青钱柳黄酮及三萜调节人体肠道菌群作用研究
2019-06-11陈玉慧郭彤彤郑晓杰
陈玉慧,郭彤彤,张 鑫*,郑晓杰
(1.宁波大学 海洋学院,浙江 宁波315211;2.温州科技职业学院 农业与生物技术学院,浙江 温州325000)
青钱柳[Cyclocaryapaliurus(Batal.)Iljinsk]是我国特有的胡桃科、青钱柳属植物。青钱柳富含黄酮、三萜类物质,具有清热解暑、降血糖、降血脂、抗疲劳等多种功效,在我国作茶已有上千年历史[1-3]。黄酮类化合物是植物次生代谢产物,具有各种结构及生理活性。青钱柳中的黄酮类物质主要包括山奈酚、槲皮素和异槲皮素[4]。而青钱柳中的三萜类物质包括青钱柳独有的青钱柳酸A、青钱柳酸B、青钱柳甙I、青钱柳甙II、青钱柳甙III及阿江橄榄酸、乌江酸、齐墩果酸、乳香酸等[5]。青钱柳中的天然黄酮及三萜类化合物,因其多种药理活性而成为国内外的研究热点。
在人体结肠中,微生物发挥着影响宿主营养物质获取和能量调节的关键作用[6]。肠道微生物可分为有益细菌和有害细菌[7],其中有益菌占总数的90%以上,主体是双歧杆菌、乳酸杆菌等[8]。益生元是不易消化的食物成分,其可以通过刺激肠道中特定菌群的生长,有效改善宿主肠道微生态[9]。虽然植物多酚在体外发挥了较好的生物活性,然而与体内实验还缺乏一致性。在进入人体肠道后,植物多酚被肠道菌群代谢成不同结构和功能的衍生物,这些代谢衍生物可能是其在体内真正发挥生物活性的有效成分。据报道,不同菌株的肠道细菌的增殖对于各种植物多酚及其代谢产物具有不同的敏感性[10]。在我们之前的研究中植物多酚展现了对肠道有益细菌的增殖作用[8,11]。然而关于青钱柳中多酚类物质对于肠道菌群的影响,还鲜有报道。
本研究拟在实验室之前的工作基础上,利用大孔树脂分离纯化青钱柳黄酮及三萜,并利用体外厌氧粪样混合培养方式对于样品进行发酵培养,通过荧光原位杂交技术(FISH)分析厌氧发酵过程中双歧杆菌、乳酸菌、拟杆菌、梭状菌以及总菌群的数量变化,阐明其青钱柳黄酮及三萜对于人体肠道微生态的调节作用,为青钱柳资源的综合利用提供理论依据。
1 材料与方法
1.1 材料与试剂
青钱柳由浙江省文成县泉山中药材种植有限公司提供,去除杂质和枝杆之后,粉碎过60目筛备用。聚酰胺及AB-8树脂购于青岛海洋化工有限公司;低聚果糖购于美国Sigma公司;厌氧混合气体(10% H2、10% CO2和80% N2)和高纯N2购于宁波甬兴化工气体有限公司;4,6-联眯-2-苯基吲哚二盐酸(DAPI)荧光染料购于德国Roche 公司;表1所示的FISH专用16S rRNA寡合苷酸单链探针由上海生工生物工程技术服务有限公司合成[8]。

表1 16S rRNA探针的靶细菌种/属和探针序列
1.2 仪器与设备
Laborota 4000 真空旋转蒸发仪,德国Heidolph公司;SHB-III型循环水式真空泵,郑州长城科工贸有限公司;冷冻干燥机,宁波新芝公司;AY-120 电子精密天平、BL-220H 分析天平,日本Shimadzu 公司;YQX-I厌氧培养箱,上海跃进医疗器械厂;Zeiss Axio ImagerA1 型荧光正置显微镜系统,德国Carl Zeiss 公司)。
1.3 实验方法
1.3.1 青钱柳黄酮样品的制备
称取适量的粉碎青钱柳粉末,按照料液比1∶20加入热水,在60 ℃恒温振荡器中振荡提取40 min,4 500 ×g的条件下离心15 min得上清液,残渣用同样方法再次浸提,合并两次上清液并浓缩、冻干,得到青钱柳黄酮粗提物。
将青钱柳黄酮粗提物制成2%质量分数的溶液,经0.45 μm滤膜过滤后上聚酰胺层析柱(30 cm × 1.6 cm)。首先用水洗去水溶性杂质,洗脱流速为2.0 mL/min,洗脱2 BV(BV为柱床体积)后,用80%的乙醇洗脱,采用自动部分收集器收集洗脱液(10 mL/管),浓缩、冻干,得到青钱柳黄酮样品。
1.3.2 青钱柳黄酮含量的测定
标准曲线的绘制:精密称取芦丁标准品用80%乙醇定容,配成浓度0.4 mg/mL的芦丁标准品。精密吸取标准品溶液0.2 mL、0.4 mL、0.6 mL、1.0 mL、1.2 mL分别置于10 mL试管中,分别加入50 mg/mL亚硝酸钠溶液0.3 mL,摇匀后放置6 min后;加入10 mg/mL硝酸铝溶液0.3 mL,摇匀后用放置6 min后;加入40 mg/mL氢氧化钠溶液4 mL,加水至刻度,混匀,放置15 min。用紫外分光光度计在509 nm波长范围测定吸光度,测定3次取平均值,以吸光度为横坐标,芦丁标准品浓度值为纵坐标,绘制芦丁标准品溶液浓度与吸光度的标准曲线[12]。称取青钱柳黄酮样品粉末溶于40%乙醇,按标准曲线中所述测定方法测定吸光度,根据回归方程计算样品中的总黄酮含量。
1.3.3 青钱柳三萜样品的制备
精确称取干燥的青钱柳叶粉末0.5 g,用滤纸包好后至于索氏提取装置中用石油醚脱脂脱色3 h(等体积脱脂2次),挥干石油醚,以60%乙醇为提取剂提取,其中料液比为1:40,提取温度为60 ℃,提取时间为45 min,4 500 ×g的条件下离心10 min得上清液,同条件浸提2次,合并提取液,真空浓缩得到青钱柳三萜粗提物[5]。
将青钱柳三萜粗提物制成2%质量分数的溶液,经0.45 μm滤膜过滤后上AB-8层析柱(30 cm × 1.6 cm)。首先用水洗去水溶性杂质,洗脱流速为2.0 mL/min,洗脱 2 BV(BV为柱床体积)后,用60%的乙醇洗脱,采用自动部分收集器收集洗脱液(10 mL/管),浓缩、冻干,得到青钱柳三萜样品。
1.3.4 青钱柳三萜含量的测定
标准曲线的绘制:精密称取熊果酸标准品用甲醇定容,配成0.09 mg/mL的溶液,精确吸取0.4 mL、0.6 mL、0.8 mL、1.0 mL、1.2 mL、1.4 mL,分别加入具塞试管,水浴挥干溶剂。分别加入5% 香草醛-冰乙酸和高氯酸0.3 mL和1.0 mL,摇匀,70 ℃水浴15 min,然后迅速冷却到常温后加入4.0 mL冰乙酸,与550 nm波长处测定吸光度,以不加对照用品溶液而直接加入5% 香草醛-冰乙酸、高氯酸和冰乙酸的混合液作为空白对照,测定3次取平均值,以吸光度为横坐标,熊果酸标准品浓度值为纵坐标,绘制熊果酸标准品溶液浓度与吸光度的标准曲线。称取青钱柳三萜样品粉末溶于甲醇,按标准曲线中所述测定方法测定吸光度,根据回归方程计算样品中的三萜含量。
1.3.5 体外厌氧粪样混合培养
粪便样本的收集与处理:收集受试者(受试者为5人,均为25~30岁之间健康人群,且在近两个月内未服用过抗生素)的一次新鲜全便,置于一次性硬质无菌塑料袋中。使用时混匀并称取0.5 g,加入4.5 mL经过滤、脱氧处理的PBS缓冲液,迅速置于厌氧培养箱中,经旋涡混合器振荡混匀3 min,待用。
体外厌氧粪样混合培养按照本实验室建立的方法进行[8]。首先称取受试的儿茶素样品溶解于高压灭菌后的含氮基础培养基中,得到终质量浓度为2 g/100 mL的底物溶液。然后取粪便样液(10%、pH 7.3的PBS )150 μL悬浮于1.35 mL含有各种儿茶素样品的底物培养基溶液中,混合均匀后放入厌氧培养箱(10% H2、10% CO2、80% N2),37 ℃ 发酵培养。在培养时间为 0、6、12、24 h时分别取 100 μL 样液,供细胞FISH计数。实验设3个重复,实验对照为不加任何青钱柳黄酮或三萜的样品。
1.3.6 菌体计数
菌体计数采用FISH法[8]。将 300 μL 的多聚甲醛(4 g/100 mL、pH 7.2)加入到 100 μL体外厌氧粪样混合培养发酵样液中,置于4 ℃冰箱中冷藏 16 h,离心后用PBS缓冲液冲洗两次,再用300 μL PBS-酒精(1:1)保存菌体。取菌体样液10 μL,滴入经过铬明胶溶液包埋过的载玻片圆孔内,平铺于整个圆圈,在阴暗处自然风干3 h,然后用50%、80%、96%的乙醇溶液进行连续脱水操作各3 min,自然风干。将1 μL 50 ng/μL 的探针试剂与9 μL杂交缓冲液加到载玻片的菌体试样中,然后迅速将载玻片置入含有杂交缓冲液的湿盒中,根据探针类型采用不同温度条件(探针Bif164:50 ℃,探针Lab158:45 ℃,探针Bac303:45 ℃,探针Chis150:50 ℃)进行杂交10 h。杂交结束后,迅速用清洗缓冲液与超纯水冲洗载玻片,取出试样载玻片,在避光条件下自然风干。测定总菌体数时,将10 μL 1.25 ng/μL的DAPI溶液加入到杂交后的载玻片上,保持5 min,然后用超纯水冲洗,自然干燥。所有样品用Zeiss Axio Imager A1型荧光正置显微镜系统进行观测,采用AxioVision荧光成像软件对每个试样随机选择10-15个视野进行拍照,记录荧光光点。
1.3.7 数据分析
在对杂交后的各种菌体进行拍照与荧光光点计数后,根据试样浓度以及稀释倍数等计算出细菌总数,并取其以10为底的对数。实验数据采用SPSS软件进行显著性分析。
2 结果与分析
2.1 青钱柳黄酮及三萜样品的制备
青钱柳经热水浸提,浓缩和冻干,得到黄酮及三萜粗提物。由于粗提物中含有一定量茶多糖等杂质,因此根据黄酮及三萜的特性,分别选取聚酰胺及AB-8树脂,利用大孔树脂层析柱对其进行分离纯化。经过聚酰胺大孔树脂柱层析分离之后,青钱柳中黄酮的含量提高到95%。同时,经过AB-8大孔树脂柱层析分离之后,青钱柳三萜的含量提高到90%。
2.2 青钱柳黄酮及三萜对肠道中有益菌群(双歧杆菌和乳酸菌)的增殖作用
表2为以青钱柳黄酮和三萜为底物时,不同发酵时间有益菌群(双歧杆菌和乳酸菌)的增殖情况。由表2可以看出,随着发酵时间的延长,各组中双歧杆菌的数量逐渐增多。在每个发酵时间点,阳性对照低聚果糖都具有最好的促进双歧杆菌增殖的效果,同时在不同发酵时间点,青钱柳黄酮和三萜样品与对照组相比,都可以显著促进双歧杆菌的增殖(P<0.05)。当培养时间为6 h及12 h时,青钱柳黄酮和三萜同样促进了双歧杆菌的增殖,但差异不显著(P> 0.05)。而厌氧培养24 h时,青钱柳黄酮相对于三萜效果更好,可能是由于其不同的结构导致,相应的构效关系还有待进一步研究。
由表2中可以看出,在发酵时间分别为0、6、12与24 h时,乳酸菌数量在添加青钱柳黄酮和三萜的实验组中显著增多,说明样品能积极促进乳酸菌增殖。与对照组相比,在不同发酵时间点,青钱柳黄酮和三萜对乳酸菌的增殖作用也具有显著差异(P< 0.05),说明其对乳酸菌也有较好的增殖效果。与对双歧杆菌的影响类似,在厌氧培养24 h时,青钱柳黄酮相对于三萜发挥了更好的促进乳酸菌增殖的效果。
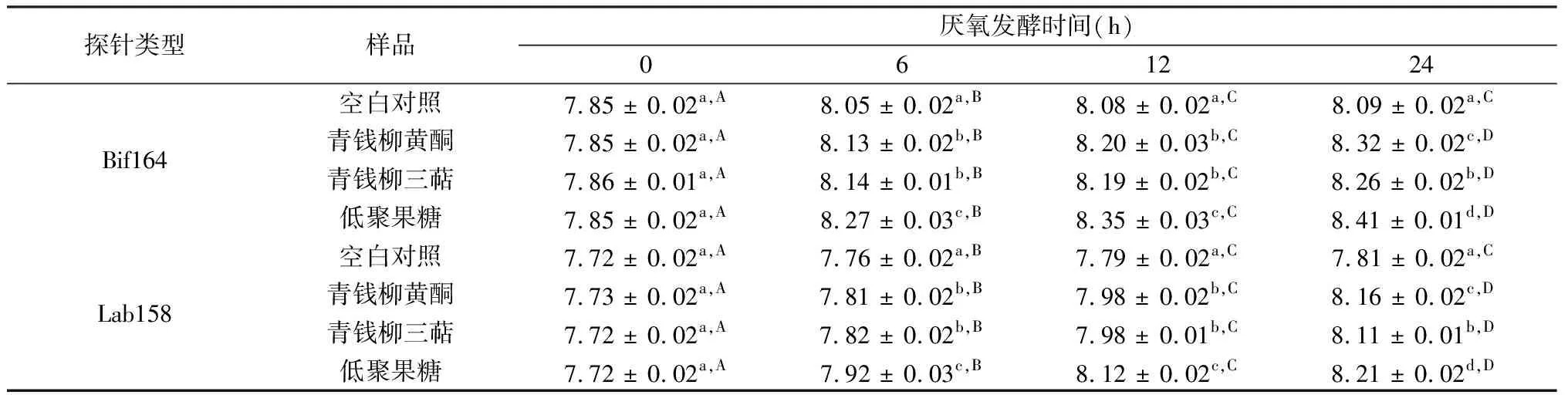
表2 青钱柳黄酮和三萜体外厌氧发酵0、6、12、24 h后双歧杆菌和乳酸菌的数量 lg(CFU/mL)
注:同列不同小写字母代表同一时间不同样品之间对于该种菌属有显著性差异(P<0.05);同行不同大写字母代表同一样品对于该种菌属在不同发酵时间有显著性差异(P<0.05)。下同。
2.3 青钱柳黄酮及三萜对肠道中拟杆菌和梭状菌的增殖作用
表3为以青钱柳黄酮及三萜为底物时,不同发酵时间拟杆菌和梭状菌的增殖情况。由表中可以看出,虽然每组拟杆菌的数量均随着发酵时间延长逐渐增加,然而,相对于空白对照组,各组样品均有效抑制了拟杆菌的增殖。在厌氧发酵超过6 h之后,阳性对照组的低聚果糖在每个时间点都显示了最为明显的抑制增殖的效果。同时与对照组相比,青钱柳黄酮及三萜对拟杆菌的增殖均表现出显著的抑制作用(P<0.05)。当发酵时间为6 h时,青钱柳黄酮及三萜抑制拟杆菌增殖的效果差异不显著(P<0.05),而发酵时间为12和24 h时,青钱柳黄酮相对于三萜,更为显著抑制了拟杆菌的增殖(P<0.05)。
与拟杆菌类似,青钱柳黄酮及三萜同样显著抑制了梭状菌的增殖。由表3可以看出,随着发酵时间的延长,虽然各组拟杆菌的数量不断增加,但与对照组相比,在不同的发酵时间点,青钱柳黄酮及三萜均显著抑制了梭状菌的增殖。而且值得注意的是,在发酵时间为12和24 h时,青钱柳黄酮相对于三萜,发挥了更好的抑制拟杆菌增殖的效果(P<0.05)。
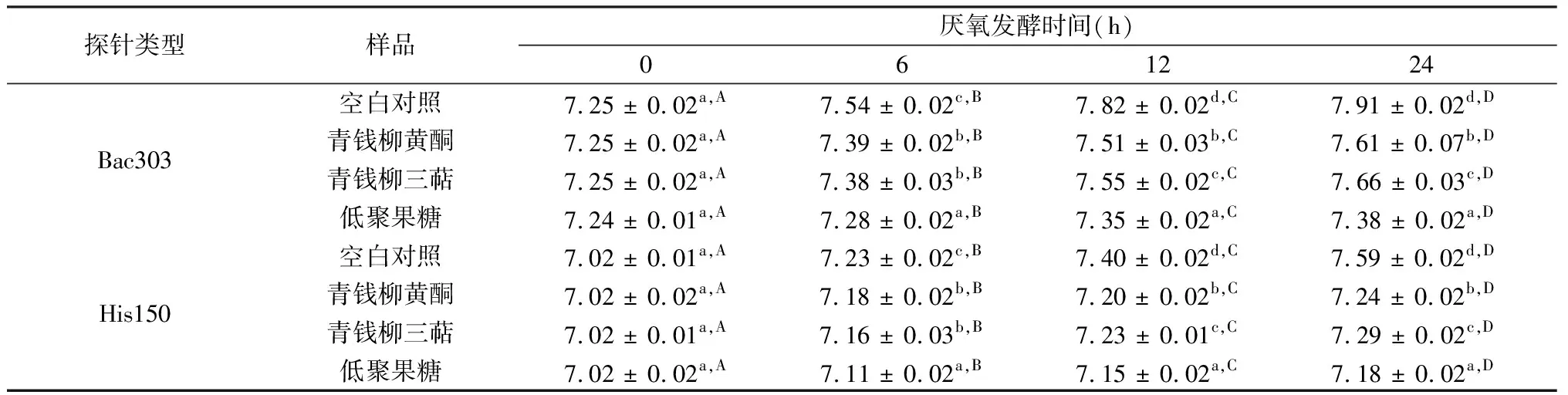
表3 青钱柳黄酮和三萜体外厌氧发酵0、6、12、24 h后拟杆菌和梭状菌的数量 lg(CFU/mL)
2.4 青钱柳黄酮和三萜对肠道总菌群数的影响
表4为以青钱柳黄酮和三萜为底物时,不同发酵时间肠道总菌群数的变化。由表中可以看出,随着发酵时间的延长,肠道总菌群数量在逐渐增多,在各组样品中,不同发酵时间的肠道总菌群数,均具有差异显著性(P<0.05),同时各组样品对肠道内总菌群的影响差异不显著(P>0.05),说明青钱柳黄酮和三萜基本不会影响肠道内总菌群的数量。结合前面的实验数据可以得出,青钱柳黄酮和三萜只是改变了肠道内的菌群结构,而对总菌群数量无显著影响。

表4 青钱柳黄酮和三萜体外厌氧发酵0、6、12、24 h后总菌群的数量 lg(CFU/mL)
3 结 论
人体消化道内的正常微生物群大部分寄居于结肠内,其数量达到1014个以上。双歧杆菌的部分菌属(Bifidobacteriumspp.)、乳酸菌/肠球菌的部分菌属(Lactobacillus/Enterococcusspp.)是两类对人体有益的微生物。此外,绝大多数的拟杆菌以及梭状菌对维持肠道的生态平衡也具有重要作用,它们是在人体肠道内长期定植的菌群,但是其中某些种属数量过多会破坏人体肠道内菌群平衡,引发腹泻、感染等疾病[8]。
益生元往往通过刺激人体肠道内有益菌的生长,如双歧杆菌、乳酸菌等而对人体健康起到积极的作用[13]。膳食消费不被人体消化的益生元,对于肠道内双歧杆菌数量的增殖具有促进作用[14]。在目前研究中,低聚果糖对有益菌的生长起到了最明显的增殖作用,同时青钱柳黄酮和三萜对于肠道有益菌(双歧杆菌和乳酸菌)的生长也起到了类似的促进作用。而且,青钱柳黄酮和三萜对于肠道中拟杆菌和拟杆菌等的生长产生了抑制作用。有研究报道,蓝莓多酚提取物能促进双歧杆菌和乳酸菌在肠道内的定殖,从而竞争性的抑制病原菌在肠道内的黏附和定殖[15]。
肠道细菌对于植物多酚在人体肠道内的生物转化起到了重要作用[16],因此植物多酚可能是肠道细菌发酵利用的合适底物。肠道细菌在肠道内可以通过对于酚类化合物去糖基化从而释放苷元,同时催化甲氧基脱甲基反应。多酚的代谢产物可以到达结肠被肠道微生物利用,从而对肠道微生物的数量以及构成产生影响[17]。肠道内的双歧杆菌、乳酸菌等益生菌通过调节肠道微生态平衡来发挥益生作用,同时双歧杆菌、乳酸菌等有益菌的增殖,可以有效抑制大肠内氨、粪臭素以及胺类致癌物等的形成[18]。青钱柳黄酮和三萜发挥益生元功能,可能的机理是其作为抗氧化剂调节代谢活动中产生的氧化应激,从而为肠道中有益菌生长和繁殖提供更有利的环境[19]。一般而言,肠道内双歧杆菌和乳酸菌等有益菌数量的增加可以更加有效的减少氨、粪臭素的形成,增加有机酸的产生,降低结肠及粪便的pH值,抑制病原菌的生长。因此,青钱柳黄酮和三萜对肠道微生态的调节发挥了积极作用。
本实验以中国特产青钱柳为原料,制备得到了高纯度的青钱柳黄酮和三萜。体外厌氧粪样混合培养实验结果表明,青钱柳黄酮和三萜对双歧杆菌、乳酸菌都有较好的增殖作用,并对拟杆菌、梭状菌的生长有一定的抑制作用。青钱柳黄酮和三萜只是改变了肠道内菌体的组成,对总体菌群的数量基本没有影响。在厌氧发酵24小时后,青钱柳黄酮对有益菌的增殖作用最强,同时相对于三萜显著抑制了拟杆菌和梭状菌的增殖。青钱柳黄酮及三萜类物质在体内的生物转化,可能对于调节肠道的菌群平衡有着重要的作用,相关肠道代谢衍生物的方面,还需要进一步的研究。本实验说明青钱柳黄酮和三萜可以通过影响肠道菌群结构,对于积极调节宿主肠道微生态,为甲基化儿茶素的作用机理研究及深入开发利用奠定了良好基础。
