烟株发育过程中硝酸盐积累与调控的研究进展
2019-05-20冯雨晴杨惠娟史宏志
冯雨晴,杨惠娟,史宏志
河南农业大学烟草学院/烟草行业烟草栽培重点实验室/烟草农业减害研究中心,郑州市金水区文化路95号 450002
硝态氮是植物根系吸收利用的主要无机氮素形态,以硝酸盐的形式广泛存在于环境中[1]。硝酸盐作为一类重要的含氮化合物,其含量的高低不仅直接影响烟叶产量和质量的形成,更对烟叶安全性和人体健康有着直接或间接的影响。现代医学证明,人体摄入的硝酸盐经过体内微生物作用还原成的亚硝酸盐可导致血液的载氧能力下降;另一方面,亚硝酸盐还可与人体的次级胺,如仲胺、叔胺及氨基酸反应,在胃腔中(pH=3)形成强致癌物—亚硝胺,从而诱发人体消化系统癌变[2-4];更重要的是,烟草叶片中的硝酸盐是烟叶和烟气中强致癌物质烟草特有亚硝胺(TSNA)的重要前体物[5]。研究表明,TSNA主要在调制、贮存陈化过程中逐步形成和积累,在调制过程中硝酸在微生物作用下还原为亚硝酸,进而与生物碱发生亚硝化反应[6],在烟叶贮藏过程中硝酸盐在高温条件下可产生气态氮氧化物,进而与生物碱反应生成TSNAs[7-8]。TSNAs可引发动物形成肺癌、腺癌和食道癌等病变,尤其是N?-亚硝基降烟碱(NNN)和4-(N-甲基亚硝胺基)-1-(3-吡啶基)-1-丁酮(NNK)被国际癌症研究组织(IARC)列为最高级别I类致癌物[9-11]。白肋烟等晾晒烟烟叶TSNAs较烤烟高几十甚至几百倍,与其烟叶硝酸盐含量偏高密切相关,近期研究表明,烟叶硝酸盐含量与 TNSAs 积累呈现出显著性直线相关[5],减少烟叶中硝酸盐积累量对于降低卷烟制品中NNN和NNK的含量至关重要[12]。关于TSNA的相关研究一直是国际烟草研究的热点之一,减少烟气TSNA含量的有效途径之一是降低烟草原料中的硝酸盐含量,这已在通过栽培、生物技术调控烟叶硝态氮含量相关研究和利用不同硝酸盐含量的烟梗来改变卷烟硝酸盐含量的研究中得到证实[13-15]。因此,控制和减少烟叶硝态氮积累对降低烟叶TSNA形成和提高卷烟安全性具有十分重要的意义。对此,笔者对烟叶硝酸盐的积累与影响因素以及硝酸盐的调控技术进行了综述,以期为后续硝酸盐的调控研究提供参考。
1 白肋烟硝酸盐代谢及积累机理
硝酸盐代谢途径及积累机制图如图1所示。
外界土壤环境中的硝酸盐通过根系表皮和皮层细胞膜上的硝酸转运蛋白(nitrate transporters,NRT)进入根系细胞内,是植物获取硝态氮(即硝酸盐)并利用的第一步,属于根部主动跨膜运输[16-17]。根系形态和硝酸盐转运效率是影响硝酸盐吸收量的关键因素,此外,光照、温度、介质pH、呼吸抑制剂和氧化磷酸化等因素对硝酸盐的吸收和转运速率也具有重要影响[18-19]。
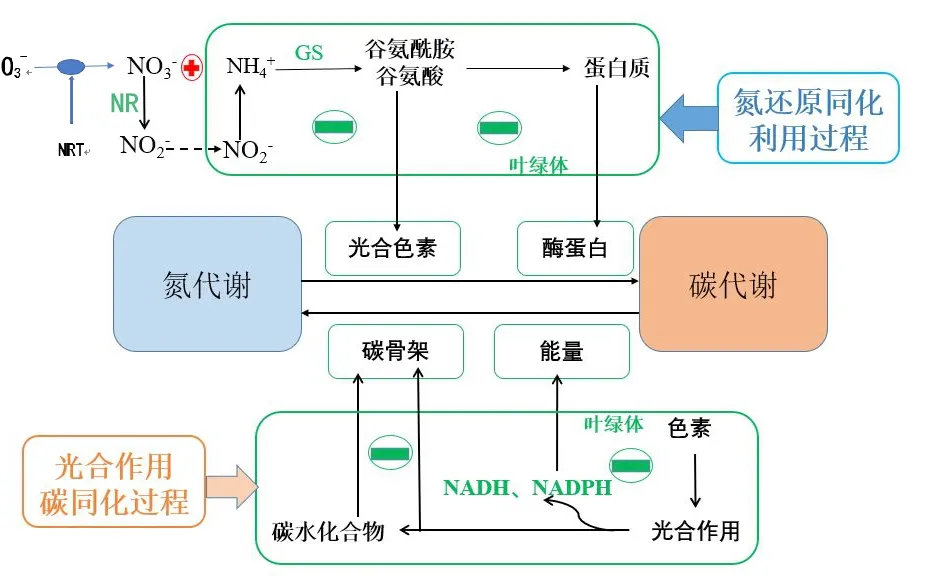
图1 硝酸盐代谢途径及积累机制图Fig.1 Nitrate metabolic pathway and accumulation mechanism
烟草属于喜硝态氮作物,对土壤中硝态氮的吸收主要是通过硝酸盐转运蛋白来完成[20]。硝酸盐的转运系统对于硝酸盐的吸收和转运至关重要,根据高等植物对的亲和力,将硝酸盐转运系统主要分为2类,即高亲和转运系统(High-affinity nitrate transport system,HATS)和低亲和转运系统(Low-affinity nitrate transport system,LATS)。当外界硝酸盐浓度高于0.5mmol/L时,低亲和转运系统在转运过程中发挥主要作用;当外界硝酸盐浓度在0.1-0.5mmol/L时,高亲和转运系统在转运过程中发挥主要作用[18]。目前,关于硝酸盐吸收和转运方面,人们多集中在NRT1、NRT2、CLCa/b和SLAH3等家族的研究。植物中具有与其对应的低亲和力硝酸盐转运蛋白NRT1和高亲和力硝酸盐转运蛋白 NRT2 两个转运蛋白家族,这两个家族均由跨膜pH梯度驱动,以协同转运机制转运和质子H+[21]。基因NRT/PTR FAILY3.1和NRT/PTR FAILY7.3属于低亲和硝酸盐转运子,基因NRT2.1和NRT2.5属于高亲和硝酸盐转运子,均参与硝酸盐转运,对硝酸盐分配和利用具有重要影响[22]。NRT1.1又名CHL1,是通过硝酸盐类似物筛选氯酸盐得到的CHL1突变体发现的第一个硝酸盐运输蛋白基因[23]。研究发现,转运子NRT1.1和NRT1.2主要在根毛和根表皮表达,NRT1.4主要在叶柄中表达,NRT1.5主要在根木质部中柱鞘细胞表达,NRT1.8主要在维管束中木质部薄壁细胞中表达,NRT1.9主要在根韧皮部伴胞细胞中表达,NRT1.7主要在老叶韧皮部伴胞细胞和筛板中表达,其中NRT1.4能够调控酸盐的动态平衡,NRT1.5、NRT1.8和NRT1.9主要参与硝酸盐的长距离运输,NRT1.7能够将硝酸盐从老叶转运到内叶和叶片中硝调控硝酸盐的源-库供应[24-26]。在模式作物拟南芥上,硝酸盐转运子基因NRT1.1多在根和地上部的生长区表达[27]。2015年在籼稻中克隆到了NRT1.1B基因,研究发现它可以加强植物对的吸收、从地下部向地上部转运、增强信号响应,以此来提高作物对氮素的利用效率[28]。目前,已从烟草中克隆获得高亲和硝酸盐转运蛋白NRT2 家族成员NRT2.2、NRT2.3、NRT2.4基因,并揭示了其在硝酸盐吸收转运过程中的作用[29-31]。
1.2 硝酸盐的还原和同化
植物的氮素同化作用多是从硝酸盐的还原同化开始的,硝酸盐被吸收进入植物根系细胞后,小部分留在根部被还原,而大部分被运送到地上部分,在硝酸还原酶(EC 1.6.6.1,nitrate reductase,NR)的作用下形成亚硝酸盐,然后被亚硝酸还原酶(EC 1.7.7.2,nitrite reductase,NiR)进一步还原成NH3,最后NH3被谷氨酰胺合成酶(EC 6.3.1.2,glutamine synthetase,GS)作用形成谷氨酰胺和谷氨酸,随后能够发生氨基转化形成各种氨基酸进入植物体代谢活动[32]。
1.2.1 硝酸还原酶的作用
NR是硝酸盐代谢途径的第一个酶,也是硝酸盐还原的限速酶[33]。NR主要分布在植物的根、茎和叶中,并定位于细胞质中,其活性大小因物种、品种、年龄、部位等因素不同而不同。硝酸还原酶属于典型的诱导酶,其活性受底物浓度、激素、光、温度、CO2和氧气等环境条件的调控。SOOD等[34]研究发现,能诱导NR活性,而则会抑制NR活性;在冬小麦上研究发现,高温能引起叶片NRA下降[35]。另外,NR基因表达水平还易受碳水化合物、亚硝酸盐、谷氨酰胺和等物质的影响。NR在光下发生去磷酸化,引起NRA升高;同时光合作用和碳水化合物可为硝酸盐还原同化过程提供必须的能量,而亚硝酸盐、谷氨酰胺和作为硝酸盐代谢产物能对其代谢过程进行反馈调节[36-37]。在拟南芥中编码NR的主要有两个基因AtNIA1和AtNIA2,这两个基因的表达受诱导,它们编码蛋白的活性也受多种因素的影响,如光照、pH、供氧量等[38-39]。
1.2.2 谷氨酰胺合成酶的作用
硝酸盐经NR和NiR还原后形成NH3,而NH3需要被谷氨酰胺合成酶(GS)同化才能完成氮素还原同化过程[40]。GS是NH3同化的中心酶,也是植物催化无机氮向有机氮转化的关键酶,对氮素利用具有重大贡献。GS对NH3的亲和力较强,能在低浓度下结合NH3,对减少细胞氨积累和避免氨损害现象有重要作用[41]。谷氨酰胺和谷氨酸是GS同化硝酸盐的产物,能为蛋白质和叶绿素等生物合成提供氮素供体。GS具有多种同工酶形式,在高等植物中,主要存在2种GS同工酶:胞质型,又称胞液型,GS1,主要位于细胞质中;叶绿体型或质体型,又称GS2,其中在叶片中主要位于叶绿体中称为叶绿体型,而在根中主要存在于质体中称为质体型[42]。有研究指出,GS1基因主要参与种子萌发时营养贮藏和叶片衰老时氮素再利用的运转工作,而GS2基因主要参与光呼吸中NH3、硝酸盐还原NH3以及循环NH3的同化利用活动[43]。
1.3 硝酸盐的贮存和再利用
植物吸收的部分硝态氮会在根部被还原为亚硝态氮,但大部分则通过蒸腾作用以形式经木质部输送到地上部的茎、叶等器官,分布于细胞的液泡和原生质中。在植物细胞中,硝酸盐多数是贮藏在液泡(储存库,storage pool,SP)中,而仅有一小部分存在于细胞质(代谢库,metabolism pool,MP)[44]。由于硝酸还原酶主要存在于原生质中,其中的硝酸盐可以迅速还原,不易累积;而液泡中的硝酸还原酶活性低,硝酸盐难以被还原,因而较易累积。这可能是造成硝酸盐累积的一个重要原因。揭示硝酸盐在液泡膜的转运调控机制,调动植物两个硝酸盐库中的硝酸盐使之得到更高程度的再利用,是提高植物氮素利用效率和降低植物体内硝酸盐含量的有效途径之一[45]。在烟草生产中,一般氮肥施用时间较早,烟株在生长中后期可能会出现缺氮或低氮情况,储存库的硝酸盐会释放到代谢库中以维持正常代谢活动。CLCa和CLCb定位于液泡膜,被认为是负责硝酸盐从液泡出入的转运子[46-47]。在拟南芥上发现,氯离子通道(CLC)基因家族中共有7位成员参与了硝酸盐在液泡膜内外的转运,改变其中的任一位成员都将影响硝酸盐在叶片中的积累[48]。另外,GS1基因在氮素转运和循环中起关键作用,该基因过表达后能明显提高氮代谢、生物量和产量,GS2基因对提高氮素利用和改善农艺性状等方面有显著影响[49]。
2 影响硝酸盐积累的因素
2.1 遗传因素
一般情况下,烟草中硝酸盐的含量因基因型而异,由烤烟的0.10%至白肋烟的3.3%不等,晒烟和雪茄烟硝酸盐含量也较高[50]。赵晓丹[51]等选取不同类型和品种烟叶进行比较,发现其硝酸盐含量差异显著。不同类型和品种间烟叶硝酸盐含量的差异多是由遗传因素控制,主要表现在硝酸盐吸收和转运、硝酸盐同化利用以及氮效率差异等方面,白肋烟氮效率低,最适施氮量达15-18kg/667m2,烤烟最适施氮量为3-6 kg/667m2,晒烟介于两者之间。李亚飞等[52]研究表明,烟叶硝酸盐积累与碳氮代谢关键基因及转录因子关系密切。
2.1.1 氮代谢关键基因
硝酸盐还原同化基因NR和GS对硝酸盐调控有重要意义,与烤烟相比,白肋烟烟叶NRA和GSA显著低于烤烟,造成其硝酸盐大量积累[52]。烟叶氮同化相关基因(NIA1、NIA2、GS1、NDHA-1)表达水平低于烤烟,是引起白肋烟烟叶硝酸盐积累的直接原因[53]。目前,已经成功从烟草中克隆出高亲和硝酸盐转运蛋白基因 NRT2 家族成员AtNRT2.2、AtNRT2.3、AtNRT2.4,并揭示了其在硝酸盐吸收转运过程中的作用[29-30,31],为进一步通过分子改良进行氮素营养调控提供了理论依据。
2.1.2 碳代谢关键基因
白肋烟是一种Yb基因突变体,其烟叶色素含量仅为普通绿色烟草的1/3左右,对光合作用不利[56]。Lewis等[57]研究表明,白肋烟Yb基因突变引起其氮效率下降,白肋烟吸收氮较多,而光合速率弱,光合产物少,因此被同化的氮素较少,多余氮未被利用以硝态氮形式储存。与烤烟相比,白肋烟烟叶碳固定和糖类物质合成相关基因(CP12-2、ATJ8、EGY1、GDHA、AGPS1、SPS2、SUS2、DPE2、TPS11)表达水平较低,造成烟叶碳水化合物的形成和积累较少,能量供应不足是硝酸盐大量积累的根本原因[52]。
2.2 发育因素
同一品种不同部位间硝酸盐含量有所差异,在植株体内,新生的部位活性强,不易积累硝酸盐,而老组织活性较弱,积累硝酸盐能力较强。试验表明,不同部位烤后烟叶硝酸盐含量表现为中部叶>上部叶>下部叶[58]。烟草叶片和烟梗在化学组成上差异较大,与调制后叶片相比,烟梗的硝态氮含量和 TSNAs 积累量显著高于叶片[59]。
2.3 生理因素
从生理学的观点来看,进入植物的硝态氮必须经过硝酸还原酶的作用还原成氨,才能形成氨基酸和蛋白质,参与植物的氨代谢过程。因此,氮代谢关键酶活性对于硝酸盐积累量具有很大影响。碳氮代谢是植物活动的基础代谢,两者密切相关。氮代谢为烟株碳代谢提供氨基酸、酶蛋白和光合色素[60]。光合作用及其产物为硝酸盐还原同化作用提供碳骨架和能量动力,对植物硝酸盐积累具有重要影响[61]。在大麦和玉米作物色素缺乏突变体中,其NRA较野生型品种下降30%-40%,与其突变体的色素含量下降,光合产物减少,还原作用活动所需的能量供应和还原力降低密切相关[62]。有研究表明,白肋烟氮素还原能力比烤烟弱,是其硝酸盐大量积累的直接原因,色素含量低、光合作用和碳固定能力弱从而导致不能为其氮代谢过程提供充足的能量,是其硝酸盐大量积累的主要原因[52]。
2.4 生态因素
生态条件主要包括气象条件和土壤条件,是影响烤烟烟叶化学成分的重要影响因素。气象条件是指烟叶生长所处的气候因子,包括光照、温度、降雨量以及空气的相对湿度等;土壤条件是指植烟土壤的各种理化性质包括土壤的pH值、有机质、大中微量元素、松紧度、含水量等等,古战期等从气候和土壤等方面表述了其对烤烟烟叶化学成分的影响,对优质烤烟生产提供理论参考[63]。
光照强度通过影响光合作用调节硝酸还原酶活性,最终影响植株体内累积,光周期缩短和弱光照均会导致的累积[64]。Burton等[65]把一片烟叶分成41块,分析其和分布和关系,得出最高出现在叶片基部,朝叶尖方向降低。光作为重要的生态环境因子通过调节的吸收、基因表达和硝酸还原酶活性来调节氮代谢[66-67]。高祖民等[68]指出,在遮光处理条件下,小白菜体内硝酸盐积累量是不遮光条件下的30多倍。王德芳[69]研究表明在正常光照条件下光合作用良好,植株生长量大,吸入的硝酸盐被稀释而不致累积很多,同时光合作用可提供硝酸还原酶的能量,使之转化为铵态氮,也有利于减少硝酸盐的累积。
硝态氮的吸收、运输与水分的运动密切相关。质流是水分驱动的物质运动,蒸腾作用的持续进行,使溶解于水中的硝态氮向植物体内各处迁移,分布于不同器官的组织内部及外部空间的水分中,因此,硝态氮的代谢过程离不开水分。干旱情况下,硝酸还原酶活性受抑制,硝态氮的积累显著增加[71]。因此,合理灌水是调控烟叶硝态氮含量的重要手段。
土壤是烟叶生长的基础,土壤肥力水平与烟叶硝酸盐积累关系密切。许自成等从土壤有机质含量、pH值、主要营养元素等土壤肥力因素对烤烟叶片硝酸盐积累影响进行阐述[72]。土壤有机质含量是表征土壤供氮能力的重要指标,陈伟等[73]研究表明,不同等级烟叶硝酸盐含量与土壤有机质呈极显著正相关。
2.5 栽培因素
磷、钾肥在促进了一系列生理活动的同时,促进了无机态氮化合物合成蛋白质,从而减少了硝酸盐含量[77]。磷是烟草NR和NiR的重要组成成分,对烟草氮素吸收和同化具有重要的影响[58]。钾离子可提高植株硝酸盐酶活性,降低硝酸盐含量,与硝酸盐含量呈极显著负相关[78]。氯离子同时对硝酸根离子有拮抗作用和取代效应,可以抑制硝化细菌的活性,促进氨基酸含量增加并合成蛋白质,从而降低植株硝酸盐含量[79]。Hitoshi研究表明:缺锌时,烟草的RNA含量显著下降,使RNA的代谢受阻进而影响蛋白质的合成,造成硝酸盐的积累[80]。
土壤中Ka、Ca、Mg元素含量决定了土壤pH,土壤pH值又直接影响硝酸盐和亚硝酸盐吸收和积累,因此,它们在植物N素转化中起了很重要的作用[81]。在众多微量元素中,钼对植物硝酸盐积累的影响最为明显。钼是NR的重要组分,钨和钼是同族元素,与钼存在竞争关系,缺钼或施用钨元素会抑制NR活性,引起植株硝酸盐积累[82-83]。
目前烤烟生产中,肥料多采用条施和穴施的方式施入,肥料利用率较低[84]。张学军等[85]研究表明,不同施氮处理对土壤溶液硝态氮含量有直接影响,采取滴灌施肥技术可以在不影响产量的条件下减少施氮量,从而降低蔬菜硝酸盐含量。
种植密度是决定作物有效截光叶面积、影响群体光合效能和田间微气象的主要因素, 而留叶数则直接影响烤烟打顶后干物质的生产与分配[86],研究表明,种植密度及留叶数对烤后烟叶硝酸盐含量的影响明显[87]。种植模式对烟草产质量、养分利用和土壤理化性质有很大影响,烤烟连作显著降低烤烟根、茎和叶的生物量[88],同时造成烟地养分比例失调[89]。
打顶在促进根系发育的同时,促进对氮素的吸收,硝酸还原酶作为一种诱导酶,其活性也得到加强,因而促使硝态氮含量下降。研究表明,不同打顶时间对硝态氮含量影响不同,第一朵中心花开放时进行打顶是保证烟叶中硝酸盐含量适量的有效方式[90]。
3 降低硝酸盐积累技术
3.1 运用生物技术和遗传育种进行品种改良
筛选低硝酸盐积累的优良品种是降低硝酸盐积累的一种有效途径,根据目标,选用合适的亲本是快速且高效地培育优良品种的有效方法[91]。随着分子生物学的发展,人们逐渐开始采用生物技术进行品种改良,能达到事半功倍的效果。Lu等[92]以白肋烟为材料,通过表达NiA基因以获得高NR活性的植株,在晾制结束后发现,烟叶硝酸盐和TSNAs均有明显降低。Quillere等[93]对烟草植株过表达NR基因,发现对叶片内的含量降低的影响十分显著。GS同工酶作为氮素代谢的核心之一,是作物体内氮素同化和转移再利用的关键酶。Oliveira等[94]研究表明,GS1基因过表达烟株叶片的鲜重、干重、可溶性蛋白含量和谷氨酰胺合成酶活性都明显增加。Zheng等[95]发现过表达苎麻BnGS1-2基因的烟株对氮素的吸收和同化能力增强,并导致株高增加且鲜重和干重提升。
有报道表明,硝酸盐转运蛋白基因的过量表达可显著提高作物的氮素利用效率和产量[96-97]。朱思慧等[98]成功将吊兰的NRT 基因 CcNPF8.3.1(Protein NRT1/PTR Family 8.3)、NRT 基 因 CcNPF8.3.2、NRT 基因 CcNPF5.2和NRT 基因 CcNPF8.1转入到双缺陷型多形汉逊酵母和烟草中,NRT 基因的表达在一定程度上提高了对硝酸盐的吸收转运功能和植株的抗病性。据报道,在不影响产量的情况下,白肋烟氮素转运蛋白基因CLC-Nt2的下调使其硝酸盐含量降达60%-70%[12]。
如今,陆续有研究借助叶绿体基因工程分别将蓝细菌和藻类的CO2固定机制引入到烟草中,通过改良烟草叶绿体基质上的 RuBisco 酶来提高 CO2的固定速率[99-100]。除此之外,将拟南芥中编码果糖 1,6 - 二磷酸醛缩酶(Fructose 1,6-bisphosphate aldolase)的基因AtptAL 转入到烟草叶绿体中,其醛缩酶活性较野生型烟草提高1.4~1.9 倍,在高 CO2浓度下可达 2.2 倍,能够促进 CO2的固定,提升光合效率[101]。此项技术有利于改善白肋烟叶色突变的缺陷,从而有助于减少硝酸盐的积累。
烟草不同品种间的氮肥利用率存在很大的差异,沈善敏等[102]认为,通过品种改良可使氮肥利用率提高20%~30%,因此筛选氮高效品种或基于氮高效功能基因为靶标的分子育种应是提高氮肥利用率最有效的生物学途径[103-105]。
3.2 运用化学手段调控生理代谢
硝酸盐吸收、转运和还原同化过程复杂,易受硝化抑制剂、激素、碳水化合物等化学物质的调控[106]。液泡膜H+焦磷酸酶(V-PPase)能促进营养贮藏,而液泡膜ATP酶(V-ATPase)能抑制营养贮藏,V-PPase和V-ATPase抑制剂能促进NR活性而降低植株组织硝酸盐含量[107]。氮抑制剂能控制硝化速度,减少氮素的损失,从而提高氮肥利用率[108]。国内外的研究表明氮抑制剂对蔬菜的硝酸盐积累有明显的抑制作用,目前常用的氮抑制剂有双氰胺(DCD),与只施尿素相比, 硝酸盐含量明显下降,用量以占总施入纯氮量的 10 % ~20 %为宜[109]。叶面喷施 0.05 %浓度的钼肥水溶液和 DCD复合施用其降低硝酸盐的效果是两者的和[110]。陈翔等[13]在四川达州研究表明,白肋烟施氮量为210kg/hm2时,采用现蕾打顶结合打顶后涂抹仿生型信号分子 (Bionics Signal Molecule,BSM)的方法能够有效降低上部叶硝酸盐含量。在白肋烟打顶后,喷施生长素、萘乙酸和水杨酸等激素物质能调控烟叶硝酸盐含量[7]。在旺长期喷施钼酸钠同时在成熟期喷施钼酸钠和草丁膦的方法有利于促进烟叶发生氨气挥发现象,可有效促进烟叶氮素还原同化,使调制后烟叶硝酸盐含量下降47%,进而降低烟叶TSNA含量[111]。
碳氮代谢是植物活动的基础代谢,两者密切相关。喷施丙三醇、壳寡糖和海藻糖等外源糖类均有助于提高烟叶色素含量和光合作用,降低硝酸盐含量[112-114]。
3.3 合理肥料运筹进行营养调控
土壤有机质含量的高低会影响氮 、磷、硫和其它营养元素的供应。史宏志等[115]研究表明施氮量和基追肥比例对肥料氮的吸收利用率均有显著影响,在0~255 kg/hm2施氮范围内,以施氮量 210 kg/hm2,基肥比例为40%~60%为好。陈翔等[13]研究表明,在施氮量为210 kg/hm2时白肋烟烟叶中的硝酸盐积累量最低。在进行优化施肥时,除考虑施氮量外,氮肥的施用方式也是影响烟叶硝酸盐含量的重要因素,孙泽东等研究表面滴灌施肥技术将滴灌与施肥相结合,能够提高肥料利用率,促进烤烟氮素吸收和利用[116]。同时在一定氮素水平下,合理搭配使用不同形态氮肥,对减少烟株含量具有一定效果[102]。高秀瑞等[117]在研究中发现,采用甘氨酸替代部分硝态氮肥料的方法能有效地降低硝酸盐含量。运用水肥一体化等先进的高效施肥方式可显著提高烟株的NR活性和NITR基因的相对表达量,提高氮肥利用率[118],与减氮相结合可显著降低烟叶硝态氮积累。
氮、磷、钾比例失调,也是硝酸盐积累的一个重要因素。研究表明,在氮磷肥充足的情况下,叶面喷施钾肥能显著降低烤烟烟叶硝酸盐含量[119]。氯离子通过对硝酸根离子的拮抗作用和取代效应,抑制硝化细菌的活性,促进氨基酸含量增加并合成蛋白质,从而降低植株硝酸盐含量[79]。因此对少数缺氯土壤可适当施用含氯肥料。烟田施用硼、锌、钼微肥后,烤烟质量和品质明显改善[120]。施用钼肥后,烟叶NR活性和叶绿素含量升高,光合作用增强,硝酸盐含量下降[121]。研究表明,在旺长期喷施钼酸钠后,烟叶NR和GS活性升高,硝酸盐还原同化作用增强,对降低烟叶硝酸盐含量有利[111]。
3.4 其它农艺措施
光合作用与硝酸盐的含量密切相关,充足的光照能促进氮代谢。因此,为了减少硝酸盐的积累,需要改进光照条件,合理安排烟叶的栽植密度和留叶数 ,从而避免烟叶上下棚叶位之间的相互遮阴。Steingrover等[122]在晚上增加几小时的低光照处理,能够降低液泡中的硝酸盐含量。水分缺乏会抑制NR的合成,引起烟叶硝酸盐大量积累,而适量的灌水处理能明显降低烟叶硝酸盐含量[71]。烤烟轮作能有效改善土壤微生物环境[123],提高烟叶产量和质量,促进肥料利用率,对降低烟叶硝酸盐含量有利[124]。
嫁接作为一种栽培技术手段被广泛应用,在适宜的环境条件下,嫁接植株的根系活力明显提高,叶片数目大幅度增加,同时光合生产能力得到了提升[125-127]。嫁接玫瑰中,利用特定的砧木能刺激 NR 的活性和对氮素的吸收能力[128]。嫁接甜瓜中的氮素含量、关键酶活性、酶活基因相对表达量、转运蛋白基因(CmNRT2.4、CmNRT2.5)和控制转录因子HY5的基因相对表达量均显著高于自根甜瓜幼苗,地上部生物量显著增加,生长势和氮代谢能力增强[129]。研究表明,烤烟嫁接苗氮代谢关键酶活性和叶绿素含量均高于自根苗,对降低硝酸盐积累量有利[130]。
4 展望
硝酸盐是烟叶TSNAs形成的重要前体物,烟叶硝态氮含量增高是造成烟叶调制和贮藏过程中TSNA形成的重要因素,在明确硝酸盐积累机理的基础上,采取遗传改良和生理、农艺调控技术降低硝态氮积累,是降低烟叶TSNA含量、提高烟叶安全性的重要途径,也是烟草科研的重要方向。硝酸盐积累量与氮效率密切相关,提高氮效率,减少氮肥施用量,可有效降低烟叶硝态氮积累。因此,降低烟叶硝酸盐含量对于减氮增效,促进烟叶优质高效生态安全意义重大。
硝酸盐代谢途径主要包括硝酸盐吸收、转运和还原同化环节,易受品种、施肥、光、温度、水分、土壤pH等环境因子的影响,烟叶硝酸盐积累是复杂代谢后的综合表现。以往对于降低硝酸盐的研究多集中在耕作、施肥及土壤改良方面,这些以调控外界环境为主导的植物营养调控手段有一定的局限性。烟叶硝酸盐含量的差异多是由遗传因素控制,如何通过基因工程等生物技术手段选育低硝酸盐白肋烟品种仍是研究的重点方向。目前,已经筛选到控制硝酸盐积累的一些关键基因,但这些基因与硝酸盐积累的关系需进一步验证,对控制硝酸盐积累的关键基因需进一步挖掘。加上氮代谢与碳代谢的偶联关系,对于碳代谢过程中关键酶基因及转运蛋白对硝酸盐积累的作用尚有待探究。
烟草是产量、品质严重依赖硝态氮的作物,烟叶硝态氮含量受氮肥种类及用量、土壤供肥特性、品种对氮素利用效率和利用模式等因素的综合影响,因此必须注重多种途径和技术的结合,比如提高氮素还原同化能力与增加光合碳固定转化能力相结合,提高氮素利用率与氮肥减量和调整形态相结合,减少有害物质形成与优质烟叶生产相结合等,有效促进优质低害烟叶生产。
