抑郁倾向对个体情绪调节目标的影响——来自事件相关电位的证据*
2019-06-17杨小光郑文瑜
李 红 杨小光 郑文瑜 王 超
抑郁倾向对个体情绪调节目标的影响——来自事件相关电位的证据
李 红杨小光郑文瑜王 超
(深圳大学教育学部心理学院;深圳市情绪与社会认知科学重点实验室;深圳市神经科学研究院, 深圳 518060)
目前抑郁症情绪失调的研究主要关注策略的选择和应用, 但是对于情绪调节目标是否异常及其背后的电生理基础尚不清楚。情景选择是成熟的情绪反应产生之前运用的一种调节策略, 可以反映情绪调节目标。本研究要求抑郁倾向被试与控制组被试观看并选择快乐、中性和悲伤场景图片, 同时记录事件相关电位(ERP)和主观情绪偏好。结果发现, 抑郁倾向组悲伤图片的LPP波幅显著小于健康控制组, 并更多地选择再次观看悲伤图片。此外, 抑郁倾向组对于悲伤情绪的偏好显著增加, 对快乐情绪的偏好则显著降低。结果提示抑郁倾向个体对快乐刺激的趋近动机和对悲伤刺激的回避动机都降低, 从而使得抑郁倾向被试的情绪调节目标为更多地体验到悲伤。
情绪调节; 情绪调节目标; 抑郁倾向; 趋避动机; 晚期正成分
1 引言
情绪是人们生活的晴雨表, 有效控制自身情绪发生的时间、强度以及表达情绪的方式对我们的身心健康尤其重要。情绪是个体对外部和内部事物产生的主观体验, 是一种可被调节的反应倾向。情绪调节则是个体以情绪作为调整的对象, 对当前情绪的发生、体验与表达的影响(Gross, 1998), 达到个体希望获得的情绪体验。因此情绪调节的结果取决于个体想要达到的情绪状态(情绪调节目标)和改变情绪所采用的方法(情绪调节策略)。例如要想降低负性情绪体验, 人们就需要直接在降低负性情绪的方向做出努力, 并且使用适当有效的情绪调节策略来达到目标。
情绪调节目标决定了情绪调节的方向。一般来说, 人们的情绪调节目标是调节情绪使快乐体验最大化, 也就是要体验正性、积极情绪, 回避负性、消极情绪(Tamir, 2009)。情绪调节策略则决定了如何在所选的方向上有效地改变情绪, 人们可以运用多种调节策略来达成特定的情绪调节目标。根据情绪调节过程模型(Gross, 1998, 2015), 在完全成熟的情绪反应产生之前运用的情绪调节策略更为有效, 而情景选择策略就是其中一种人们经常或首先使用的调节策略。它指的是个体选择有可能改变情绪的刺激(电影、图片等)以调节自身情绪, 因此情绪调节的目标决定了个体对刺激类型的选择。如果人们想要体验积极、正性的情绪, 就会在情景选择过程中选择可以诱发正性情绪的刺激, 反之亦然。因此情景选择可以反映情绪调节的目标, 而且个体一般倾向于选择能够诱发正性情绪的情景和回避诱发负性情绪的情景(Harmon-Jones, Harmon-Jones, Amodio, & Gable, 2011; Millgram, Joormann, Huppert, & Tamir, 2015; Tamir & Ford, 2012)。
情绪失调是抑郁症患者的核心特征之一, 表现为长时间的抑郁情绪、丧失快乐等(Aldao, Nolen- hoeksema & Schweizer, 2010)。目前有关抑郁症及抑郁倾向被试情绪失调的研究大多集中于研究这些个体在选择何种策略以及如何有效地应用该策略上是否存在异常。研究发现, 与正常人相比, 抑郁症病人更频繁地采用不良策略(比如沉思和表达抑制)而更少采用适应性策略(比如认知重评和分心) (Aldao et al., 2010; Garnefski & Kraaij, 2006; Joormann & Gotlib, 2010)。而对于抑郁症病人运用特定情绪策略能力的研究显示, 当让抑郁症病人自发地选择情绪调节策略并执行时, 无法有效调节负性情绪; 但当对他们进行指导后, 他们运用认知重评和表达抑制进行情绪调节的能力与正常人并没有显著差异(Ehring, Tuschen-Caffier, Schnülle, Fischer & Gross, 2010)。但是很少有研究关注情绪失调与情绪调节目标之间的关系, 特别是抑郁倾向被试与正常人的情绪调节目标的差异。虽然Millgram等(2015)已经提示抑郁症病人与正常人的情绪调节目标有差异, 但这种差异潜在的神经生理学基础目前尚不清楚。
晚期正成分(late positive potential, LPP)作为事件相关电位(event related potentials, ERPs)的一个重要成分, 大约在刺激呈现300 ms之后出现, 是研究情绪反应的神经生理学指标(Ito, Larsen, Smith, & Cacioppo, 1998)。在抑郁症的情绪信息加工领域, 抑郁症病人以及抑郁程度较高的个体都对正性和负性刺激表现出更弱的LPP波幅(Admon & Pizzagalli, 2015; Hajcak, MacNamara & Olvet, 2010; MacNamara, Kotov & Hajcak, 2016; Proudfit, Bress, Foti, Kujawa & Klein, 2015)。而且Weinberg和Hajcak (2010)近期的研究发现, 抑郁症个体观看奖励性和威胁性刺激时都表现出更弱的LPP波幅。此外, LPP可以作为表征情绪性刺激的动机性注意以及动机强度的指标(Lang & Bradley, 2010; Schupp et al., 2000)。有研究表明对正性刺激的趋近动机越强则这些刺激诱发的LPP波幅越大(Briggs & Martin, 2009a; Weinberg & Hajcak, 2010), 动机强度越高的正性刺激越能够诱发更大的LPP波幅(Gable & Harmon-Jones, 2013)。同样地, 对负性刺激的回避动机越强也会诱发更大的LPP波幅, 如威胁性刺激和残肢断臂的图片会比损失和污染的场景诱发更大的LPP波幅(Leutgeb, Schäfer & Schienle, 2009; Michalowski et al., 2009)。这些研究都表明了LPP波幅反映了情绪刺激诱发的趋避动机的强度, 因此LPP可以作为较好的表征趋避动机强度的生理指标(Lang & Bradley, 2010; Schupp et al., 2000)。而且以往与动机相关的理论认为个体的目标, 包括情绪调节目标(Millgram et al., 2015), 是由趋近和回避动机诱发的(Elliot & Thrash, 2002)。
综上所述, 本研究应用事件相关电位(ERP)技术考察抑郁倾向被试与健康控制组被试对快乐、中性和悲伤场景图片的加工以及随后对这些图片的选择情况, 在此过程中记录两组被试的脑电信号。另外, Tamir (2009)等认为对情绪的偏好也可以反映被试的情绪调节目标。因此, 我们将在实验中让被试评估自己对快乐和悲伤这两种情绪的偏好, 作为情绪调节目标的另一个指标。根据已有文献提供的证据, 本研究对实验结果提出以下预期:1)抑郁倾向组被悲伤图片诱发的LPP波幅小于健康控制组, 这表征了抑郁倾向组对悲伤图片的回避动机更弱, 可能造成他们选择再次观看的悲伤图片多于健康控制组;2)抑郁倾向组被快乐图片诱发的LPP波幅同样小于健康控制组, 这表征了抑郁倾向组对快乐图片的趋近动机更弱, 可能造成他们选择再次观看的快乐图片少于健康控制组。通过本研究可获得抑郁倾向个体对快乐、悲伤和中性三种类型图片进行加工的神经生理证据, 从而加深我们对抑郁症情绪失调的神经机制的了解, 同时找到更多的情绪相关的生物标记物以帮助我们对抑郁倾向个体及重度抑郁个体进行干预和治疗。
2 方法
2.1 被试
40名广东省某大学在校大学生(男女各20名)参加了实验, 其中健康控制组20名(年龄:20.8 ± 1.79岁, 男性11人, 女性9人), 抑郁倾向组20名(年龄:20.6 ± 1.98岁, 男性9人, 女性11人)。两组被试均为右利手, 视力或矫正视力正常。每个被试都在实验前签订了相应的知情同意书。被试入组标准(采用二次筛选法):首先, 在广东省某高校公选课上给本科生发放1100份贝克抑郁量表(Beck Depression Inventory-II, BDI-Ⅱ), 剔除信息以及量表条目填写不完整的问卷。根据量表得分, 筛选出BDI-Ⅱ≥ 19分的大学生定义为抑郁倾向初筛组, BDI ≤ 13分的大学生定义为健康对照初筛组。之后在1~2周内通过电话的方式邀请符合以上抑郁量表初筛标准的43名大学生(23名抑郁倾向初筛, 20名健康对照初筛)进入结构式访谈。依据精神障碍诊断与统计手册DSM临床定式访谈(The structured clinical interview for DSM, SCID), 由一名经过训练的研究生对其进行诊断与鉴别诊断。通过SCID定式临床检查, 排除归于丧离者(症状归于近期遭遇的重大应激创伤事件(如亲人离世等)); 排除双向情感障碍、重性抑郁障碍、焦虑障碍、脑器质性精神障碍; 排除具有精神分裂症等重性精神疾病史或家族史个体; 并排除定期服用药物或者使用其他非药物性影响中枢神经系统的物质与神经系统疾病史个体。SCID-I是用以对DSM-IV轴I的大多数障碍确定诊断的半定式检查,既可用于精神专科患者的诊断, 也可用于在普通医疗部门就诊的患者, 或者是那些并不认为自己患有精神疾病的个体, 如在社区精神疾病普查中用于对精神病患者家属的调查。SCID-I的交谈方式和诊断范围适用于成人(18岁以上)。经过访谈, 排除3名正在服药的抑郁症患者, 最终得到抑郁倾向组和健康控制组被试各20人。
2.2 实验材料及程序
2.2.1 材料
首先, 在中国情绪图片系统(Chinese affective picture system, CAPS) (白露, 马慧, 黄宇霞, 罗跃嘉, 2005), 国际情绪图片系统(International affective picture system, IAPS) (Lang et al., 1997)以及网络上选择快乐图片75张, 中性图片75张, 悲伤图片100张。然后, 正式实验前邀请25名健康的大学生对这些图片的效价、唤醒度及其诱发的具体情绪以9点量表进行评定。经过评定, 选择符合实验要求的快乐、悲伤、中性图片各60张, 总计180张图片作为正式实验的材料。所有的图片都以相同的亮度和对比度呈现在灰色背景上(中央呈现, 3.0°×3.5°)。
对所选图片效价和唤醒度进行单因素方差分析, 结果表明, 不同类型图片的效价有差异,(2, 177) = 445.45,< 0.001, η= 0.83。Tukey事后检验表明快乐图片的效价(6.47 ± 0.90)显著高于中性图片(5.17 ± 0.34)和悲伤图片(3.06 ± 0.51), 所有< 0.001, 而中性图片的效价显著高于悲伤图片,< 0.001。分析结果显示不同类型图片的唤醒度也有差异,(2, 177) = 234.40,< 0.001, η= 0.73。快乐图片的唤醒度(5.29 ± 1.06)与悲伤图片的唤醒度(5.15 ± 0.80)没有显著差异,= 0.662, 并且都显著大于中性图片的唤醒度(2.37 ± 0.56), 所有< 0.001。另外, 被选择的快乐图片能有效地诱发快乐情绪(快乐 = 5.95 ± 0.91), 而悲伤图片能够有效地诱发悲伤情绪(悲伤 = 5.47 ± 0.87)。
2.2.2 程序
为了测定被试的实验当时的情绪, 我们要求被试填写正负性情绪量表(Positive and Negative Affect Schedule, PANAS, 5点评分, 20个项目), 然后进行图片选择任务, 在任务进行之前先让被试练习6个trials (快乐、悲伤、中性各2个trials), 紧接着开始正式图片选择任务。
图片选择任务的程序如图1所示。首先, 屏幕中间会出现注视点“+”1000 ms, 然后, 每张图片会在屏幕上呈现1500 ms, 三种类型的图片随机呈现。被试的任务是观看图片后决定他们是否喜欢刚呈现的图片中的场景。接着, 被试按“1”则表示喜欢该场景, 图片会再一次呈现2000 ms, 或者按“2”表示不喜欢该场景, 会出现黑色屏幕2000 ms。在实验的后半段, 按键设置与前一半trials相反, 即按“1”看黑色屏幕2000 ms, 按“2”则重新看图片2000 ms。按键设置做被试内平衡。
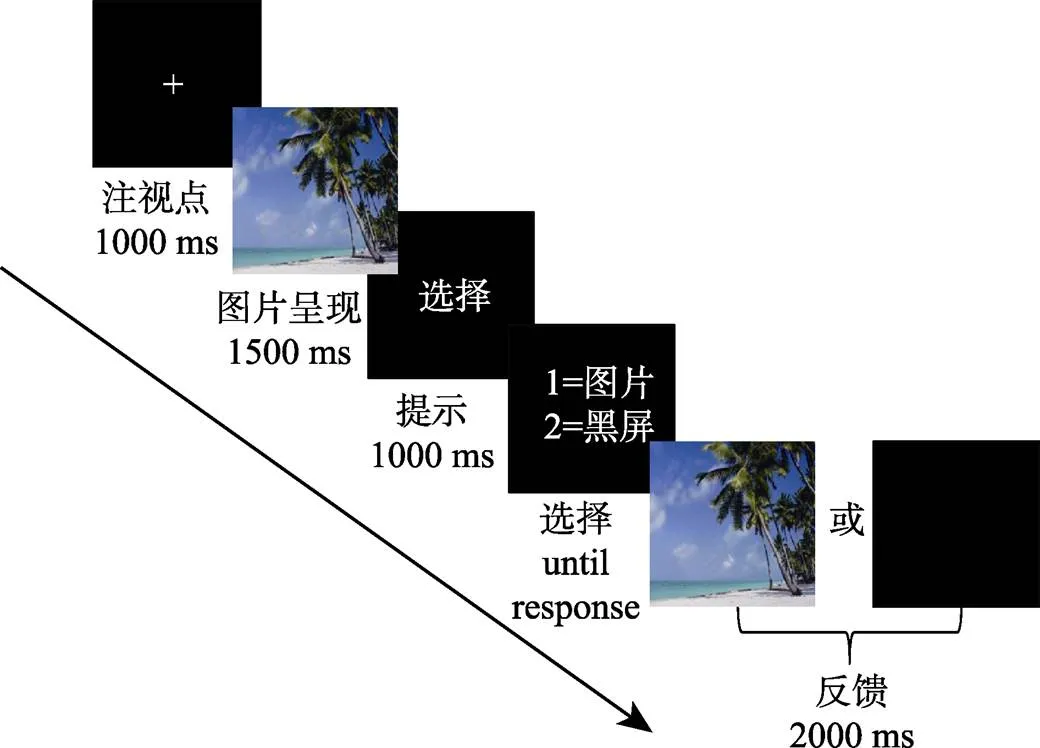
图1 图片选择流程图
被试完成图片选择任务之后, 每张图片会再次呈现2 s, 要求被试按键评估每张图片让他们感受到的快乐和悲伤程度(9点评分, 1非常微弱, 9为非常强烈, 分数越高表明情绪体验越强)。完成图片评价任务后, 被试还需要报告他们平常想要体验的快乐和悲伤程度(7点评分, 1 = 非常微弱, 7 = 非常强烈), 以此来反映他们对快乐和悲伤的偏好程度。
2.3 数据采集及分析
使用64导脑电放大器(Brain Products, Gilching, 德国)采集脑电和眼电数据, 电极的阻抗低于5 kΩ。在线记录时以FCz为参考电极, 离线转为双侧乳突平均参考。在线右眼正下方安置电极记录垂直眼电, 离线后选用FT9为水平眼电。脑电中的水平和垂直眼电利用ICA方法去除。
数据分析采用Brain Vision Analyzer 2.1。脑电数据依次经过以下处理:滤波(滤波带通0.01~30 Hz, 零相位延迟)、分段(−200至1500 ms)、基线矫正(−200至0 ms)、叠加平均。本研究分析的ERP成分包括中央−额区的P2, 前额−中央的N2, 以及中央−顶区的LPP。由于早期和中期的ERP成分具有较尖锐的峰值, 因此对P2和N2采用基线−峰值进行度量; 而晚期的LPP波形较钝, 故采用平均波幅进行度量。为了使ERP结果具有更高的信噪比, 每个ERP成分由该成分波幅最大的3至4个电极点的平均值计算而来。根据已有相关文献, 脑电地形图以及ERP波形图选取电极点和ERP成分的分析时间窗(Dennis & Hajcak, 2009; Feng et al., 2014; Hajcak et al., 2010; Yang, Zhu, Wang, Wu, & Yao, 2011), 具体而言, P2成分的分析电极点为POz, PO3, PO4, Oz, O1, O2, 峰值检测窗口为180~230 ms。N2成分的分析电极点为Fz, FC3, FCz, FC4, Cz, 峰值检测窗口为200~300 ms。LPP成分的分析电极点为CPz, CP1, CP2, 平均波幅计算窗口为600~1200 ms。脑电数据分析中我们剔除了参考点双侧乳突(TP9和TP10)数据无法收集以及波幅大于±100 μV的被试4人(其中抑郁倾向组2人, 健康控制组2人), 最后纳入数据分析的被试为36名。
2.4 统计
统计分析采用SPSS Statistics 20.0 (IBM, Somers,美国)。我们将利用独立样本t检验研究两组被试实验当天的正性情绪和负性情绪状态是否存在差异, 其中组别(抑郁倾向组、健康控制组)为自变量, PANAS评分为因变量。之后我们采用双因素重复测量方差分析研究图片类型(快乐、中性、悲伤)和组别对图片的快乐和悲伤的情绪体验程度, 图片选择数量, 脑电成分P2、N2和LPP幅度的影响, 其中图片类型为被试内变量, 组别为被试间变量。此外, 我们还以组别为被试间变量, 以情绪(快乐、悲伤)为被试内变量进行重复测量方差分析, 考察两组被试对快乐和悲伤的情绪偏好程度是否有差异。最后, 我们采用相关分析的方法来考察图片选择和情绪偏好等关键变量之间的相关关系。
3 结果
3.1 行为
3.1.1 被试基本信息和主观情绪评定
抑郁倾向组的BDI-Ⅱ评分(25.11 ± 4.05)显著高于健康控制组(5.22 ± 3.08),(34)= 16.10,< 0.001; 两组被试在年龄((34) = 0.350.73)和性别((1,= 36) = 0.44,= 0.505)上都没有显著差异(见表1)。此外, 对两组被试的PANAS评分进行分析, 发现抑郁倾向组的正性情绪(19.77 ± 7.46)与健康控制组(20.33 ± 6.29)没有显著差异,(34)= 0.24,= 0.239。同样, 抑郁倾向组的负性情绪(12.72 ± 2.35)与健康控制组(11.38 ± 4.09)也没有显著差异,(34) = −1.20,= 0.811。
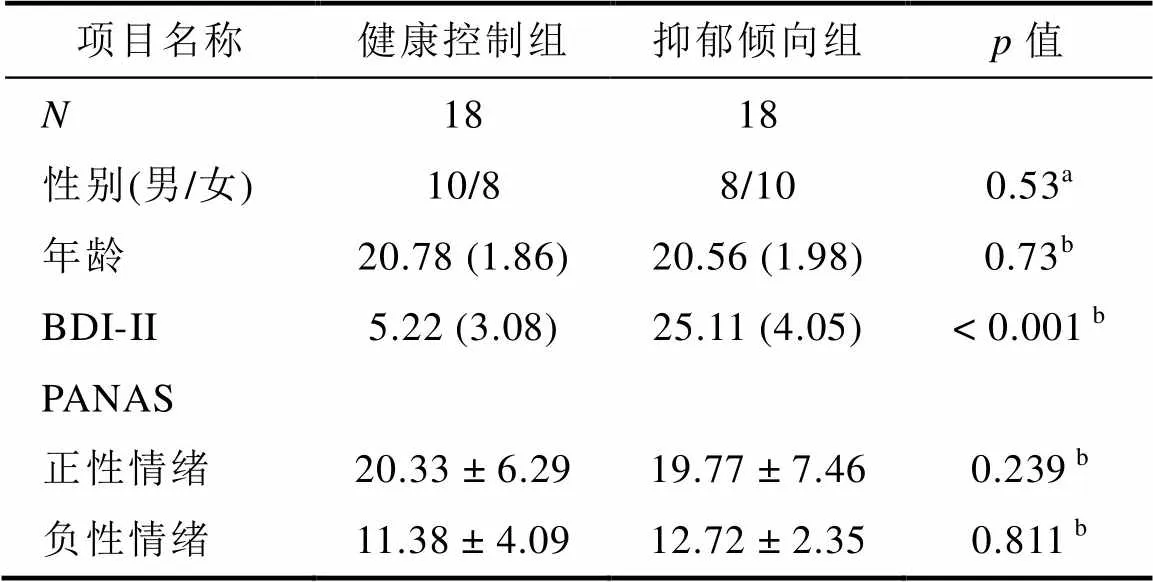
表1 被试的人口学信息和临床特征
注:表中呈现真值或平均值(标准差)。BDI-II = Beck Depression Inventory-Ⅱ (贝克抑郁量表第二版)。表示进行了卡方检验;进行了双样本检验。
我们对被试评定的三种类型图片的快乐程度进行分析, 球形检验结果表明被试内变量不符合球形假设,(2,= 36) = 13.18,0.001。因此我们使用Greenhouse-Geisser方法校正, 图片类型的主效应显著,(1.51, 51.15) = 148.51,< 0.001, 偏η= 0.81。组别主效应不显著,(1, 34)= 1.10,= 0.302。组别与图片类型的交互效应不显著, Greenhouse- Geisser adjusted(1.51, 51.15) = 1.82,= 0.180。事后检验现快乐图片诱发的快乐程度(5.62 ± 1.01)显著高于中性图片(2.17 ± 1.06)和悲伤图片(1.32 ± 0.36) (中性:(35) = 17.50,< 0.001; 悲伤:(35) = 22.93,< 0.001), 而中性图片诱发的快乐程度显著高于悲伤图片,(35) = 5.09,< 0.001 (见表2)。
对三种类型图片诱发的悲伤程度进行分析, 球形检验结果表明被试内变量不符合球形假设,(2,= 36) = 34.63,0.001。Greenhouse-Geisser方法校正结果显示, 图片类型的主效应显著,(1.21, 41.22) = 245.84,< 0.001, 偏η= 0.88。组别主效应不显著,(1, 34) = 0.18,= 0.68。组别与图片类型的交互效应不显著, Greenhouse-Geisser adjusted(1.21, 41.22) = 1.17,= 0.729。事后检验发现悲伤图片诱发的悲伤程度(5.42 ± 1.67)显著高于中性图片(1.59 ± 0.75)和快乐图片(1.32 ± 0.35) (中性:(35) = 16.93,< 0.001; 快乐:(35) = 16.07,< 0.001), 而中性图片诱发的悲伤程度显著高于快乐图片,(35) = 2.77,0.009 (见表2)。
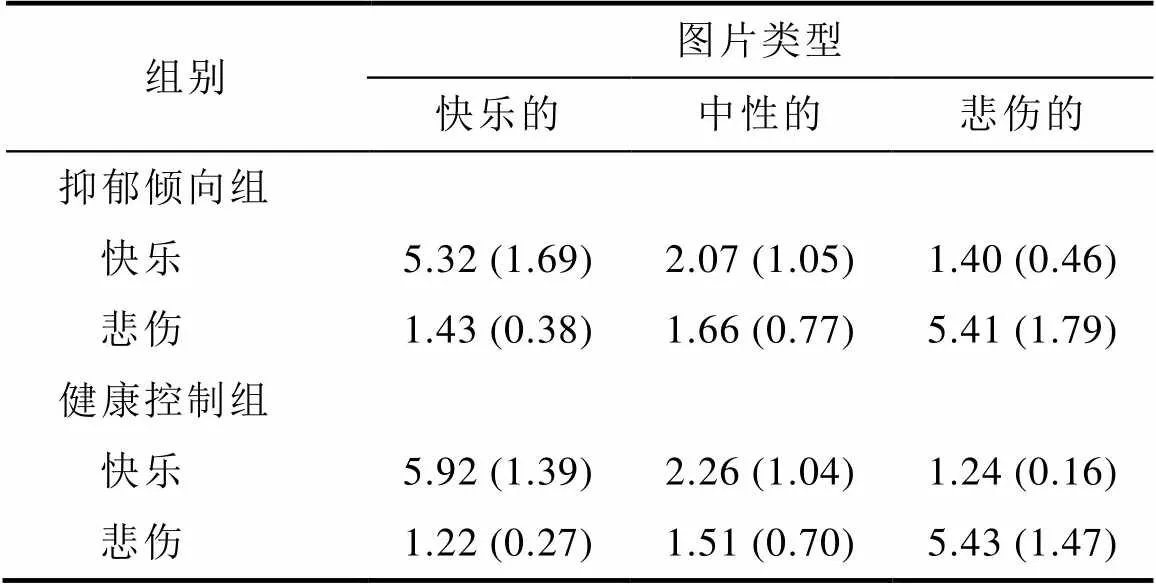
表2 两组被试对三种图片的情绪反应
注:表中呈现均值, 括号内为标准差。
3.1.2 图片选择
为了检验两组被试对悲伤图片和快乐图片的选择是否有差异, 本研究以组别(抑郁倾向组, 健康控制组)为被试间变量, 以图片类型(快乐、中性、悲伤)为被试内变量进行重复测量方差分析。分析结果显示被试内变量不符合球形假设,(2,= 36) = 7.11,0.029。Greenhouse-Geisser方法校正结果显示组别与图片类型交互效应显著,(1.68, 56.96) = 6.63,= 0.004, 偏η= 0.16 (见图2)。进一步在图片类型的每个水平上的简单效应分析发现, 对于再次观看悲伤图片的数量, 抑郁倾向组显著多于健康控制组,(34) = 2.95,0.006。而对于再次观看快乐图片和中性图片的数量, 抑郁倾向组选择与健康控制组没有显著差异; 对于快乐图片,(34) = 2.89,0.097; 对于中性图片,(34) = 0.75,0.457。在抑郁倾向组, 被试选择再次观看快乐图片的数量显著多于悲伤图片,(17) = 2.75,0.014, 且他们选择再次观看快乐图片和悲伤图片的数量与中性图片都没有显著差异; 对于快乐图片,(17) = 2.49,0.023; 对于悲伤图片,(17) = 1.46,0.163。在健康控制组, 被试选择再次观看的快乐图片数量都显著多于中性图片和悲伤图片; 对于中性图片,(17) = 4.52,0.001; 对于悲伤图片,(17) = 8.32,0.001。并且选择再看的中性图片数量显著多于悲伤图片,(17) = 5.77,0.001 (见表3)。
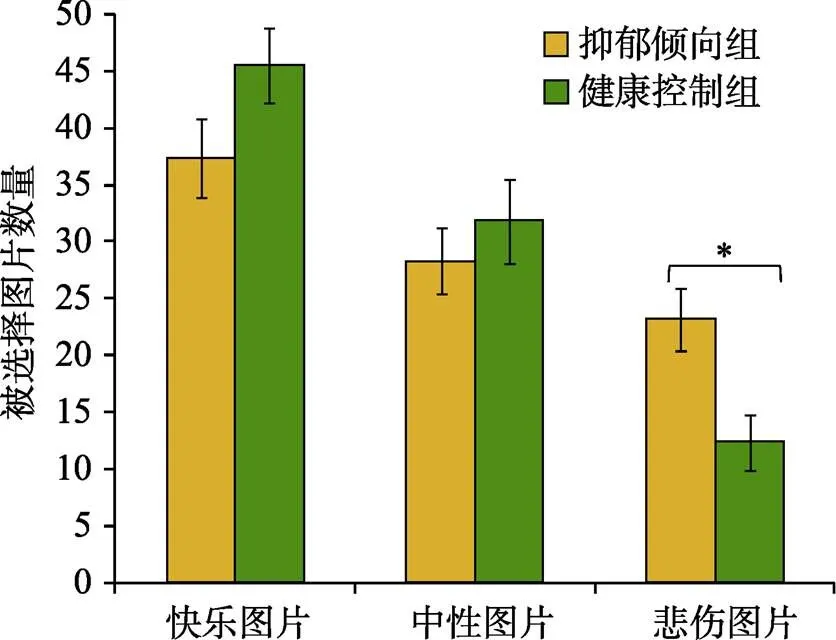
图2 两组被试对三种图片(快乐、中性和悲伤)的选择, 纵轴表示被试选择再看一遍的图片数量。*表示p < 0.05。
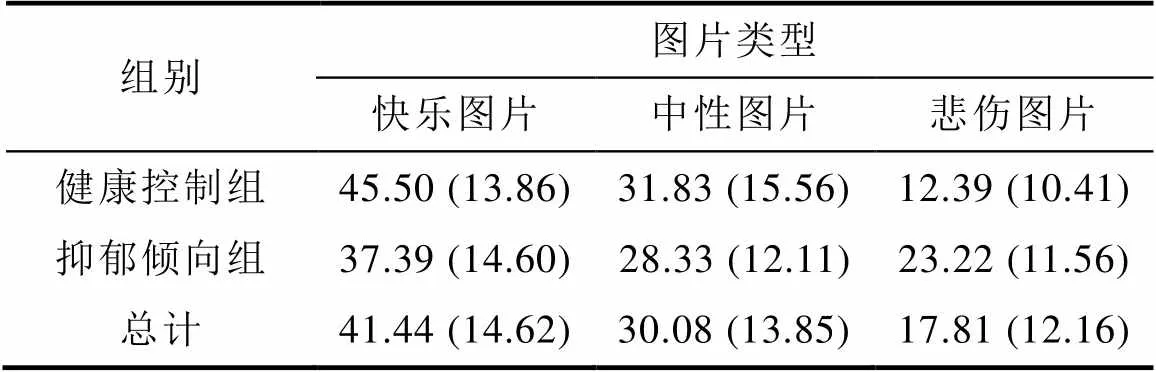
表3 两组被试对三种图片的选择
方差分析表明图片类型主效应显著, Greenhouse- Geisser adjusted(1.68, 56.96) = 37.97,0.001, 偏η= 0.53。对图片类型的主效应事后分析表明, 两组被试选择的快乐图片的数量(41.44 ± 14.62)明显多于中性图片(30.08 ± 13.85)和悲伤图片(17.81 ± 12.16) (中性:(35) = 4.81,0.001; 悲伤:(35) = 6.59,0.001); 而且所选择的中性图片的数量要明显多于悲伤图片,(35) = 4.57,0.001。组别主效应不显著,(1, 34) = 0.007,= 0.933 (见表3)。
3.1.3 情绪偏好
为了研究两组被试对不同情绪目标上偏好程度的差异, 本研究以组别(抑郁倾向组, 健康控制组)为被试间变量, 以情绪(快乐, 悲伤)为被试内变量进行重复测量方差分析。结果表明, 组别与情绪交互效应显著,(1, 34) = 12.05,0.001, 偏η= 0.26。进一步的简单效应分析发现, 对于快乐情绪的偏好, 抑郁倾向组(4.78 ± 1.06)显著小于健康控制组(5.61 ± 0.85),(34) = 2.60,0.014。对于悲伤情绪的偏好, 抑郁倾向组(2.67 ± 1.03)却显著大于健康控制组(2.00 ± 0.91),(35) = 2.89,0.007。在抑郁倾向组中, 被试对快乐情绪的偏好(4.78 ± 1.06)显著高于悲伤情绪(2.72 ± 1.02),(17) = 5.04,0.001。在健康控制组中, 被试对快乐情绪的偏好(5.61 ± 0.85)显著高于悲伤情绪(1.89 ± 0.68),(17) = 14.70,0.001。情绪主效应显著,(1, 34) = 144.78,0.001, η= 0.81。被试对快乐情绪的偏好(5.25 ± 0.15)显著大于悲伤情绪(2.33 ± 0.16)。组别主效应不显著,(1, 34) < 0.001,1.00。
最后相关分析结果表明, 被试对悲伤情绪的偏好与他们选择的悲伤图片的数量呈正相关(34) = 0.47,= 0.004, 并且被试对快乐情绪的偏好与他们选择的快乐图片的数量呈正相关(34) = 0.48,= 0.003 (见表4)。

表4 主要变量的描述统计及变量间Pearson相关
注:表格呈现了情绪偏好和被试选择的图片数量之间的Pearson相关。*< 0.05, **< 0.01。
3.2 ERP结果
3.2.1 P2与N2
对于P2的峰值, 被试内变量满足球形假设,(2,= 36) = 1.00,= 0.605。组别与图片类型的交互效应不显著,(2, 68) = 2.13,= 0.126。组别主效应也不显著,(1, 34) = 0.46,= 0.502。但图片类型的主效应显著,(2, 68) = 5.97,= 0.004, 偏η= 0.15。事后检验发现, 悲伤图片(7.60 ± 3.69 μV)和 快乐图片诱发的P2波幅(7.47 ± 3.60 μV)都显著高于中性图片(6.78 ± 3.26 μV) (悲伤:(35) = 2.63,0.013; 快乐:(35) = 3.44,0.002), 但是悲伤图片和快乐图片之间没有显著差异,(35) = 2.63,0.641 (见图3 A/B)。
对于N2的峰值, 被试内变量满足球形假设,(2,= 36) = 2.21,= 0.332。组别与图片类型的交互效应不显著,(2, 68) = 0.58,= 0.564; 组别主效应也不显著,(1, 34) = 0.75,= 0.393。但图片类型的主效应显著,(2, 68) = 17.11,< 0.001, 偏η= 0.335。事后分析显示, 悲伤图片诱发的N2波幅(−10.06 ± 5.39 μV)显著大于中性图片(−9.08 ± 4.64 μV)和快乐图片(−7.69 ± 4.50 μV) (中性:(35) = 3.21,0.003; 快乐:(35) = 5.56,0.001); 中性图片诱发的N2波幅显著大于快乐图片诱发的N2波幅,(35) = 2.81,0.008 (见图3 C/D)。
3.2.2 LPP
对于LPP的平均波幅, 被试内变量满足球形假设,(2,= 36)= 3.23,= 0.199。组别与图片类型的交互效应显著,(2, 68) = 3.29,= 0.043, 偏η= 0.09 (见图4)。进一步在图片类型的每个水平上的简单效应分析表明, 只有悲伤图片诱发的LPP波幅在抑郁倾向组(3.89 ± 2.91 μV)显著小于健康控制组(6.94 ± 2.82 μV),(34) = 3.19,0.003; 对于中性图片所诱发的LPP波幅, 抑郁倾向组(1.22 ± 2.45 μV)与健康控制组(2.48 ± 2.43 μV)没有显著差异,(34) = 2.83,0.133; 对于快乐图片所诱发的LPP波幅, 抑郁倾向组(3.65 ± 2.06 μV)与健康控制组(5.01 ± 2.81 μV)没有显著差异,(34) = 1.65,0.109 (见图5)。此外, 对于健康控制组图片诱发的LPP波幅, 悲伤图片(6.94 ± 2.82 μV)显著大于快乐图片(5.01 ± 2.81 μV)和中性图片(2.48 ± 2.43 μV) (快乐:(17) = 3.62,= 0.002; 中性:(17) = 6.93,< 0.001), 且快乐图片诱发的LPP波幅显著大于中性图片((17) = 4.24,= 0.001)。对于抑郁倾向组的LPP波幅, 悲伤图片(3.89 ± 2.91 μV)与快乐图片(3.65 ± 2.06 μV)之间并没有显著差异,(17) = 0.50,= 0.627; 但两种图片诱发的LPP波幅都显著大于中性图片(1.23 ± 2.45 μV) (悲伤:(17) = 5.36,0.001; 快乐:(17) = 4.51,< 0.001)。
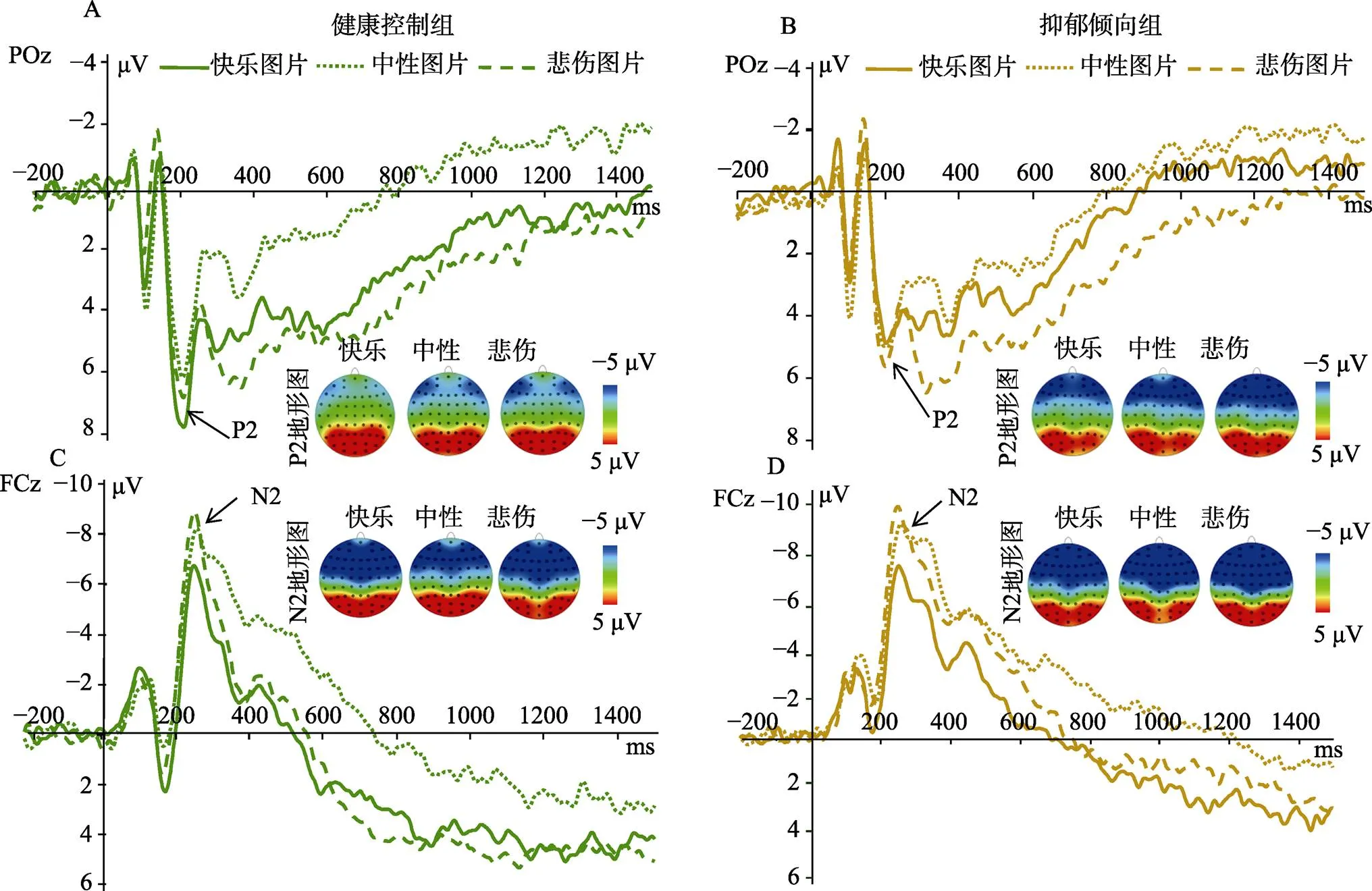
图3 A, B表示两组被试在POz电极点上对三种图片的P2的ERP总平均图, A表示健康控制组, B表示抑郁倾向组。P2成分时间窗为180~230 ms。C, D表示两组被试在FCz电极点上对三种图片的N2的ERP总平均图, C表示健康控制组, D表示抑郁倾向组。N2成分时间窗为200~300 ms。

图4 两组被试在CPz电极点上对三种图片的LPP的ERP总平均图, 其中左侧(A)表示健康控制组, 右侧(B)表示抑郁倾向组。LPP成分时间窗为600~1200 ms。

图5 两组被试在CPz电极点上对三种类型的图片的LPP波幅
方差分析表明组别主效应显著,(1, 34) = 6.51,= 0.015, 偏η= 0.16, 图片类型主效应显著,(2, 68) = 43.30,0.001, 偏η= 0.56。事后检验发现, 抑郁倾向组被图片诱发的LPP波幅(2.93 ± 0.52 μV)显著小于健康控制组(4.81 ± 0.52 μV)。悲伤图片诱发的LPP波幅(5.42 ± 3.22 μV)显著大于快乐图片诱发的LPP波幅(4.33 ± 2.53 μV)和中性图片诱发的LPP波幅(1.85 ± 2.49 μV) (快乐:(35) = 2.83,0.008 ; 中性(35) = 7.80,0.001)。且快乐图片诱发的LPP波幅显著大于中性图片诱发的LPP波幅,(35) = 6.71,< 0.001。
4 讨论
本文研究了抑郁倾向被试对快乐、中性和悲伤三种情绪场景图片的加工和选择, 首次采用事件相关电位技术研究抑郁倾向被试的情绪调节目标。研究发现, 与正常人相比, 抑郁倾向组对于悲伤情绪的偏好显著增加, 对快乐情绪的偏好则显著降低。而且在面对悲伤情景时, 抑郁倾向个体更多地选择再看悲伤图片, 悲伤图片诱发的LPP波幅显著低于正常个体。LPP波幅可以反映动机的强度(Gable & Harmon-Jones, 2013), 因此我们的结果提示抑郁倾向被试对负性刺激的回避动机显著降低, 从而比正常人更多的选择再次观看悲伤图片。
首先, 我们发现与中性图片相比, 悲伤图片和快乐图片都会诱发更大的LPP波幅。这与前人的研究结果一致, 即情绪性刺激(图片、文字)比中性刺激诱发更大的LPP波幅(Codispoti, Ferrari & Bradley, 2006; Dennis & Hajcak, 2009; Pastor et al., 2008; Proudfit et al., 2015)。而且在健康控制组中, 悲伤图片诱发的LPP波幅显著大于快乐图片, 提示正常被试对负性情绪刺激的评估和编码更为精细和复杂(Olofsson, Nordin, Sequeira & Polich, 2008)。研究表明负性情绪刺激可以诱发被试更高的唤醒度, 且唤醒度越高, 图片诱发的LPP波幅越大(Cuthbert, Schupp, Bradley, Birbaumer & Lang., 2000; Schupp et al., 2000), 因此被试可能会投入更多的注意资源, 从而实现情绪刺激的精细编码。从生物进化的角度来看, 负性事件和刺激对人的生存和进化更具有威胁性, 可以诱发更强的情绪和动机反应, 从而触发更深层次和更精细的加工。相关的研究结果也表明令人厌恶的图片或文字刺激确实会比积极的刺激诱发更强的电生理反应, 产生强烈的情感和动机反应(Baumeister, Bratslavsky, Finkenauer & Vohs, 2001; Diamond, 2001)。而在我们的研究中, 抑郁倾向被试在对图片材料进行加工时, 悲伤图片与快乐图片诱发的LPP波幅并没有显著差异, 这就提示对于抑郁倾向被试对负性情绪刺激的精细加工能力减弱, 或者负性信息可能不能够诱发抑郁倾向个体更强的情绪和动机反应。
其次, 我们还发现抑郁倾向个体加工悲伤图片所产生的LPP幅度显著小于正常人。这个结果也与以往研究的结果一致, 即抑郁症病人以及抑郁程度较高的个体对情绪性刺激(图片、文字)都表现出更弱的LPP波幅(Blackwood, 1990; Macnamara et al., 2016; Weinberg, Perlman, Kotov & Hajcak, 2016)。而且情绪刺激所诱发的LPP波幅与抑郁症病人的抑郁程度呈负相关(Macnamara et al., 2016; Weinberg et al., 2016)。研究表明LPP能反映动机的强度却不能反映动机的方向, 但情绪效价却与动机的方向有关, 正性情绪与趋近动机有关, 而负性情绪与回避动机有关(Bamford & Ward, 2008; Neumann & Strack, 2000)。
已有研究表明回避动机越强的负性刺激, 诱发的LPP波幅也更大(Leutgeb et al., 2009; Michalowski et al., 2009)。例如, 威胁性刺激和残肢断臂的图片会比损失和污染的场景诱发更大的LPP波幅(Schupp, Cuthbert et al., 2004; Schupp, Öhman, et al., 2004; Schupp, Junghöfer, Weike & Hamm, 2004)。相反地, 对负性刺激的回避动机越强的个体, 刺激所诱发的LPP波幅也更大。例如, 蜘蛛恐惧症个体观看蜘蛛图片比观看其他的负性图片产生更大的LPP波幅, 且蜘蛛恐惧症个体比正常人在观看蜘蛛图片时产生了更强的LPP波幅(Leutgeb et al., 2009; Michalowski et al., 2009)。另外, 焦虑症病人观看愤怒面孔和恐惧面孔也会有更大的LPP波幅(MacNamara et al., 2016)。而我们的研究发现与健康控制组相比, 抑郁倾向组悲伤图片诱发的LPP波幅显著降低, 说明抑郁倾向被试对于负性刺激的回避动机减弱, 从而使得在行为学研究中抑郁倾向个体选择的悲伤图片比正常个体多。Millgram等(2015)在正常人和抑郁症病人的对照研究中也发现, 抑郁症病人更倾向于选择继续观看悲伤图片。这些结果都提示抑郁倾向个体对负性信息的回避动机降低, 从而导致在生活中他们无法及时回避负性刺激和事件。
此外前人的研究发现对正性刺激的趋近动机越强, 刺激诱发的LPP波幅越大(Gable & Harmon-Jones, 2013; Gable & Poole, 2014; Hajcak et al., 2010)。例如, 与动机性较小的正性刺激(比如热烈的运动场景)相比, 色情图片能够诱发更强的LPP波幅(Briggs & Martin, 2009b; Hajcak et al., 2010)。动机强度较高的欲求刺激(如甜品)能够诱发更大的LPP波幅(Gable & Harmon-Jones, 2013), 且只有较高动机强度的正性刺激能够诱发更大的LPP, 中性刺激没有这种效应(Gable & Harmon-Jones, 2013; Gable & Poole, 2014)。在我们的研究中, 我们并没有发现被试组间PANAS评分存在显著差异, 可以排除被试当时情绪状态对图片选择任务以及LPP波幅的影响。此外两组被试对图片的评定没有差异, 且两组被试对快乐图片的选择没有显著差异, 并且由快乐图片所诱发的LPP波幅也没有显著差异, 但却存在这样的趋势, 即与正常被试相比, 抑郁倾向被试对图片快乐评分更低, 悲伤评分更高, 加工快乐图片所产生的LPP波幅较弱, 选择继续观看的快乐图片也较少。这可能是因为我们选取的只是具有抑郁倾向的被试, 并没有临床上的抑郁症状, 这些结果在某种程度上已经提示抑郁倾向被试对快乐刺激的趋近动机也比正常被试弱, 从而比正常个体较少地选择再次观看快乐图片。
对于与情绪加工过程有关的早期成分P2的研究发现, 与中性图片相比, 快乐图片和悲伤图片诱发的P2波幅更大, 但这两者之间没有显著的差别, 这与之前的研究的结果一致(Carretié, Hinojosa, Martín-Loeches, Mercado & Tapia, 2004; Carretié, Mercado, Hinojosa, Martín-Loeches & Sotillo, 2004)。P2成分被认为反映了情绪图片加工早期阶段快速而粗略的注意资源分配, 情绪性刺激会得到更多注意资源的分配, 因此得到优先的加工, 呈现出更强的波幅。对于中期成分N2的研究, 我们发现悲伤图片诱发的N2波幅大于中性图片和快乐图片诱发的N2波幅。这一结果支持情绪加工的“负性偏向”理论(Krendl, Zucker & Kensinger, 2017; Olofsson et al., 2008), 这种负性偏向被认为是个体对负性刺激快速注意资源分配的结果。这种对负性刺激的快速反应对个体和种群来说具有适应意义, 可以帮助人们更快地探测和躲避具有威胁性的情境(Olofsson et al., 2008)。因此与正性和中性的刺激相比, 注意会自动地朝向威胁性刺激(Öhman & Mineka, 2001)并对其优先加工。此外, 我们的研究显示中性图片诱发的N2波幅大于快乐图片, 这一结果可能与N2对负性刺激的抑制功能有关(Nieuwenhuis, Yeung, van den Wildenberg & Ridderinkhof, 2003)。研究表明N2成分可以表征对负性刺激的主动抑制, N2波幅越强表明被试的主动抑制能力越强(Lewis et al., 2008)。我们要求被试观看图片时决定自己是否喜欢再看刚呈现的图片, 因此被试在观看图片时可能会自动地对负性情绪进行抑制, 当其对信息进行加工的时候, 负性刺激应该会比中性刺激诱发更大的波幅; 而快乐图片是最不需要抑制的, 因此快乐图片会比中性图片诱发更小的N2波幅。
综上所述, 对于快乐刺激, 越大的LPP波幅表示越强的趋近动机; 对于悲伤刺激, 越大的LPP表示越强的回避动机。而本研究结果发现抑郁倾向个体对快乐图片和悲伤图片的LPP均小于正常个体。这就表明抑郁倾向个体对快乐刺激的趋近动机和对悲伤刺激的回避动机都弱于正常个体, 从而使得他们比正常个体更少选择能够使他们感到快乐的场景, 且更少回避那些使他们感到悲伤的场景。因此抑郁症病人或抑郁倾向个体比正常个体倾向于更多地体验到悲伤, 且更少地体验到快乐。我们的研究将有助于深入了解抑郁症情绪失调的神经机制, 同时找到更多的情绪相关的生物标记物以帮助对抑郁倾向个体及重度抑郁个体进行干预和治疗。下一步还可以从以下几个方面对抑郁症个体的情绪调节目标及其潜在的神经机制进行研究:1)选用多种情绪类型的场景图片(比如, 恐惧, 厌恶等), 考察抑郁症个体与正常个体之间情绪调节目标的差异是否对其他具体情绪也会有同样的结果; 2)考察抑郁症个体对社会性情绪载体(比如, 情绪性面孔)进行加工时的行为与神经活动; 3)辨别抑郁症个体的情绪调节目标对他们的情绪失调的影响是短暂的还是长期的。
Admon, R., & Pizzagalli, D. A. (2015). Dysfunctional reward processing in depression.,114–118https://doi.org/10.1016/j.copsyc.2014.12.011
Aldao, A., Nolen-hoeksema, S., & Schweizer, S. (2010). Emotion- regulation strategies across psychopathology : A meta-analytic review.,(2)217–237.https://doi.org/10.1016/j.cpr.2009.11.004
Bai, L., Ma, H., Huang, Y.-X., & Luo, Y. J. (2005). The development of native chinese affective picture system--A pretest in 46 college students.,(11), 719–722.
[白露, 马慧, 黄宇霞, 罗跃嘉.(2005). 中国情绪图片系统的编制——在46名中国大学生中的试用.,(11), 719–722.]
Bamford, S., & Ward, R. (2008). Predispositions to approach and avoid are contextually sensitive and goal dependent.,(2)174–183. https://doi.org/ 10.1037/1528-3542.8.2.174
Baumeister, R. F., Bratslavsky, E., Finkenauer, C., & Vohs, K. D. (2001). Bad is stronger than good.,(4)323–370. https://doi.org/10.1037//1089- 2680.5.4.323
Briggs, K. E., & Martin, F. H. (2009a). Affective picture processing and motivational relevance: Arousal and valence effects on ERPs in an oddball task.,(3)299–306. https://doi.org/10.1016/ j.ijpsycho.2009.01.009
Briggs, K. E., & Martin, F. H. (2009b). Affective picture processing and motivational relevance: Arousal and valence effects on ERPs in an oddball task.,(3), 299–306. https://doi.org/ 10.1016/j.ijpsycho.2009.01.009
Carretié, L., Hinojosa, J. A., Martín-Loeches, M., Mercado, F., & Tapia, M. (2004). Automatic attention to emotional stimuli: Neural correlates.,(4), 290–299. https://doi.org/10.1002/hbm.20037
Carretié, L., Mercado, F., Hinojosa, J. A., Martín-Loeches, M., & Sotillo, M. (2004). Valence-related vigilance biases in anxiety studied through event-related potentials.,(2), 119–130. https://doi.org/10.1016/ S0165-0327(02)00242-2
Codispoti, M., Ferrari, V., & Bradley, M. M. (2006). Repetitive picture processing: Autonomic and cortical correlates.,(1), 213–220. https://doi.org/10.1016/ j.brainres.2005.11.009
Cuthbert, B. N., Schupp, H. T., Bradley, M. M., Birbaumer, N., & Lang, P. J. (2000). Brain potentials in affective picture processing: Covariation with autonomic arousal and affectivereport.(2), 95–111. https://doi.org/ 10.1016/S0301-0511(99)00044-7
Dennis, T. A., & Hajcak, G. (2009). The late positive potential: A neurophysiological marker for emotion regulation in children.,(11), 1373–1383. https://doi.org/10.1111/ j.1469-7610.2009.02168.x
Ehring, T., Tuschen-Caffier, B., Schnülle, J., Fischer, S., & Gross, J. J. (2010). Emotion regulation and vulnerability to depression: Spontaneous versus instructed use of emotion suppression and reappraisal.,(4), 563–572. https://doi.org/10.1037/a0019010
Elliot, A. J., & Thrash, T. M. (2002). Approach-avoidance motivation in personality: Approach and avoidance temperaments and goals.,(5), 804–818. https://doi.org/10.1037//0022- 3514.82.5.804
Feng, C., Li, W., Tian, T., Luo, Y., Gu, R., Zhou, C., & Luo, Y.-J. (2014). Arousal modulates valence effects on both early and late stages of affective picture processing in a passive viewing task.,(4), 364–377. https://doi.org/10.1080/17470919.2014.896827
Gable, P. A., & Harmon-Jones, E. (2013). Does arousal per se account for the influence of appetitive stimuli on attentional scope and the late positive potential?,(4), 344–350. https://doi.org/10.1111/psyp.12023
Gable, P. A., & Poole, B. D. (2014). Influence of trait behavioral inhibition and behavioral approach motivation systems on the LPP and frontal asymmetry to anger pictures.,(2), 182–190. https://doi.org/10.1093/scan/nss130
Garnefski, N., & Kraaij, V. (2006). Relationships between cognitive emotion regulation strategies and depressive symptoms: A comparative study of five specific samples.,(8), 1659–1669. https://doi.org/10.1016/j.paid.2005.12.009
Gross, J. J. (1998). The emerging field of emotion regulation: An integrative review.,(3), 271–299. https://doi.org/10.1037/1089-2680.2.3.271
Gross, J. J. (2015). The extended process model of mmotion regulation: Elaborations, applications, and future directions.,(1), 130–137. https://doi.org/ 10.1080/1047840X.2015.989751
Hajcak, G., MacNamara, A., & Olvet, D. M. (2010). Event- related potentials, emotion, and emotion regulation: An integrative review.,(2), 129–155. https://doi.org/10.1080/87565640903526504
Harmon-Jones, E., Harmon-Jones, C., Amodio, D. M., & Gable, P. A. (2011). Attitudes toward emotions.,(6), 1332–1350. https://doi.org/ 10.1037/a0024951
Ito, T. A., Larsen, J. T., Smith, N. K., & Cacioppo, J. T. (1998). Negative information weighs more heavily on the brain: The negativity bias in evaluative categorizations.,(4), 887–900. https://doi.org/10.1037/0022-3514.75.4.887
Joormann, J., & Gotlib, I. H. (2010). Emotion regulation in depression: Relation to cognitive inhibition.,(2), 281–298. https://doi.org/10.1080/02699930903407948
Krendl, A. C., Zucker, H. R., & Kensinger, E. A. (2017). Examining the effects of emotion regulation on the ERP response to highly negative social stigmas.,(3), 349–360.
Lang, P. J., Bradley, M. M., & Cuthbert, B. N. (1997). International Affective Picture System (IAPS): Technical Manual and Affective Ratings. NIMH Center for the Study of Emotion and Attention, 39–58. https://doi.org/10.1027/ 0269-8803/a000147
Lang, P. J., & Bradley, M. M. (2010). Emotion and the motivational brain.,(3), 437–450. https://doi.org/10.1016/j.biopsycho.2009.10.007
Leutgeb, V., Schäfer, A., & Schienle, A. (2009). An event- related potential study on exposure therapy for patients suffering from spider phobia.,(3), 293–300. https://doi.org/10.1016/j.biopsycho.2009.09.003
Lewis, M. D., Granic, I., Lamm, C., Zelazo, P. D., Stieben, J., Todd, R. M., … Pepler, D. (2008). Changes in the neural bases of emotion regulation associated with clinical improvement in children with behavior problems.,(3), 913–939. https://doi.org/10.1017/ S0954579408000448
MacNamara, A., Kotov, R., & Hajcak, G. (2016). Diagnostic and symptom-based predictors of emotional processing in generalized anxiety disorder and major depressive disorder: An event-related potential study.,(3), 275–289. https://doi.org/10.1007/s10608- 015-9717-1
Michalowski, J. M., Melzig, C. A., Weike, A. I., Stockburger, J., Schupp, H. T., & Hamm, A. O. (2009). Brain dynamics in spider-phobic individuals exposed to phobia-relevant and other emotional stimuli.,(3), 306–315. https://doi.org/10.1037/a0015550
Millgram, Y., Joormann, J., Huppert, J. D., & Tamir, M. (2015). Sad as a matter of choice? Emotion-regulation goals in depression.,(8), 1216–28. https:// doi.org/10.1177/0956797615583295
Neumann, R., & Strack, F. (2000). Approach and avoidance: The influence of proprioceptive and exteroceptive cues on encoding of affective information.,(1), 39–48. https://doi.org/10.1037// 0022-3514.79.1.39
Nieuwenhuis, S., Yeung, N., van den Wildenberg, W., & Ridderinkhof, K. R. (2003). Electrophysiological correlates of anterior cingulate function in a go/no-go task: Effects of response conflict and trial type frequency.,(1), 17–26. https:// doi.org/10.3758/CABN.3.1.17
Öhman, A., & Mineka, S. (2001). Fears, phobias, and preparedness: Toward an evolved module of fear and fear learning.,(3), 483–522. https:// doi.org/10.1037//0033-295X.108.3.483
Olofsson, J. K., Nordin, S., Sequeira, H., & Polich, J. (2008). Affective picture processing: An integrative review of ERP findings.,(3), 247–265. https:// doi.org/10.1016/j.biopsycho.2007.11.006
Pastor, M. C., Bradley, M. M., Löw, A., Versace, F., Moltó, J., & Lang, P. J. (2008). Affective picture perception: Emotion, context, and the late positive potential.,(1), 145–151. https://doi.org/10.1016/ j.brainres.2007.10.072
Proudfit, G. H., Bress, J. N., Foti, D., Kujawa, A., & Klein, D. N. (2015). Depression and event-related potentials: Emotional disengagement and reward insensitivity.,, 110–113. https://doi.org/10.1016/ j.copsyc.2014.12.018
Schupp, H., Cuthbert, B., Bradley, M., Hillman, C., Hamm, A., & Lang, P. (2004). Brain processes in emotional perception: Motivated attention.,(5), 593–611.
Schupp, H. T., Cuthbert, B. N., Bradley, M. M., Cacioppo, J. T., Ito, T., & Lang, P. J. (2000). Affective picture processing: The late positive potential is modulated by motivational relevance.,(2), 257–261. https://doi.org/10.1111/1469-8986.3720257
Schupp, H. T., Junghöfer, M., Weike, A. I., & Hamm, A. O. (2004). The selective processing of briefly presented affective pictures: An ERP analysis.,(3), 441–449. https://doi.org/10.1111/j.1469-8986.2004. 00174.x
Schupp, H. T., Öhman, A., Junghöfer, M., Weike, A. I., Stockburger, J., & Hamm, A. O. (2004). The facilitated processing of Threatening Faces: An ERP analysis.,(2), 189–200. https://doi.org/10.1037/1528-3542.4.2.189
Tamir, M. (2009). Differential preferences for happiness: Extraversion and trait-consistent emotion regulation.,(2), 447–470. https://doi.org/ 10.1111/j.1467-6494.2008.00554.x
Tamir, M., & Ford, B. Q. (2012). When feeling bad is expected to be good: Emotion regulation and outcome expectancies in social conflicts.,(4), 807–816. https://doi.org/10.1037/a0024443
Twenge, J. M., & Campbell, W. K. (2001). Age and birth cohort differences in self-esteem: A cross-temporal meta- analysis.(4), 321–344.
Weinberg, A., & Hajcak, G. (2010). Beyond good and evil: The time-course of neural activity elicited by specific picture content.,(6), 767–782. https://doi.org/ 10.1037/a0020242
Weinberg, A., Perlman, G., Kotov, R., & Hajcak, G. (2016). Depression and reduced neural response to emotional images: Distinction from anxiety, and importance of symptom dimensions and age of onset.(1), 26–39. https://doi.org/ 10.1037/abn0000118
Yang, W., Zhu, X., Wang, X., Wu, D., & Yao, S. (2011). Time course of affective processing bias in major depression: An ERP study.,(3), 372–377. https://doi.org/10.1016/j.neulet.2010.10.059
Emotional regulation goals of young adults with depression inclination: An event-related potential study
LI Hong; YANG Xiaoguang; ZHENG Wenyu; WANG Chao
(School of Psychology, Shenzhen University, Shenzhen 518060, China) (Shenzhen Key Laboratory of Affective and Social Cognitive Science, Shenzhen University, Shenzhen 518060, China)(Shenzhen Institute of Neuroscience, Shenzhen 518060, China)
Researches on deficits in emotion regulation of depression have mainly focused on the selection and application of emotion-regulation strategies; however, it remains unclear whether it is also related to emotion-regulation goals, i.e., the direction of emotion regulation. Situation selection is an antecedent-focused regulation strategy that is worked before the emotional reactions occur and it can be used as an index of emotional-regulation goals.
In our current study, the event-related potential (ERP) technique was used to investigate the emotion-regulation goals of young adults with depression inclination. Participants were asked to freely select the emotion-inducing scenes in which they want to put themselves and to report their emotional preferences。
ERP results revealed that the amplitudes of Late Positive Potential (LPP) were significantly decreased when viewing the sadness scene in young adults with depression inclination, and they selected to view sadness scene more frequently than healthy young adults. In addition, the ratings of sadness preferences were significantly higher among subjects with depression inclination, while the happiness preferences were lower.
The current results suggest that, compared to the control group, the individuals with depression inclination are more willing to use situation selection to maintain or enhance their sadness rather than weaken it or enhance their happiness. These findings further indicate that emotion regulation goals of depressive subjects may be related to their motivations for selecting emotional stimulus, and provide a new perspective for exploring the causes and mechanisms of emotion regulation deficits in depressive disorders.
emotion regulation; emotion regulation goal; depression inclination; approach-avoidance motivation; late positive potential
2018-03-14
* 广东省普通高校创新团队建设项目(2015KCXTD009), 广东省省级(基础研究与应用研究)重大项目(2016KZDXM009), 国家自然科学基金项目(31600920), 深圳市基础研究布局项目(JCYJ20150729104249783), 深圳市孔雀计划项目(KQTD2015033016104926), 深圳市自由探索项目(JCYJ20170818102123707)资助。
王超, E-mail: chao.wang@szu.edu.cn
10.3724/SP.J.1041.2019.00637
B845; B842
