食用菌异迟眼蕈蚊苏云金芽孢杆菌筛选及杀虫蛋白基因鉴定
2019-05-23王帆帆曲绍轩林金盛李辉平侯立娟蒋宁骆昕马林韩巨才
王帆帆,曲绍轩,林金盛,李辉平,侯立娟,蒋宁,骆昕,马林,3*,韩巨才
(1.山西农业大学农学院,山西 太谷 030801;2.江苏省农业科学院蔬菜研究所,南京 210014;
3.江苏省高效园艺作物遗传改良重点实验室,南京 210014)
眼蕈蚊科(Sciaridae)是食用菌栽培中危害较广的一类虫害,有6属27种[1],其中异迟眼蕈蚊(Bradysia difformis)是优势种群之一[2-3]。异迟眼蕈蚊幼虫直接潜入食用菌菌袋,取食菌丝和子实体,且排出粪便滋生病菌[4];其成虫飞行携带的病原菌、线虫、螨虫等导致二次侵染[5]。目前针对异迟眼蕈蚊的防治方法主要有物理防治和化学防治,但由于物理防治多针对成虫[6],对幼虫无效,而化学防治会不同程度地抑制菌丝生长[7],且存在残留、抗性、再猖獗问题,所以生物防治成为研究食用菌虫害防控的热点。
苏云金芽孢杆菌(Bacillus thuringiensis,Bt)是生物农药中应用最为广泛的一种微生物杀虫剂。Bt在形成芽孢时可以产生具有杀虫活性的杀虫晶体蛋白(insecticidal crystal proteins,ICPs)[8],幼虫取食后中毒死亡。近年来,有关Bt杀虫晶体蛋白的结构、功能及其特异性研究已较为深入。经X-射线晶体衍射的方法,学者们已经得到其三维结构,主要包括3个结构域,因结构不同而各有分工,分别是参与孔道形成的DomainⅠ和参与受体结合的DomainⅡ和DomainⅢ[9]。有关特异性研究表明,不同的杀虫晶体蛋白对双翅目、鳞翅目、鞘翅目及线虫等有相应的杀虫活性[10],其中对双翅目类害虫有活性的主要包括cry2、cry4/10、cry11、cry16-17、cry19、cry24-25、cry27-29、cry30、cry32ab、cry32cd、cry40、cyt1、cyt2等基因控制的蛋白[11]。已报道苏云金芽孢杆菌以色列变种(Bacillus thuringiensisvar.israelensis,Bti)制剂对双孢蘑菇栽培过程中的眼蕈蚊有较好的控制效果[12],但有关Bt防治食用菌异迟眼蕈蚊及其杀虫晶体蛋白鉴定的报道较少。由于Bt对非目标性生物安全,具有专一、高效和对人畜无害的优点,备受人们的青睐。本文通过菌株筛选,发现了对异迟眼蕈蚊有明显杀虫效果的Bt菌株,明确了其主要的杀虫晶体蛋白类型,为应用生防菌Bt防治食用菌虫害奠定了基础。
1 材料与方法
1.1 供试昆虫、菌种、培养基及试剂
供试昆虫为在实验室内用平菇出菇袋饲养的3龄异迟眼蕈蚊幼虫,供试食用菌为平菇(Pleurotus ostreatus),品种为8801。二者均由江苏省农业科学院蔬菜研究所食用菌实验室提供。
分离培养基:酵母粉5g,蛋白胨10g,氯化钠10g,醋酸钠34 g,水1 L。产孢培养基:牛肉膏5 g,蛋白胨10 g,琼脂20 g,水1 L。液体发酵培养基:牛肉膏3 g,蛋白胨5 g,葡萄糖10 g,水1 L。LB(Luria-Bertani)培养基:蛋白胨10 g,酵母粉5 g,氯化钠10 g,水1 L。
EZUP柱式细菌基因组DNA抽提试剂盒购自生工生物工程(上海)股份有限公司,聚合酶链式反应(polymerase chain reaction,PCR)相关试剂购自宝生物工程(大连)有限公司,其他试剂均为国产进口分析纯试剂。
1.2 土样采集
土壤样品采集对象为南京市紫金山不同海拔和不同植被覆盖下的土壤。采用棋盘法进行采集,先铲去表层土1~2 cm,再取土壤样品约50 g,放入自封袋,备用。
1.3 Bt菌株的筛选
采用醋酸钠-抗生素法分离Bt菌株[13]。称土样2 g于含玻璃珠和分离培养基的锥形瓶中,在培养基中加入终质量浓度为400 μg/mL的青霉素钠盐和硫酸庆大霉素,于190 r/min、30℃摇床上培养4 h。培养结束后取土壤悬浮液5 mL,80℃水浴20 min,悬浮液稀释10倍后取500 μL,涂布于产孢培养基平板上,30℃静置培养3 d。挑取表面粗糙、边缘不规则的菌落,分别制作玻片样本,进行石炭酸复红染色,油镜观察菌落形态,选择有菌体、孢子和伴孢晶体的菌株纯化、保存。
1.4 Bt菌株室内毒力测定
采用药液浸渍饲喂法测定野生Bt菌株对3龄异迟眼蕈蚊幼虫的毒杀效果。为了减少制剂与菌株在浓度上的差异,分离提纯Bti制剂(苏云金芽孢杆菌以色列变种可湿性粉剂,由武汉科诺生物科技股份有限公司提供)中的菌株作为对照菌株。将1.3中纯化的Bt菌株和Bti对照菌株置于LB液体培养基中,于30℃、190 r/min条件下振荡培养过夜,按10%的菌液量转接至液体发酵培养基中,于30℃、190 r/min条件下振荡培养72 h。用不同菌液分别浸泡大小一致的平菇子实体10 min,然后用滤纸吸干表面多余菌液,放入直径9 cm的培养皿中。培养皿底部放一层滤纸,在滤纸上滴加700 μL相应菌液保湿。每皿挑30头幼虫,以清水处理的菇片作为空白对照,每个处理重复3次。室温培养72 h后观察并记录死亡率。
1.5 Bt菌株对食用菌菌丝的影响
平菇菌丝稀释液与冷却至35℃左右的液态LB固体培养基以体积比1∶40混匀,制成平板。选择对异迟眼蕈蚊有明显杀虫活性的菌株,按1.4中所述Bt菌株液体发酵方法制备发酵液,分别取5 μL滴加至平板。每个处理重复3次,用无菌水作对照。25℃恒温静置培养3 d,观察菌丝生长状况。
1.6 Bt菌株16S rDNA分析
选取对异迟眼蕈蚊有明显杀虫活性且不影响菌丝生长的菌株,在LB液体培养基中振荡培养过夜(28℃,190 r/min),提取菌株基因组DNA。用细菌16S rDNA通用引物27F和1492R进行PCR扩增。扩增产物由擎科生物工程(南京)股份有限公司进行测序,测序结果通过在NCBI(https://www.ncbi.nlm.nih.gov/)上进行BLAST比对,初步确定菌株分类学地位。采用Mega 6.0分子生物学分析软件中的邻位相连法构建菌株系统发育树。
1.7 Bt菌株杀虫蛋白基因型的聚合酶链式反应-限制性片段长度多态性(polymerase chain reaction-restricted fragment length polymorphism,PCR-RFLP)方法鉴定
以Bt菌株基因组DNA为模板,用13对引物(表1)进行PCR-RFLP扩增[14],确定各菌株含有的对双翅目昆虫有杀虫活性的毒蛋白基因型数量及类别。
2 结果与分析
2.1 Bt菌株的筛选结果
本试验共采集33份土壤样品,获得了82个与芽孢杆菌菌落形态相似的单菌落,通过玻片显微镜观察,获得了17株可产生伴孢晶体的菌株,如图1所示。

表1 杀虫晶体蛋白基因型鉴定引物序列Table 1 Primer sequences used for identification of insecticidal crystal protein genotypes
2.2 Bt菌株室内毒力测定分析及对食用菌菌丝生长的影响
用筛选的17株Bt菌株发酵液饲喂异迟眼蕈蚊72 h后,发现其具有不同程度的杀虫效果,如图2所示。其中:有7个菌株的校正死亡率高于50.00%,且高于对照菌株Bti;菌株70-4的校正死亡率高达100.00%。对平菇菌丝生长的影响试验结果(图3)显示,7个校正死亡率高于50.00%的Bt菌株对菌丝均无明显抑制作用。
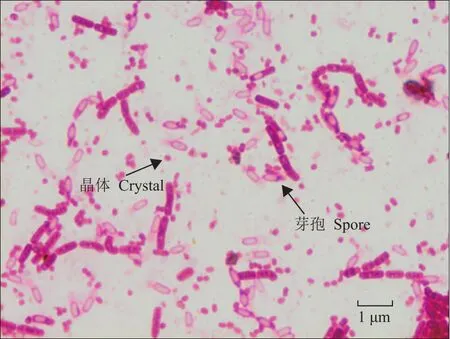
图1 Bt菌株芽孢、晶体形态Fig.1 Morphology of spore and crystal of Bt strain

图2 Bt菌株对异迟眼蕈蚊幼虫的室内毒力测定结果Fig.2 Results of toxicity test of Bt strains against the larvae of B.difformis

图3 Bt菌株对平菇菌丝生长的影响Fig.3 Effect of Bt strains on hypha growth of P.ostreatus
2.3 Bt菌株16S rDNA分析
16S rDNA片段测序结果显示,菌株69-5、70-2、70-4、71-1、72-3、83-1、214-6和Bti的16S rDNA序列大小在1 368~1 459 bp之间。将7个分离菌株和Bti的16S rDNA序列与在GenBank(https://www.ncbi.nlm.nih.gov/genbank/)中已发表的序列进行同源性比对,如图4所示:各分离菌株与B.thuringiensis菌株ATCC-10792和FJAT-45929同源性最高,达99%以上,可初步确认这7个菌株在分类学上属于苏云金芽孢杆菌。
2.4 Bt菌株杀虫蛋白基因型鉴定
对分离的7个Bt菌株和Bti的毒蛋白基因型鉴定结果(表2)初步表明:7个菌株包含cry4/10、cry11、cyt1和cyt2等4种基因型,其中菌株69-5和70-2含有的毒蛋白基因型较多,为cry4/10+cry11+cyt1+cyt2复合基因型,在室内毒力活性测定中杀虫活性最高的70-4菌株仅含有cry11和cyt12种毒蛋白基因型。对照菌株Bti含有cry4/10、cry11、cry30、cyt1和cyt2等5种基因型。8个菌株都含有cry11基因型,菌株69-5、70-2和Bti中包含cry4/10基因型,其余菌株基因组合类型也各不相同。
3 讨论
在实际生产中,对食用菌迟眼蕈蚊的防治主要集中于化学防治,噻虫嗪[15]、除虫脲[16]和苦参碱[17]等均具有良好的杀虫效果,但易影响食用菌菌丝的生长。近年来,学者们纷纷转向生防菌的研究,Bt菌株BRC-LLP29[18]和JQ23[19]对眼蕈蚊的防治率均达70%以上,但菌株BRC-LLP29对香菇等食用菌菌丝生长有明显抑制作用。菌株70-4对平菇菌丝生长无影响且对异迟眼蕈蚊校正死亡率达100%,防治效果明显。本试验中7个分离菌株的校正死亡率均高于对照菌株Bti,该制剂主要用于防治双翅目中库蚊科、蚊科等虫害,防治对象不同,会导致防治效果存在一定差异。目前,有关食用菌异迟眼蕈蚊的生物防治鲜有报道,故本试验中的分离菌株具有很好的应用价值。

图4 7个Bt菌株的16S rDNA系统发育树Fig.4 16S rDNAphylogenetic tree of seven Bt strains
我国的Bt菌株资源丰富,迄今为止已有70个血清型,83个亚种[20]。针对新Bt菌株杀虫晶体蛋白基因型的发掘与鉴定也随之展开[21]。已报道菌株FPT3[22]对眼蕈蚊杀虫活性达80%以上,其表达毒蛋白分子质量主要在70 kD左右,但未对其进行相关基因型的筛选和鉴定;IBARRA等[23]对Bt菌株筛选的结果显示,菌株LBIT315对双翅目埃及伊蚊的毒性最高,致死中质量浓度(lethal concentration of 50%,LC50)仅 4.12 ng/mL,为cry4A+cry4B+cry11+cyt1+cyt2组合基因型;SHISHIR等[24]筛选的菌株Btk HD-73和Bt JSc1对双翅目瓜实蝇的活性分别达96%、93%,共含有cry1、cry2、cry93种基因型。但有关双翅目食用菌异迟眼蕈蚊毒蛋白基因型的筛选鲜有报道。本研究筛选的菌株69-5和70-2为cry4/10+cry11+cyt1+cyt2复合基因型,菌株70-4为cry11和cyt1复合基因型,但70-4的杀虫活性高于69-5和70-2,表明毒蛋白基因cry11和cyt1对食用菌异迟眼蕈蚊杀虫活性的提升起着至关重要的作用,且菌株所含基因型的多样性可以决定杀虫活性的广谱性[25],但与其毒力大小并不存在一定的关系。
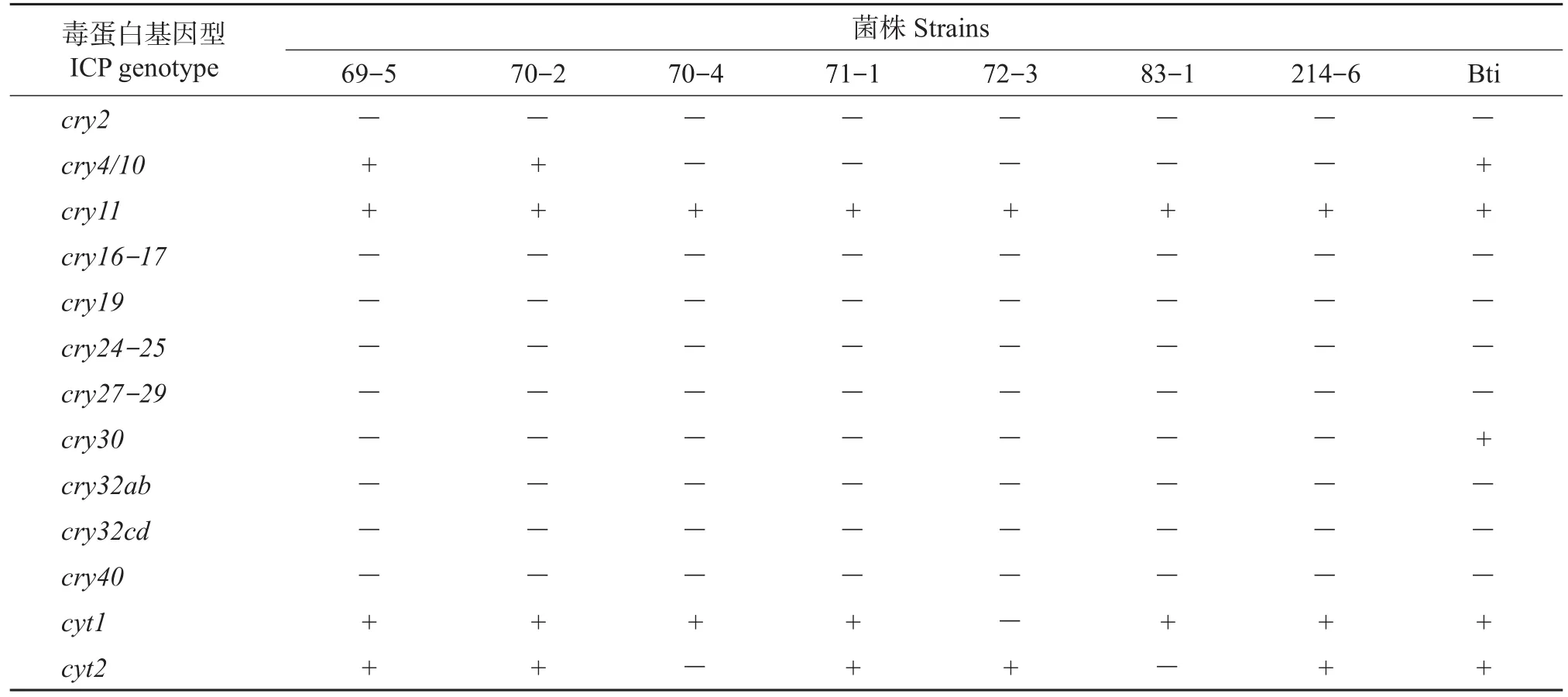
表2 Bt菌株杀虫蛋白基因型的鉴定Table 2 Identification of insecticidal crystal protein(ICP)genotype in Bt strains
目前,相关研究表明毒蛋白的杀虫活力主要取决于以下3点:1)不同Bt亚种在孢子形成阶段会合成特殊的蛋白酶,相关酶对某些毒蛋白进行修饰后会减弱或增强其杀虫活性[26]。2)原毒素在昆虫中肠内经不同的蛋白酶处理后,产生的多肽片段长短不同,会直接影响其毒力,每种毒蛋白都有特定的酶作用位点[27]。3)不同菌株间相同基因型表达量的不同也会明显影响其杀虫效果。本试验分离的7个菌株的校正死亡率明显高于Bti对照菌株,推测主要原因在于菌株存在种间差异,且防治对象不同,而具体原因及其机制有待于进一步的研究。
Bt杀虫剂已经在其他作物上表现出良好的杀虫活性和对环境的友好性,对食用菌眼蕈蚊的室内毒力测定和对食用菌菌丝的影响研究亦可看出其在食用菌虫害防控中的应用前景,可通过发酵条件优化[28]和药剂剂型开发[29]等方式提高杀虫活性。但由于食用菌在相对密闭的小环境中栽培,虫害种类相对较少,而长期大量使用同种类的Bt杀虫剂会导致害虫耐药性的产生[30]。因此,可通过毒蛋白结构域交换、定点突变、肽添加等不同的策略增强Bt杀虫剂对特定害虫的毒性[31]。下一步将致力于将各菌株的杀虫晶体蛋白分别进行表达载体的构建,尤其是对70-4菌株中cry11和cyt1对食用菌异迟眼蕈蚊的杀虫活性进行深入研究。
