新疆野苹果作为苹果砧木利用的研究进展
2019-06-11吴玉霞覃伟铭何天明
郑 点,吴玉霞,覃伟铭,何天明
(1. 新疆农业大学 林学与园艺学院,新疆 乌鲁木齐830052; 2. 新疆巴音郭楞蒙古自治州沙依东园艺场,新疆 库尔勒841000)
新疆野苹果[Malussieversii(Ledeb)Roem],又名塞威士苹果,主要分布在中亚诸国和我国新疆的伊犁、塔城地区[1]。在过去的几十年里,新疆野苹果在我国陕西、山东、河北、新疆等苹果产区常作为苹果砧木和园林绿化树种被广泛利用[2-5]。新疆野苹果根系发达,生产上普遍表现耐旱、耐寒、耐瘠薄、耐盐碱性等优良抗性[6]。过去30年来,为了更好地利用这一野生资源作为苹果砧木,许多学者从新疆野苹果对生物逆境和非生物逆境的抗性和以及嫁接亲和性角度进行了广泛研究,在抗逆机理及其评价指标方面积累了大量文献。本文对有关新疆野苹果作为苹果砧木使用密切相关的抗病性、抗虫性、耐旱性、耐寒性、耐盐碱性等性状的研究文献进行了归纳和总结,兹将有关结果报道如下,以资参考。
1 新疆野苹果对两种生物逆境胁迫的抗性研究
1.1 苹果小吉丁虫
当新疆野苹果作为苹果砧木来利用时,我们主要关注其症状表现在枝干或根系部位的生物胁迫类型,而不在于叶片和果实部位。正因如此,新疆野苹果对蛀干性害虫苹果小吉丁虫(AgrilusmaliMatsumura)的抗性研究首先成为我们关注的焦点。
苹果小吉丁虫为害新疆野苹果是外来物种入侵导致生态灾害的典型案例。在林佩钧先生2000年出版的《天山野果林综合研究》一书中,并无此虫对野苹果为害的记载[1]。实际上,苹果小吉丁虫为害野苹果最早的公开报道始于1999年[7]。
笔者对伊犁新源县喀拉布拉乡和巩留库尔德宁镇的400株野苹果单株做了定点调查,没有发现一株完全抗虫的单株,并且在野苹果分布区域内的农家种植的栽培苹果如‘富士’、‘元帅’等品种也无一幸免。梅闯等在新源县改良场做的野外调查表明新疆野苹果群体中存在高抗苹果小吉丁虫单株类型,高抗类型和低抗类型植株在枯枝率及总虫口数、羽化孔数等抗虫指标间差异较大。但定义为高抗的5株单株幼虫数为0的只有XY-90,但这一单株具有一定量的羽化虫数,所以绝对抗苹果小吉丁虫的野苹果单株迄今尚未发现[8]。
因此,将新疆野苹果作为砧木品种引用时,苹果小吉丁虫的检疫工作应引起足够重视,因为该虫不仅为害作为砧木用的新疆野苹果,也不同程度为害主栽苹果品种(即接穗部分)[9]。以下案例值得借鉴:2006年青海尖扎县有45%的苹果园受苹果小吉丁虫为害,严重的果园有60%的苹要树受害[10]。甘肃平凉1993年苹果小吉丁虫为害率16%,1997年达到100%[11]。
1.2 土壤连作障碍(苹果再植病)
我国人口多,耕地面积有限,通过在农地上新栽苹果树以发展苹果产业颇不现实。加上苹果老产区进入老龄化的果园比例越来越高,在老果园上重新建园产生的土壤连作障碍(苹果再植病)严重制约着苹果的更新换代。美国苹果产业也面临着同样问题,筛选抗病品种成为产业发展的刚性需求。Isutsa等从总生物量、根坏死、根部真菌和根腐线虫(PratylenchuspenetransCobb)4个标准评价苹果种内或种间杂交对连作障碍的抗病和耐病作用,发现所测试的品种都不抗苹果再植病(Apple replant disease, ARD),但新疆野苹果[Malussieversii(Ledeb)Roem]表现出明显耐病[12]。Fazio等也在其研究中发现苹果砧木中存在抗连作障碍的遗传基因,希望利用新疆野苹果种质获得抗连作障碍的砧木[13]。
我国学者王元征等对包括新疆野苹果在内的5种苹果砧木幼苗对连作土壤适应性差异进行了研究,发现连作对5种苹果砧木的外在影响表现为生物量、株高地径的下降,其中平邑甜茶适应性最强,新疆野苹果次之。生理指标中如CAT酶活性的影响在平邑甜茶中表现最小,新疆野苹果次之,从另一方面说明连作对砧木根系细胞造成的伤害中新疆野苹果受到的影响较小[14]。
中外学者的相关研究均表明新疆野苹果是一个较耐苹果再植病的苹果砧木品种,这为我国老苹果产区选择抗再植病的砧木提供了可能。
2 新疆野苹果对几种非生物逆境的抗性研究
2.1 抗寒性
作为苹果砧木使用的新疆野苹果,生产实践中对其抗寒性的要求并不高。因为我国大部分苹果适生区如黄土高原和胶东半岛的冬季极端低温一般要高于新疆野苹果的原生境天山山区。新疆野苹果分布于新疆境内土层薄、高度差大、生境脆弱的天山山坡地,其原生境温度日较差较大,极端低温可达-30 ℃以下,正因此新疆野苹果具有一定的抗寒能力[1]。
实验室的研究也证实了这一点。文玉珍等采用低温胁迫(4 ℃)处理新疆野苹果离体叶片,分别测定不同胁迫天数下的离体叶片的过氧化物酶活性、丙二醛含量、脯氨酸含量和氧化物歧化酶活性等生理指标。表明新疆野苹果离体叶片在低温胁迫下,体内的保护性酶以及渗透调节物质会大量积累用来抵御其在低温胁迫下所收到的伤害,由此说明新疆野苹果具有一定的耐寒能力[15]。王泽华等采用电导法和叶片解剖结构分析对不同生境下的新疆野苹果进行了抗寒性比较研究,参考枝条相对电导率高低,结合半致死温度,测量其叶片厚度、上表皮厚度、栅栏组织厚度等指标,得出新疆野苹果不同生境下抗寒性由强到弱排序依次为:托里县野苹果>额敏县野苹果>巩留县野苹果>霍城县野苹果﹥新源县野苹果。由此,将抗寒要求作为第一指标选择野苹果作为砧木采种时,托里县野苹果可作为首选[16-17]。张军等[18]用电EIS(电阻抗图谱法)法对新疆巩留县和新源县2个居群的新疆野苹果的当年生枝的抗寒性进行了研究,证明巩留居群的抗寒性高于新源居群,与王泽华的上述研究结果相吻合。
对新疆野苹果与其它苹果砧木品种的抗寒性有多位学者做了比较研究。赵玲玲等了测定了山定子[Malusbaccata(Linn.) Borkh.]、新疆野苹果[Malussieversii(Ledeb)Roem]、烟台沙果[M.prunifoli(Willd) Borkh]和莱芜难咽(M.micromalusMakino)4种苹果砧木在低温胁迫下的叶片的多胺含量,由此对其耐寒性进行对比研究。结果表明4 种苹果砧木耐寒性从高到低依次为:山定子>新疆野苹果>莱芜难咽>烟台沙果[19]。刘君等在新疆伊犁地区通过果园调查方式对不同砧木的抗寒性进行了比较,发现苹果不同砧木在伊犁苹果产区抗寒性大小表现为:GM256>新疆野苹果>辽砧2号[20]。马子华利用活体电阻法对黄海棠、新疆野苹果和怀来海棠三种苹果砧木抗寒性的研究表明:黄海棠最强,怀来海棠次之,新疆野苹果最差[21]。
2.2 抗旱性
谭冬梅采用20%聚乙二醇对平邑甜茶(MalushupehensisRehd)和新疆野苹果[Malussieversii(Ledeb)Roem]模拟干旱胁迫处理后,发现新疆野苹果叶片保持细胞膜完整性的时间比平邑甜茶长。对叶片进行DNA含量、0.1% NBT染色、相对导电率和RNA含量测定等生理测定表明,平邑甜茶再处理后DNA和RNA含量有所下降,而新疆野苹果下降速度明显缓慢。由此可见,在干旱胁迫下,新疆野苹果先发生程序性死亡以应对干旱,其抗旱的机制要强于平邑甜茶[22]。张静等采用电导法和检测MDA(丙二醛)含量对新疆野苹果、平邑甜茶、八棱海棠、‘M26’、‘M9’的抗旱性进行了鉴定。最终将这几种苹果砧木抗旱性分为四类:不抗、低抗、中抗和高抗。不抗:‘M9’;低抗:‘M26’;中抗:平邑甜茶和八棱海棠;高抗:新疆野苹果。按照砧木抗旱性由强到弱依次排序为:新疆野苹果>平邑甜茶>八棱海棠>‘M26’>‘M9’[23]。从以上两例研究结果初步分析,新疆野苹果的抗旱性是其最为优异的一个抗逆性状,在干旱地区如黄土高原地区发展苹果产业时应作为砧木品种的首选。
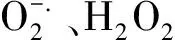
其实,抗旱性也是新疆野苹果抗逆性状研究中最为深入的一例,在分子水平上对其抗旱机理的研究有了率先突破。徐彥通过抑制性差减杂交的方法构建了新疆野苹果根组织在干旱胁近下诱导表达差减文库和抑制表达差减文库,测定鉴定出16个生物学功能在内的268个有效的UniESTs[26]。Zhao等克隆了新疆野苹果的MsDREBA5基因,并发现包括干旱胁迫在内的非生物逆境胁迫可诱导该基因的表达,而盐胁迫则抑制该基因的表达[27]。Peng等检测了新疆野苹果等苹果砧木在干旱胁迫下促分素原活化蛋白激酶(MAPK)基因的表达变化,发现其表达水平与抗旱性呈正相关,抗旱性最强的新疆野苹果MAPK表达水平最高[28]。
2.3 抗(耐)盐性
为了筛选高抗盐苹果砧木,多位研究者在不同试验条件对不同要砧木的抗盐性做了鉴定。刘兵等采用含NaCl的营养液浇灌珠美海棠[Maluszumi(Matsum)Reder]和新疆野苹果[Malussieversii(Ledeb)Roem]组培苗,比较了它们的抗盐性,通过综合比较盐胁迫下珠美海棠和新疆野苹果各项生理生化指标的变化,得出珠美海棠的耐盐性强于新疆野苹果的结论[29]。Lan等比较了八棱海棠(MalusrobustaRehd)和新疆野苹果[Malussieversii(Ledeb)Roem]通过砂培在不同盐浓度下叶片的组织解剖结构变化。通过光学显微结构表明,对比在盐胁迫的环境下,八棱海棠和新疆野苹果的叶片厚度,海绵组织、上下表皮发生不同程度的加厚,栅栏层组织厚度减小。八棱海棠比新疆野苹果叶片细胞结构损伤小,更能够完整的保持细胞结构。八棱海棠对盐胁迫的适应性要高于新疆野苹果,对盐胁迫表现出较强的适应性[30]。尹蓉通过水培研究的方法,通过抗逆性相关生长性状结合电导率、丙二醛含量、抗氧化物酶研究,也表明八棱海棠为耐盐性强的苹果资源,而新疆野苹果则归为耐盐性中等的一类[31]。于玮玮等的研究表明新疆野苹果幼苗在盐胁迫下可通过积累脯氨酸和可溶性糖,提高 SOD和 POD 的活性,以减缓盐胁迫对植株的伤害且具有一定的耐盐性[32]。
林冰冰等2016等对不同地点不同株系采集的新疆野苹果单株的抗盐性进行了深入研究。发现新疆野苹果耐盐碱性居群间变异幅度不仅小于种间变异,也小于居群内3个性状分离程度,且同一居群的株系间各个性状的种间差异相当。说明苹果属资源抗性种间差异明显,且新疆野苹果种内抗逆性分布虽然广泛,居群内抗逆性分离在株系内和株系内分离更严重[33]。
2.4 抗缺铁性
我国苹果产区有大面积的石灰性钙质土壤,因土壤铁被大量固定而有使土壤有效铁含量偏低,极易发生叶片缺铁性黄化症状。筛选铁高效砧木成为根治这一顽疾的良方,中国农业大学在这一领域做了大量工作。韩振海研究表明,在苹果属的不同砧木中,小金海棠(M.XiaojinensisCheng M.H.et Jiang N.G.)表现出较高的抗缺铁特性,八棱海棠吸收和利用铁素能力稍弱,而山定子、青海花叶、新疆野苹果却极易发生失绿[34]。吴玉霞等通过溶液培养,施以缺铁胁迫,评价了4种砧木的抗铁性:相对于新疆野苹果、平邑甜茶和黄太平而言,八棱海棠表现出较强的抗缺铁特性,是一个理想的铁高效苹果砧木基因型[35]。焦晓峰在田间做的调查也证实了吴玉霞水培试验的结果:小金海棠、八棱海棠和新疆野苹果三种砧木,在南疆钙质土壤上小金海棠和八棱海棠的抗缺能力最强,新疆野苹果发生黄化症状明显,表现出抗缺铁能力较弱。以上研究表明,在极易发生缺铁黄化的地区选用苹果砧木时,新疆野苹果要慎用[36]。
3 嫁接亲和性研究
新疆野苹果目前生产应用的主要途径是作为砧木资源,所以与主栽品种的嫁接亲和性是一个重要性状。李开花等以新疆野苹果为基砧,采用劈接法嫁接了‘新冠’、‘红肉苹果’和‘新帅’,根据嫁接成活率、接穗萌芽率等指标评价了嫁接亲合性。结果表明上述三个苹果品种嫁接在新疆野苹果上其成活率均在80%以上,对比嫁接口愈合程度,认定‘红肉苹果’与新疆野苹果嫁接亲合力最强,嫁接口比较平滑,其次为‘新冠’和‘新帅’,均表现出一定量的大脚现象和瘤状突起,表明一定的不亲和现象[37]。本课题组引进了大量新疆伊犁州农科院以新疆野苹果为基砧、以矮化砧GM233、T337等中间砧嫁接繁殖的苹果苗木,定植于位于新疆巴音郭楞蒙古自治州沙依东园艺场,苗木生长一年,成活率均在80%以上,嫁接口无明显裂皮症状,说明新疆野苹果与矮化砧木也有良好的嫁接亲和性。
4 新疆野苹果作为苹果砧木利用研究的前景展望
综上所述,新疆野苹果作为砧木利用的文献多集中在抗逆生理指标的测定以及表观性状的调查方面,从分子水平上阐明新疆野苹果的抗逆机制的研究才刚刚起步。目前,随着分子生物学和基因工程的飞速发展,从分离抗逆功能基因、利用分子手段确定抗逆性评价指标已经可能。因此,在今后的研究中,分子机制的研究是一个方向趋势。最近,有关新疆野苹果全基因组测序的研究成果为这一领域的拓展奠定了基础[38]。
另外,目前关于新疆野苹果非生物逆境的抗性研究多在水培、砂培和组织培养条件下完成,与大田条件相差甚远,研究成果对实际的商业化栽培指导意义有限。因此,在今后的研究中,应重视在大田条件特别是在盐碱重的地区新疆野苹果的抗性研究。
