛醇缩合作用进而形成色素物质[13-15],甚至严重影响酶活性及蛋白质生物效价及营养健康功能[16-18]。
目前,MDA对部分植物蛋白结构及功能的影响已有研究,但MDA氧化对米糠蛋白结构及功能的研究较少。因此,选MDA代表脂质过氧化反应中的活性次生氧化产物,研究脂质过氧化产物对米糠蛋白结构和功能性质以及构效关系的影响。本研究有助于全面深入了解米糠蛋白的氧化机理,为米糠蛋白产品的合理研发提供思路,促进米糠蛋白产业的发展。
1 材料与方法
1.1 材料与试剂
米糠蛋白由实验室自制;1,1,3,3-四甲氧基丙烷(1,1,3,3-tetramethoxy propane,TMP)、盐酸胍 美国Sigma公司;Lowry法蛋白质含量测定试剂盒 上海荔达生物科技有限公司;二硝基苯肼 天津博迪化工股份有限公司;其他试剂均为分析纯。
1.2 仪器与设备
F-4500荧光分光光度计 日本Hitachi公司;MAGNA-IR560傅里叶变换红外光谱系统 美国尼高力公司;PHSJ-4A型实验室pH计 中国上海雷磁公司;XW-80A旋涡混合器 上海青浦沪西仪器厂。
1.3 方法
1.3.1 米糠蛋白提取
取原料米糠过60 目筛,按照1∶10(g/mL)加入正己烷,室温下不断搅拌脱脂4 h,反复脱脂2~3 次,在4 000 r/min离心10 min,上清液用蒸馏法回收正己烷,脱脂后将米糠置于通风橱挥发48 h,再将脱脂米糠过60 目筛,取10 g放入100 mL蒸馏水中,用0.1 mol/L NaOH溶液调节溶液pH 9.5,在50 ℃条件下搅拌2 h,然后在3 000 r/min离心20 min,将沉淀去除,再用0.1 mol/L HCl溶液调节溶液pH 3.8后3 000 r/min离心20 min,将去除上清液,沉淀即为米糠蛋白,将米糠蛋白用蒸馏水洗脱两次并最终调节pH 7.0,并进行冻干备用。
1.3.2 MDA氧化米糠蛋白的制备
参考Adams等[9]方法通过水解TMP制备MDA。将10.0 mL 5.0 mol/L盐酸加入31.6 mL蒸馏水中充分混匀后加入8.4 mL TMP,快速移入避光水浴锅中在40 ℃条件下进行30 min氧化反应,通过隔时搅拌处理防止团聚物产生,TMP水解处理后迅速用6 mol/L氢氧化钠调节反应溶液pH 7.6,继而利用0.05 mol/L pH 7.4的磷酸盐缓冲液定容TMP水解液至1 L。通过摩尔吸光系数31 500 L/(mol·cm)精确定量MDA的浓度,为了保证MDA的氧化活性,反应储液需要现配现用。
为制备MDA氧化诱导米糠蛋白,利用0.01 mol/L、pH 7.6的磷酸盐缓冲溶液调配米糠蛋白溶液至质量浓度为10 mg/mL,并加入0.5 mg/mL NaN3防止蛋白腐败变性,别用0.001、0.01、0.1、1 mol/L的MDA溶液作为氧化诱导剂加入的米糠蛋白溶液中,经过隔日避光25 ℃恒温条件下连续振荡进行氧化处理,诱导米糠蛋白发生MDA氧化。氧化诱导后米糠蛋白溶液迅速进行4 ℃冷浴处理,之后转移至低温离心机中30 min二次离心处理,留存上清液通过低温透析处理后滤去未参与反应的MDA,剩余蛋白组分经过24 h冷冻干燥后即可制备为MDA诱导氧化米糠蛋白。
1.3.3 米糠蛋白羰基含量测定
蛋白质氧化程度主要利用Huang Youru等[19]羰基含量的测试分析方法进行测试。1.3.2节制备的氧化米糠蛋白充分溶解于去离子水中,经过2 h充分搅拌后进行4 ℃低温10 000×g离心30 min,通过Lowery法确定上清液中氧化蛋白质量浓度,通过稀释调控上清液中氧化蛋白质量浓度为3~6 mg/mL。
将上述0.35 mL的氧化蛋白溶液加入1 mL 2 mol/L HCl溶液中(含有10 mmol/L 2,4-二硝基苯肼),在室温条件下水浴处理2 h。另取0.35 mL氧化米糠蛋白溶液加入1 mL不含2,4-二硝基苯肼的2 mol/L HCl溶液中,在相同处理条件进行实验作为空白对照组。实验组及对照组离心管中分别加入0.45 mL 40%的三氯乙酸溶液快速充分混合后低温10 000×g离心20 min,除去上清液,保留沉淀组分利用乙醇与乙酸乙酯的等体积溶液进行多次洗涤离心,保留最终沉淀蛋白。将收集蛋白溶解入1.0 mL 0.1 mol/L含6 mol/L盐酸胍的pH 7.0磷酸盐缓冲液,通过37 ℃水浴处理20 min,通过隔时快速摇晃防止聚沉现象的发生。以空白组为参比在367 nm波长处进行数值矫正,最终以22 000 L/(mol·cm)的消光系数计算蛋白质羰基含量。
1.3.4 米糠蛋白游离氨基的测定
400 mg邻苯二甲醛(1,2-phthalic dicarboxaldehyde,OPA)试剂充分溶解于1 mL甲醇溶液中,而后依次向溶液中加入预先配制的质量浓度为200 g/L十二烷基硫酸钠(sodium dodecyl sulfate,SDS)溶液2.5 mL及浓度为0.1 mol/L的硼酸溶液25 mL,继而转移至通风橱内加入100 μL的β-巯基乙醇,最后将溶液用蒸馏水定容至50 mL,制备成OPA溶液用于后续检测分析,量取OPA试剂4 mL与200 μL氧化米糠蛋白样品充分混匀后进行35 ℃水浴处理2 min,以蒸馏水空白组为对照在340 nm波长处测定吸光度。
1.3.5 米糠蛋白游离巯基和二硫键含量测定
利用经典的5,5’-二硫双(2-硝基苯甲酸)(5,5’-dithiobis(2-nitrobenzoic acid),DNTB)比色法对蛋白游离巯基、总巯基及二硫键含量进行分析测定。取150 mg氧化米糠蛋白加入10 mL 0.1 mol/L含有l mmol/L乙二胺四乙酸(ethylene diamine tetraacetic acid,EDTA)和1% SDS的pH 8.0的磷酸盐缓冲液A中,充分混合均匀后8 000 r/min离心30 min,利用Lowery法对上清液中氧化米糠蛋白含量进行定量。为精确测定游离巯基含量,量取3 mL上述氧化米糠蛋白溶液加入磷酸盐缓冲液A后,滴入0.1 mL DNTB溶液(39.6 mg DNTB溶解于10 mL磷酸盐缓冲液A中制备得到),充分振荡混合后进行室温1 h水浴处理,水浴处理后溶液经30 min的低温高速离心处理取上清液,通过参比DNTB空白组,在412 nm波长处测定吸光度,以13 600 L/(mol·cm)消光系数计算游离巯基含量。精确测定氧化米糠蛋白中总巯基基团含量,取1 mL氧化米糠蛋白溶液,依次加入0.05 mL β-巯基乙醇及4 mL 0.1 mol/L尿素-盐酸胍混合溶液(5∶8,V/V),充分混合后进行室温1 h水浴处理后,将10 mL 12 g/100 mL三氯乙酸加入溶液中进行室温水浴处理1 h,再次以4 500 r/min离心10 min获取沉淀物,重复上述步骤两次后,将最终沉淀物溶液于最后将沉淀溶解在10 mL 0.1 mol/L其中含有l mmol/L EDTA和1% SDS pH 8.0磷酸盐缓冲液中,再加入0.08 mL DNTB试剂,剧烈振荡摇匀溶液后在室温条件下进行水浴处理1 h,再加入10 mL 12 g/100 mL的三氯乙酸,室温条件下二次水浴处理1 h,将混合溶液中速低温离心处理10 min。蛋白沉淀物再次分散于20 mL 12%的三氯乙酸溶液中,充分离心弃去β-巯基乙醇组分,重复3 次上述处理后低温高速离心30 min。以不加DNTB的空白对比组进行参比,测定上清液412 nm波长处的吸光度,以13 600 L/(mol·cm)消光系数计算氧化米糠蛋白中总巯基含量。
1.3.6 米糠蛋白傅里叶红外光谱分析
为有效消除蛋白质荧光背景,利用BRAVO拉曼光谱仪对大豆蛋白-磷脂酰胆碱纳米乳液进行拉曼分析测试,测试条件为功率80 mW、激发光波长532 nm、曝光时间60 s,扫描300~3 500 cm-1范围内的波数谱段,每个样品都重复扫描3 次以上,由计算机做信号累加平均并绘图输出,峰位误差控制在±3 cm-1内。
1.3.7 米糠蛋白内源荧光光谱分析
将氧化米糠蛋白分散溶解于中性磷酸盐缓冲溶液中,调控氧化米糠蛋白质量浓度为0.15 mg/mL。荧光分光光度计以米糠蛋白所含的色氨酸基团为荧光探针,为避免酪氨酸的荧光干扰,设置荧光光谱的激发波长为290 nm,荧光发散光谱的扫描范围设置为300~400 nm,激发狭缝和发射狭缝宽设置为7.5 nm[20]。
1.3.8 米糠蛋白表面疏水性测定
参考Kato等[21]的8-苯胺-1-萘磺酸(8-anilino-1-naphthalenesulfonic acid,ANS)荧光探针法测定氧化米糠蛋白表面疏水性并进行适当修正。用0.1 mol/L中性磷酸盐缓冲溶液充分溶解米糠蛋白,10 000×g高速离心处理30 min除去沉淀物,以Lowery法分析测试上清液中蛋白质量浓度,通过磷酸盐缓冲液的逐步稀释,调控氧化米糠蛋白溶液质量浓度为0.05~0.4 mg/mL,取40 μL浓度为8 mmol/L的ANS溶液滴加至不同质量浓度的米糠蛋白溶液4 mL,经振荡混匀后静置3 min,然后进行荧光强度测试,测试条件为激发波长λex为390 nm,发射波长λem为468 nm,扫描夹缝5 nm,扫描速率10 nm/s。将荧光强度与蛋白质量浓度作线性图,以初始段的斜率值计为氧化米糠蛋白的表面疏水性大小。
1.3.9 米糠蛋白凝胶电泳分析
参考Laemmli[22]凝胶电泳测试方法对氧化米糠蛋白并作适度调整。分离胶和浓缩胶分别为12%、 5%。将氧化米糠蛋白样品与上样缓冲溶液混合配制蛋白质量浓度为1 mg/mL,上样缓冲溶液的配制通过加入100 mg SDS、0.25 mL β-巯基乙醇、2 mg溴酚蓝、2 mL pH 8.0的0.05 mol/L Tris-HCl溶液、2.0 mL甘油,最后定容至10 mL。在95 ℃水浴条件下加热5 min。蛋白凝胶电泳的上样量为10 μg。电泳测试过程中调控浓缩胶电压为80 V,分离胶电压为120 V。以甲醇、冰乙酸、0.1%考马斯亮蓝R-250配制染色剂侵染蛋白电泳胶,并以甲醇、冰乙酸、水配制为脱色剂用于脱除蛋白电泳胶的底色背景。
1.3.10 米糠蛋白溶解度测定
参考Shimada等[23]的方法。准确称取20 mg氧化米糠蛋白样品充分溶解于10 mL蒸馏水中,在室温条件下充分搅拌1 h,高速离心20 min除去沉淀。以Lowry法测定上清液中蛋白质质量浓度,以牛血清白蛋白为标准物绘制标准曲线。以上清液质量浓度与米糠蛋白质量浓度的比例表示米糠蛋白溶解度。
1.3.11 米糠蛋白粒径分布测定
利用Mastersizer 2000激光光散射仪对米糠蛋白的粒径分布进行分析测定。不同氧化诱导条件处理下的米糠蛋白样品溶解于去离子水中并配制溶液质量浓度为1 mg/mL,在室温条件下充分搅拌3 h,隔夜处理后保证氧化米糠蛋白充分溶解,用激光光散射仪进行米糠蛋白的粒径分布测试,折射率为1.330[24]。
1.3.12 米糠蛋白浊度测定
将氧化米糠蛋白用磷酸盐缓冲液稀释40 倍,以磷酸盐缓冲液为空白对照,用紫外分光光度计测定600 nm波长处的吸光度,浊度计算公式为:

式中:A为稀释乳液在600 nm波长处的吸光度;V为稀释倍数;I为光程差0.01 m。
1.3.13 米糠蛋白乳化性和乳化稳定性测定
取1.0 mL植物油和3.0 mL 0.2%米糠蛋白磷酸盐缓冲溶液,混合振荡并进行高速均质处理制备为均一乳液,从底部抽取50 μL样品,用5 mL 0.1% SDS溶液稀释,在500 nm波长处测吸光度A0。静置30 min后再此从底部各取50 μL样品,用5 mL 0.1% SDS溶液稀释,测吸光度A30,同溶度SDS为空白。按式(2)、(3)计算米糠蛋白乳化性和乳化稳定性:
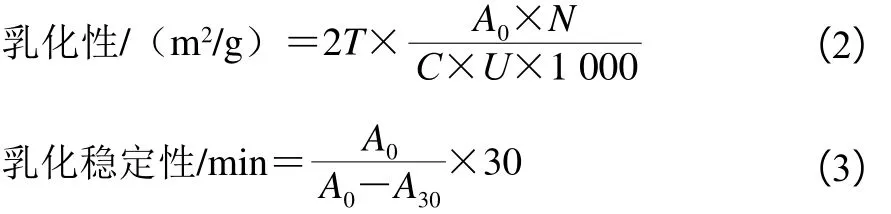
式中:T为浊度(2.303);N为稀释倍数250;C为乳化液形成前蛋白质溶液中蛋白质量浓度/(g/mL);U为乳化液中油的体积分数(0.25);A0为0 min的吸光度。
1.3.14 米糠蛋白起泡性和泡沫稳定性测定
准确称量0.2 g米糠蛋白样品置于已加入0.05 mol/L pH 7.0磷酸盐缓冲液20 mL的50 mL烧杯中。使用高速分均质机以10 000 r/min均质30 s,连续3 次约30 min,测均质后体积(V0),静置30 min后再此读取液面体积V30。起泡性和泡沫稳定性参见公式(4)、(5):

1.4 数据处理与统计分析
每个实验均进行3 次重复平行实验,利用SPSS Statistics 22软件对数据进行ANOVA差异显著性分析,用Tukey's HSD法进行ANOVA差异性检验,P<0.05,差异显著。采用Origin 9.1软件、OMNIC软件包、PeakFit 4.12软件分析等进行数据分析,图表处理及图谱分析处理。
2 结果与分析
2.1 氧化对米糠蛋白羰基含量的影响

图1 MDA氧化诱导对米糠蛋白羰基含量的影响Fig. 1 Effect of MDA concentration on carbonyl content of rice bran protein
氨基酸侧链上的NH-或NH2-基团极易受到羟自由基的攻击,进而氧化生成新的羰基基团[25-26]。如图1所示,氧化米糠蛋白羰基含量随着MDA浓度增大逐渐增加。这是由于MDA可与蛋白质分子中的伯胺以等比例的加成反应形成烯胺加合物,从而为蛋白分子引入了新的羰基基团。此外,研究发现MDA可作用于半胱氨酸、组氨酸及赖氨酸等氨基酸残基的亲核侧链基团进而反应生成席夫碱诱导蛋白羰基化[27-28]。另外,由于MDA结构的特殊性,在MDA分子上连接有两个羰基,因此MDA分子中的一个羰基氧化作用连接于蛋白质的可同时引入另外一个羰基基团[9]。Burcham等[29]研究发现MDA氧化诱导牛血清白蛋白的羰基值随着MDA氧化诱导浓度的增大及氧化诱导时间的延长而逐渐增加。
2.2 MDA氧化对米糠蛋白二级结构的影响

图2 MDA氧化米糠蛋白的红外光谱Fig. 2 FTIR spectra of MDA oxidation of rice bran protein

表1 MDA氧化对米糠蛋白二级结构的影响Table 1 Effect of oxidative modification by malondialdehyde on secondary structure contents of rice bran protein%
如图2和表1所示,随着MDA氧化诱导浓度的增加,米糠蛋白α-螺旋结构、β-折叠结构以及反平行式β-折叠结构含量逐渐增加,无规卷曲和β-转角结构含量逐渐下降。α-螺旋结构、β-折叠结构及反平行式β-折叠结构逐渐增加,这主要是与氧化米糠蛋白分子间疏水作用和静电作用增大诱导了蛋白分子聚集有关,此外MDA诱导氧化米糠蛋白的共价交联聚集也是可能原因。Sun Weizheng等[30]研究发现肌原纤维蛋白β-折叠含量随加工时间的延长氧化程度的增大而增加,并且氧化聚集与反平行式β-折叠结构逐渐增加有关。无规卷曲和β-转角结构逐渐降低可能是因为部分无规卷曲和β-转角结构转变为β-折叠结构。
2.3 MDA氧化对米糠蛋白内源荧光光谱的影响
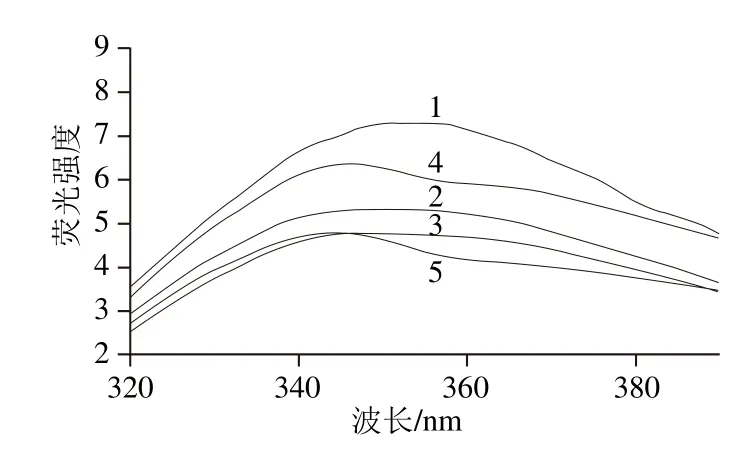
图3 MDA氧化诱导的米糠蛋白内源荧光谱图Fig. 3 Intrinsic fluorescence of rice bran protein oxidized by MDA
如图3所示,随着MDA浓度的增加,氧化米糠蛋白内源荧光λmax呈现逐渐蓝移的趋势。研究发现,MDA氧化会使兔肌球蛋白、α-晶状体球蛋白以及牛血清白蛋白的内源荧光λmax蓝移的现象[31-33],并且Traverso等[33]认为MDA诱导氧化蛋白质内源荧光强度下降主要是由于MDA诱导氧化聚集包埋了色氨酸的原因。在酸性条件MDA易与蛋白色氨酸上的吲哚基团发生反应进而猝灭色氨酸荧光[34],而由于在本研究中所采用反应条件近中性,故而排除了MDA与色氨酸吲哚基团反应引起的荧光猝灭的可能性,造成米糠蛋白内源荧光强度下降应当主要与氧化米糠蛋白的共价交联聚集有关。Sun Wenzheng等[35]研究表明荧光强度的变化可用来表征蛋白质氧化情况以及结构变化,乳液荧光强度显著降低表明蛋白质的结构逐渐展开。叶林[5]研究发现随着氧化浓度的增加,花生蛋白的荧光强度逐渐下降,表明蛋白质色氨酸残基被转移到更加疏水的非极性环境中。
2.4 MDA氧化对米糠蛋白巯基和二硫键含量的影响
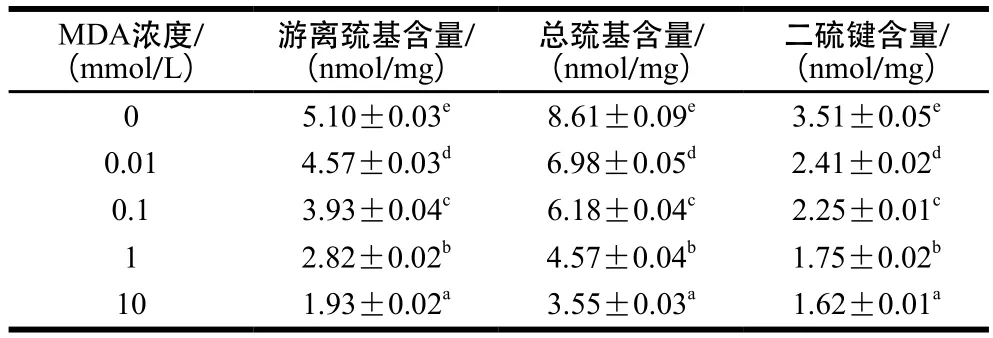
表2 米糠蛋白样品游离巯基、总巯基和二硫键的含量Table 2 Free sulfhydryl, total sulfhydryl and disulfide bond contents of rice bran protein samples oxidized by MDA
由于巯基对氧化的极高敏感性,蛋白氧化最早期现象主要为巯基的氧化[36]。如表2所示,随着MDA浓度的增加,二硫键、游离巯基和总巯基含量逐渐降低,这可能是由于自由基活性较高,氧化米糠蛋白二硫键逐渐断裂,内部的巯基虽然随着蛋白质结构解折叠而暴露,但自由基迅速与其结合形成亚磺酸和磺酸等氧化产物,使二硫键含量,游离巯基和总巯基含量不断降低。
2.5 MDA氧化对米糠蛋白游离氨基含量的影响

图4 MDA氧化对米糠蛋白游离氨基含量的影响Fig. 4 Effect of MDA oxidation on free amine content of rice bran protein
蛋白侧链上带有的氨基酸均易受到自由基和非自由基ROS攻击。如图4所示,MDA氧化诱导使米糠蛋白游离氨基含量随着氧化浓度的增加逐渐减少,MDA分子中的羰基可直接与米糠蛋白上的氨基发生共价交联反应,进而降低了米糠蛋白的游离氨基含量。
2.6 MDA氧化对米糠蛋白溶解度的影响
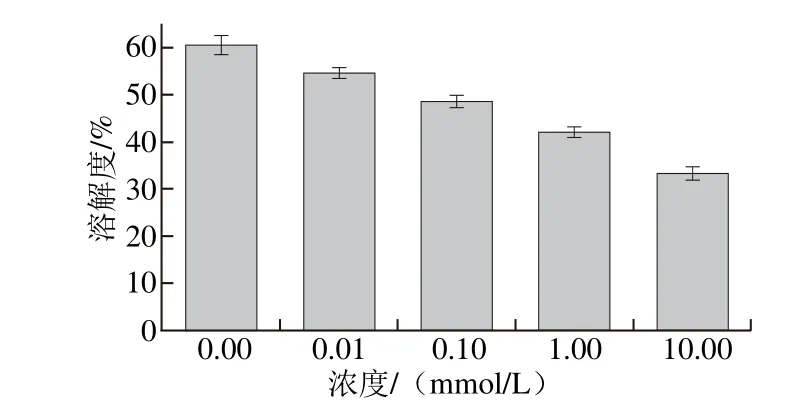
图5 MDA氧化对米糠蛋白溶解度的影响Fig. 5 Effect of MDA oxidation on solubility of rice bran protein
如图5所示,随着MDA氧化诱导浓度的增加,氧化米糠蛋白溶解度呈现逐渐下降的趋势。这主要是由于氧化米糠蛋白发生了共价交联聚集,不可溶性的交联聚集体形成降低了米糠蛋白的溶解性。刘晶等[37]研究表明低浓度MDA氧化诱导下大豆蛋白趋于可溶性聚集,而在高浓度MDA氧化诱导下大豆蛋白共价交联聚集体逐渐形成,同时诱导了可溶性聚集体的再聚集,进而形成了不可溶性蛋白聚集物,为大豆蛋白溶解性随氧化诱导浓度增大逐渐降低的原因。
2.7 MDA氧化对米糠蛋白电泳结果的影响

图6 MDA氧化诱导米糠离蛋白的电泳图Fig. 6 Electrophoresis profiles of rice bran protein with MDA
米糠蛋白是由清蛋白、球蛋白、醇溶蛋白和谷蛋白组成的。尽管现有研究已利用对不同电泳条带进行了提取和分析,但米糠蛋白中4 种主要蛋白的亚基归属尚无统一定论。Adebiyi等[38]研究指出米糠白蛋白主要亚基的分子质量约为31~45 kDa,米糠球蛋白主要亚基的分子质量约为33 kDa和53 kDa。Hamada[39]研究表明的米糠白蛋白和球蛋白的分子质量分别为10~100 kDa和10~150 kDa。Wang Changyuan等[40]研究发现米糠白蛋白的主要亚基分布于14~32 kDa分子质量区间,而米糠球蛋白主要亚基的分子质量为63、53、49、36、22 kDa,米糠谷蛋白的主要亚基的分子质量为54、37、21、13 kDa,而醇溶蛋白的主要亚基分子质量为13 kDa和22 kDa。
如图6所示,对比已有研究可以认定米糠蛋白的主要亚基的分子质量分别为62、53、49、36、25、21、18、14 kDa,米糠白蛋白主要亚基的分子质量为36 kDa和14 kDa,分子质量为62、53、49、36、21 kDa的电泳条带归属于米糠球蛋白亚基,米糠谷蛋白主要亚基的分子质量为53、36、21 kDa和14 kDa,醇溶蛋白主要亚基的分子质量分别为21 kDa和14 kDa。低氧化诱导浓度下MDA对米糠蛋白亚基组成无显著影响,随着MDA氧化诱导浓度增加,分子质量为53、49、36、21、14 kDa的亚基归属条带均有所变浅,条带逐渐变窄。MDA分子中含有两个羰基,当两个羰基与不同多肽链间的氨基同时形成席夫碱时就会导致蛋白质共价交联。由此可知,分子质量为53、49、36、21 kDa和14 kDa的亚单位产生了不可溶性交联聚集。而随着氧化诱导浓度的增加,分子质量为36、21、14 kDa的亚基逐渐消失,表明蛋白交联聚集主要发生于上述亚基,亚基条带的消失也是米糠蛋白功能性降低的重要原因。
2.8 MDA氧化对米糠蛋白表面疏水性的影响
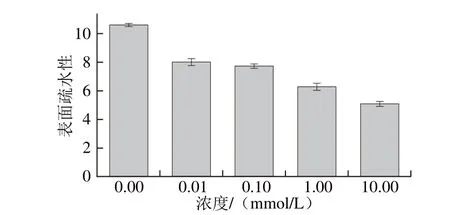
图7 MDA氧化对米糠蛋白表面疏水性的影响Fig. 7 Effect of oxidation by MDA on surface hydrophobicity of rice bran protein
如图7所示,随着MDA氧化诱导浓度的增加,米糠蛋白表面疏水性逐渐下降。这可能是由于氧化米糠蛋白分子结构展开,暴露出来的巯基和疏水基团不断发生氧化性修饰,并通过共价交联和疏水相互作用等途径形成氧化聚集体。Boatright等[41]通过大豆蛋白氧化结构及功能在不同抗氧化剂添加条件下的变化研究中发现,蛋白氧化伴随着表面疏水性的降低。Liang[42]研究发现即使避光条件下60 ℃的油脂热氧化也可诱导大豆蛋白氧化进而降低了其表面疏水性。黄友如[43]在大豆蛋白的LOX-亚油酸氧化诱导体系下,大豆蛋白的氧化程度随诱导反应时间延长逐渐增大,表面疏水性也随之逐渐降低。研究发现,在大米蛋白氧化模拟体系中发现随着反应时间的延长,大豆蛋白氧化程度持续增加,表面疏水性逐渐下降[44]。吴伟等[45]研究发现随着米糠保藏时间延长,米糠发生了氧化酸败诱导谷蛋白分子结构变化,氧化聚集体逐渐形成进而降低了蛋白表面疏水性,印证了本研究的结论。
2.9 MDA氧化对米糠蛋白粒径的影响

图8 MDA氧化诱导米糠蛋白的粒径分布Fig. 8 Particle size distribution of MDA-oxidized rice bran protein

表3 米糠蛋白样品的D43和PDITable 3 D43and PDI of oxidized rice bran protein samples
蛋白质氧化诱导的分子交联及聚集作用下蛋白质分子粒径趋于变大,通过测定蛋白质粒径分布(图8)能直观地看出蛋白质聚集或降解作用。随着MDA浓度的增加,分子平均粒径均逐渐增大。通过粒径分布图可知,天然米糠蛋白粒径主要呈现双峰分布,其中大蛋白粒径分布峰归属于氧化聚集,而氧化米糠蛋白平均粒径的增加主要与蛋白聚集体峰的增大有关。由此可以得知,无论低浓度或高浓度蛋白氧化条件下米糠蛋白均发生了氧化聚集效应。但氧化聚集与其他氧化作用对蛋白结构的影响效应相互叠加,是米糠蛋白结构及功能变化的最终原因。而随着米糠蛋白氧化程度的增加,氧化米糠蛋白粒径逐渐再聚集形成更大的聚集体。通过粒径的变化可知,高浓度MDA氧化诱导条件下米糠蛋白除发生不可溶性交联聚集外,同样发生了可溶性分子聚集。
多分散指数(polydispersity index,PDI)是衡量溶液分布的均匀程度。蛋白质PDI值越小说明蛋白质溶液越均一,PDI值越大说明溶液中存在蛋白质大小聚集体的分布差距越大。如表3所示,米糠蛋白PDI值随MDA氧化剂浓度的增加而增大,表明米糠蛋白溶解状态下分子粒径分布均一性变差,这主要是由于氧化聚集体所致。汪建明等[46]认为乳蛋白的氧化聚集体分为大粒度聚集体(约1 000 nm)和小粒度聚集体(约100 nm),氧化作用可诱导低粒度聚集体再聚集形成大粒径聚集体,大粒径聚集体也解离形成低粒度颗粒,两种行为同时存在。Promeyrat等[47]认为蛋白质粒径变化主要与蛋白质分子交联及聚集有关。黄友如[43]研究发现脂质氧化诱导的蛋白质氧化过程中蛋白质结构解折叠疏水基团外露为蛋白聚集提供了疏水聚集作用力,更多有序二级结构转化为无序结构过程中氢键的重组及分子间范德华力也为蛋白聚集提供了聚集作用条件,受多种分子间交互作用力的诱导,氧化蛋白发生了多重聚集现象,进而形成了大粒径的蛋白聚集体,是粒径增大的重要诱因。
2.10 氧化对米糠蛋白质浊度的影响
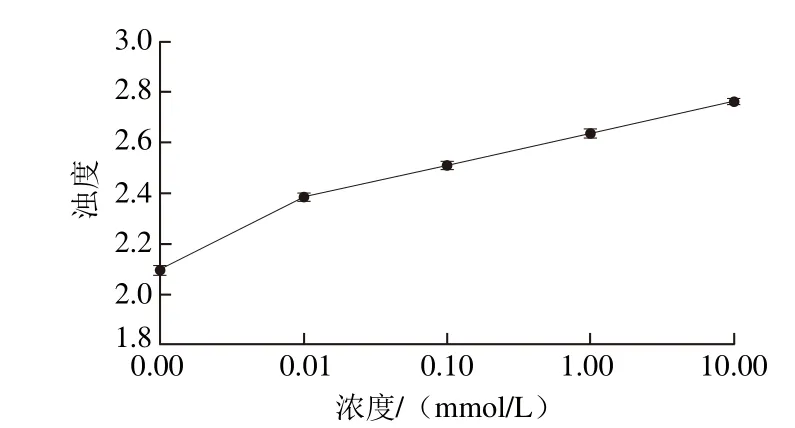
图9 MDA氧化对米糠蛋白浊度的影响Fig. 9 Effect of MDA oxidation on turbidity of rice bran protein
蛋白质溶液浊度可以反映蛋白质的聚集程度,可用于侧面衡量蛋白质相互作用大小及聚集体的相对尺寸[48-49]。由图9可知,随着氧化剂MDA浓度的增加浊度均逐渐增大,这与氧化作用促进米糠蛋白的亚基分子聚集有关,浊度的增加与前述的氧化米糠蛋白粒径增大相互验证。MDA氧化的米糠蛋白浊度增大与交联聚集体及分子聚集体形成有关。黄友如[43]通过脂肪氧合酶催化亚油酸诱导大豆蛋白聚集机理研究中发现,氧化作用诱导了大豆蛋白发生结构解折叠,暴露出更多的疏水基团,并通过疏水相互作用、静电作用或氢键等构建形成较大的聚集体,增加了大豆蛋白的浊度,并伴随着流体力学半径分布范围的扩大。李艳青[50]研究表明羟自由基氧化的鲤鱼蛋由浊度随氧化程度增大浊度逐渐提高。Cui Xuhai等[51]研究发现与天然乳清蛋白相比,经过3、5、10 h羟自由基氧化处理的乳清蛋白浊度分别增加16.9%、22.9%和24.3%。牛思思[52]研究发现,氧化蛋清蛋白的浊度有所增加,并且随着蛋清蛋白氧化程度的增大,蛋清蛋白的浊度逐渐增加。由此可知,氧化聚集普遍发生于蛋白氧化过程中。
2.11 MDA氧化对米糠蛋白质乳化性和乳化稳定性的影响
如图10所示,随着MDA浓度的增高,氧化米糠蛋白乳化性和乳化稳定性呈现逐渐降低的趋势。这可能是因为随着MDA浓度的增大,米糠蛋白表面疏水性逐渐降低,米糠蛋白发生疏水性聚集,并且由于活性醛会与蛋白质发生Schiff碱反应生成交联聚集,氧化米糠蛋白形成可溶和不可溶性聚集体均降低了蛋白分子舒张及乳化界面稳定作用,故米糠蛋白的乳化能力和乳化稳定性均呈现下降趋势。研究发现,低浓度MDA诱导的大豆蛋白氧化因结构部分解折叠,更多的疏水基团外露增加了大豆蛋白的分子结构柔性,提高了大豆蛋白分子油水界面的吸附结合能力,增加了大豆蛋白的乳化功能,另外大豆蛋白在低浓度丙烯醛氧化诱导下溶解性增加也是乳化性提高的促进因素;而高浓度MDA氧化诱导下大豆蛋白团聚为不可溶性聚集体降低了分子柔性,减少了界面吸附结合面积,降低了大豆蛋白的乳化功能[53]。对比可知,米糠蛋白由于分子交联作用强于界面舒张解折叠是其乳化功能降低的重要原因。

图10 MDA氧化对米糠蛋白乳化性和乳化稳定性的影响Fig. 10 Effect of MDA oxidation on emulsifying capacity and emulsion stability of rice bran protein
2.12 MDA氧化对米糠蛋白起泡性和泡沫稳定性的影响

图11 MDA氧化对米糠蛋白起泡性及泡沫稳定性的影响Fig. 11 Effect of MDA oxidation on foaming capacity and foam stability of rice bran protein
由图11可见,随MDA氧化诱导浓度的增加,米糠蛋白的起泡性逐渐降低,这主要是由于虽然两种蛋白氧化作用均部分解开了米糠蛋白分子结构,分子内部的疏水残基更多地暴露于蛋白表面,但此条件下过强的米糠蛋白疏水作用促进了分子间聚集,且由于活性醛会与蛋白质反应生成希夫碱则促进了不可溶性交联聚集体的形成,两种聚集作用均抑制了米糠蛋白在水气界面扩展和吸附,降低了米糠蛋白起泡性。目前尚难以解释MDA氧化诱导米糠蛋白泡沫稳定性随氧化诱导浓度增大而增强的原因,刘晶等[37]认为部分暴露的疏水性基团可相互吸引,是MDA氧化诱导蛋白泡沫稳定性增强的主要原因。
3 结 论
MDA是食品中脂质过氧化反应中产生最多的活性醛类,由于MDA含有两个羰基,在一定条件下MDA也可使得蛋白质形成共价交联[8-9],使蛋白质的功能特性降低。本研究中,米糠蛋白羰基含量均随MDA氧化诱导浓度的增加而增大,这主要是由于MDA主要与蛋白质分子中的伯胺加成反应形成烯胺加合物,从而为蛋白质引入羰基基团。MDA分子含有两个羰基,当其中一个羰基与蛋白质反应时,也会同时为蛋白质引入另一个新的羰基。米糠蛋白α-螺旋和β-折叠含量随MDA氧化诱导浓度增加逐渐增大,β-转角结构和无规卷曲结构随之呈现逐渐降低的变化趋势,表明氧化使米糠蛋白无序结构转变为有序结构,这应与氧化诱导的聚集作用有关。随着MDA氧化诱导浓度的增加,米糠蛋白表面疏水性逐渐下降,荧光λmax发生蓝移,表明MDA氧化诱导促进了米糠蛋白氧化聚集,并随着氧化诱导浓度的增大氧化聚集程度逐渐增加。低氧化诱导浓度下MDA对米糠蛋白亚基组成无显著影响,随着MDA氧化诱导浓度增加,分子质量为53、49、36、21、14 kDa的亚基归属条带均有所变浅,条带逐渐变窄。而随着氧化诱导浓度的增加,分子质量为36、21、14 kDa的亚基逐渐消失,亚基条带消失可能与大聚集体的形成及重组有关。通过随氧化诱导浓度增大氧化米糠蛋白溶解性的逐渐降低及粒径分布范围的增大可知,MDA诱导的米糠蛋白氧化聚集并存了可溶性及不可溶性聚集形式,氧化蛋白的自聚集及交联聚集分别形成可溶性及不可溶性蛋白聚集体,并可以通过荧光分析及表面疏水性分析判断,氧化聚集是MDA诱导米糠蛋白氧化的主导行为。
结合考虑随MDA氧化诱导浓度增大米糠蛋白溶解性降低、粒径增大、浊度增大的变化规律可知,MDA诱导的氧化米糠蛋白可溶性聚集体及不可溶性聚集体含量均随着氧化浓度的增大而增加,可溶性聚集的粒径逐渐增加可能是由于蛋白亚基更大程度的聚集所致或新的氧化聚集形成,而不可溶性氧化聚集的增加也可能是由于可溶性聚集体再聚集所致。在本研究中由于氧化交联形成的不可溶聚集体降低了米糠蛋白的界面延展性,使乳化性、稳定性均有所降低。
