白云-荔湾深水区新生界生物礁识别与发育模式及主控因素探讨
2019-05-16周小康万琼华
梁 杰, 曾 驿, 周小康, 罗 泽, 万琼华, 卫 哲
(中海石油(中国)有限公司 深圳分公司研究院,深圳 518054)
0 引言
生物礁具有良好的储集性能,世界上主要生物礁大油田的总储量已达40×108t以上[1]。生物礁是由造礁生物原地生长堆积形成的碳酸盐岩建隆[1-2],其生长发育受到造礁生物演化、光照及海水温度、水深等环境条件及古地理和古构造背景等因素的控制[3-5],因此生物礁的分类存在多种依据。按照造礁生物种类可以将生物礁细分为珊瑚礁、珊瑚藻礁、海绵礁等;按照礁灰岩组分可细分为基质支撑的生物礁、骨架礁及胶结礁[1-2];按古地理位置及礁体形态可以细分为岸礁、堡礁、点礁、塔礁及环礁等[1,6]。基于造礁生物种群、礁灰岩组分的分类需掌握大量的露头、岩心及薄片资料,往往用于高成熟勘探区的储层分析等研究,而古地理格局及形态分类相对容易展开,适用于礁体宏观分布的研究[1]。

图1 研究区范围Fig.1 The structure sketch of the research area
生物礁油气勘探在珠江口盆地(东部)油气勘探中占有重要的地位。目前,在珠江口盆地(东部)东沙隆起浅水区陆续发现HZ33-1、LH11-1、LH4-1、LF22-1等生物礁油藏[1,9-11],其中HZ33-1是我国海上发现的第一生物礁油藏[1,11],LH11-1是我国海上发现的最大规模的生物礁油藏[9,10-11],这一系列生物礁油藏的发现展现了珠江口盆地(东部)生物礁油气藏勘探的巨大潜力。而相对于东沙隆起浅水区生物礁油藏巨大的勘探前景,珠江盆地(东部)深水区生物礁油藏的勘探却止步不前,鲜有人对该区生物礁识别特征、发育模式及主控因素进行过系统研究。笔者通过地震资料的精细解释,结合钻井与前人研究资料,对白云-荔湾深水区生物礁进行了识别追踪,并综合生物礁发育的古构造背景对生物礁进行了分类,系统总结了该区不同类型生物礁的地震响应特征,发育模式,同时结合研究区古地理环境的变化对研究区生物礁发育的主控因素进行了系统分析,希望能为该区及类似地区生物礁油气勘探有所指导。
1 研究区概况
研究区(图1)位于南海北部大陆边缘深水、超深水区(水深为300 m~5 000 m),北部向东沙隆起和番禺低隆起过渡,南部向双峰盆地过渡,主要包括白云凹陷、云开低凸起、顺鹤隆起-云荔低隆起及其以南的超深水区域。与珠一地区诸多凹陷相似,研究区自下而上发育始新统半地堑河湖相沉积的文昌组、下渐新统河湖相煤系沉积的恩平组、上渐新统浅海陆架三角洲-深水陆坡沉积的珠海组、下中新统深水陆坡-深水扇沉积的珠江组-韩江组和上中新统深水陆坡欠补偿沉积的粤海组、上新统万山组及第四系[13-14]。截止目前,研究区钻遇碳酸盐岩井位共计16口,在恩平组、珠海组、珠江组和韩江组均有钻遇碳酸盐岩,厚度变化为0.51 m~317 m。前人对研究区碳酸盐岩识别特征、发育模式及主控因素尚未进行过系统总结。基于此,本文在类比东沙隆起浅水区碳酸盐岩地震识别特征、发育模式及主控因素[15-19]的基础上,对研究区的生物礁识别特征、发育模式及主控因素进行了系统阐述,并分析研究区生物礁发育层段的规律性,指明了研究区碳酸盐岩油气勘探的重点和下一步勘探方向,希望能为该区碳酸盐岩油气勘探指明方向。
2 生物礁的地震特征识别
国内、外学者从生物礁外形、顶界、底界、内部反射特征等方面对生物礁的地震识别进行过系统研究[9,14,19],该研究成果对本次研究具有重要指导意义。笔者基于前人研究成果从顶界、底界、内部反射特征及与围岩的接触过渡关系等方面,总结了白云-荔湾深水区碳酸盐岩在地震剖面上的识别特征。
1)顶界反射特征。由于碳酸盐岩的密度和速度均比上覆碎屑岩大,波阻抗差异明显,碳酸盐岩顶界面呈现出明显的强反射(正相位、波峰显示)、连续性好的地震反射特征。
2)底界反射特征。当碳酸盐岩下伏为海陆过渡相碎屑岩沉积,波阻抗差大,底界表现为较高连续性的反射(负相位,波谷显示)。
3)内部反射特征。碳酸盐岩台地内部整体表现为良好的成层性,中-弱振幅,中-低连续性,中频,偶见弱反射、杂乱反射-空白反射。
4)与围岩的接触过渡关系。 碳酸盐岩与碎屑岩主要有2种接触关系:①出现在碳酸盐岩与碎屑岩连续过渡区,表现为明显的碳酸盐岩强反射向碎屑岩中-弱振幅的过渡特征;②出现在碳酸盐岩建隆向碎屑岩过渡区,由于碳酸盐岩生长速度快于围岩的沉积速度,其生长要求明显的古隆起背景,因而后期的补偿沉积在碳酸盐岩的外围形成了明显的超覆现象。
为了减少该区生物礁的误判风险,我们在识别碳酸盐岩的基础上,对研究区的生物礁进行了甄别,并总结了生物礁与火山丘的识别差异,具体如下:
1)生物礁外形反射特征。生物礁是造礁生物原地生长且生长速度快于围岩沉积速度的大型碳酸盐岩建隆,具有坚固抗浪骨架,因此,在地震剖面上表现为明显的隆起加厚特征,总体形态以丘形、透镜体状、箱型为主。
2)生物礁顶界反射特征。由于礁灰岩的密度和速度均比上覆碎屑岩大,波阻抗差异明显,碳酸盐岩顶界面呈现出明显的强反射(正相位、波峰显示)、连续性好的地震反射特征,同时与上覆围岩多表现为不整合接触关系,即上覆围岩多呈超覆接触关系沉积在生物礁上部。
3)生物礁底界反射特征。生物礁底界反射相对较弱,表现为弱连续、弱振幅特征,且由于密度和速度均比下伏碎屑岩大,因此表现为负相位特征。
4)生物礁内部反射特征。生物礁内部具有平行、进积或退积地震反射特征,也可见杂乱、空白弱反射。
5)与火山丘的区别。火山丘内部为杂乱反射特征,底部不连续,可见明显火山通道,火山通道多呈塔状。
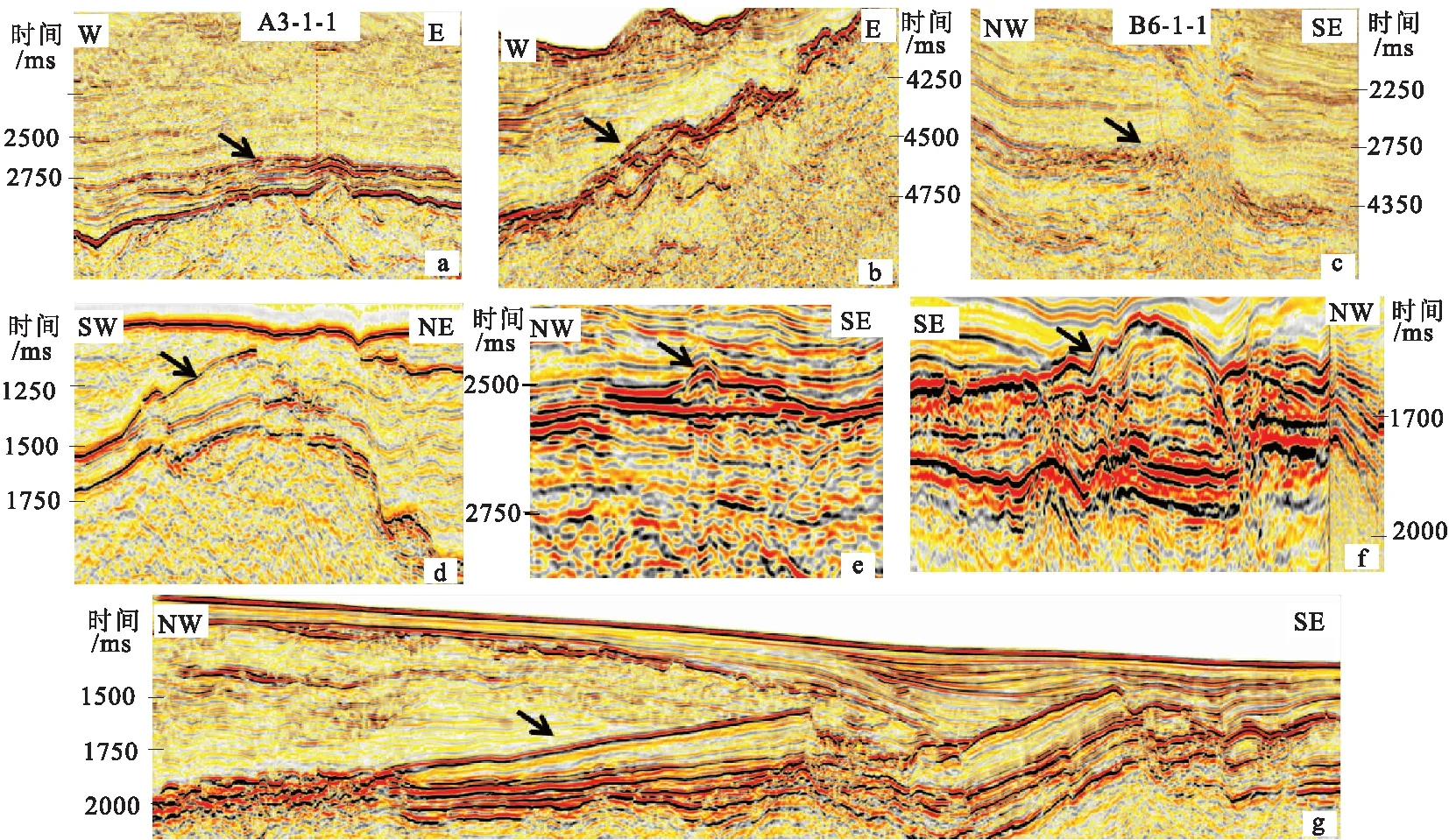
图2 碳酸盐岩沉积类型Fig.2 The developed types of carbonate in research area(a)薄层灰岩;(b)古隆起斜坡礁;(c)火山周缘礁;(d)构造后期抬升形成的构造高部位礁;(e)孤立塔礁;(f)古隆起顶部礁;(g)台地边缘礁
3 生物礁的发育模式
3.1 碳酸盐岩和生物礁的类型及识别
根据碳酸盐岩及生物礁发育的位置及发育的规律性,将研究区碳酸盐岩及生物礁划分为7种生长类型(图2),不同类型碳酸盐岩及生物礁地震反射特征及发育背景具差异性:
1)薄层灰岩。研究区已有多口井钻遇,地震相均表现为连续、强反射特征;厚度变化范围大(0.51 m~317 m),平面分布范围广;纵向上文昌组、恩平组、珠海组、珠江组和韩江组均有发育,具有“横向分布面积广,纵向分布层段多”的特征。
2)古隆起斜坡礁。该类生物礁主要分布在古隆起缓坡带,该类生物礁主要发育在顺鹤隆起斜坡带上,表现为沿着古隆起缓坡带呈追补生长特征,可细划分为4个丘状或低丘状或透镜体礁形(直径分布范围100 m~200 m,面积大于50 km2),代表四期生物礁生长期次,对于每一个透镜状礁体,可以看见清晰的同向轴近乎平行于缓坡带的地震反射特征,顶强-中反射,底部连续可追踪,内部可见平行反射特征,各个礁体与上覆地层表现为超覆不整合接触,反应了海平面的变化对生物礁体生长的影响。
3)火山周缘礁。Bb-1-1井钻遇的碳酸盐岩建隆为典型的火山周缘礁,发育在珠海组顶部及珠江组内部,礁体分布范围限于火山岛周缘,面积270 km2,厚度均大于200 m。剖面上两翼坡度很大,灰岩尖灭部位同相轴明显减弱,中部同相轴下凹部位为明显的火山口,火山岩浆再次活动向上刺穿整个珠江组,靠近火山口部位受岩浆影响严重,内部出现空白反射。
4)构造后期抬升形成的构造高部位礁。该类生物礁以发育在东沙25构造带的构造高部位礁为例,分布面积230 km2,厚度大于600 m,剖面上被晚期断层活动切断,上覆碎屑岩超覆沉积在礁体之上,顶界面中振幅、连续可追踪,底界与下覆碎屑岩不整合接触,表现为中振幅、连续可追踪,礁体内部弱反射,可将礁体划分为A、B两个生长期次。A期,生物礁响应特征为平行弱振幅、低连续、弱成层性,主体以向上建造为主,见加积、双向下超反射结构,反应生物礁生长速率大于海平面上升速率,为海侵期生物礁发育的典型地震响应特征;B期,生物礁响应特征为中弱振幅、弱连续、中频的反射,成层性较好,见强制海退的进积反射特征,但主要为向南单侧进积生长,底部见下超反射。
5)孤立塔礁。该类生物礁发育在台地边缘礁向海一侧,规模大小不等,具丘状外形,顶、底强-中反射,连续可追踪,内部可见平行、杂乱或空白地震反射特征,与上覆地层表现为超覆不整合接触。

图3 典型生物礁发育模式示意图Fig.3 Diagrams of different types of carbonate platform mode(a)台地边缘礁发育模式;(b)古隆起斜坡礁发育模式;(c)古隆起顶部礁发育模式;(d)与构造后期抬升相关的构造高部位礁;(e)与古火山相关的火山周缘礁
6)古隆起顶部礁。该类生物礁分布在云荔低凸起东南部的古隆起基底之上,灰岩发育层位主要为珠江组,规模不等。该种模式生物礁发育主要分为两个重要阶段:①礁体生长阶段 当礁体生长速率接近或等于海平面上升速率时,生物礁呈加积生长,同时伴随着海平面的短暂下降,在礁体边缘也可见进积地震相特征;②礁体淹没阶段 海平面上升速率大于生物礁生长速率时,生物礁淹没,晚期碎屑岩超覆沉积在礁体之上。
7)台地边缘礁。研究区发育的镶边碳酸盐岩台地主要分布在东沙隆起向白云凹陷的自然延伸区域,最为典型特征是台地规模大(单个台地边缘礁厚度100 m~700 m,面积70 km2~250 km2),台缘礁及台缘之下的混积陆棚厚度变化明显。其顶、底表现为强振幅、连续反射特征,内部表现为近平行地震反射特征,而台缘之下的混积陆棚主要发育在珠江组顶部,厚度明显减薄,反射轴比较杂乱,且分布范围局限,仅在隆起区分布,为海侵初期隆起边缘的镶边碳酸盐岩台地[4,6]。
3.2 生物礁的发育模式
生物礁的发育模式存在多种划分依据,如按造礁生物种类[1-2]、礁体形态[1,6],但是以上划分依据都需要掌握大量的露头、岩心及薄片资料,往往用于高成熟勘探区的储层分析等研究,对于生物礁低成熟勘探区,国外学者主要基于生物礁生长类型和发育构造背景进行划分[17],因此本文基于研究区生物礁的生长类型,考虑构造活动和古隆起背景及生物礁的分布位置,建立了生物礁的3种发育模式。
3.2.1 台地边缘礁发育模式
台地边缘礁(图3(a))主要发育在具有高能外部边缘的浅水台地边缘上。台地边缘与深水盆地之间有坡度较陡的斜坡,水体浅、能量高,是发育高能障壁礁或滩带的有利场所,其向岸一侧可发育点礁及低能的泻湖,向海一侧可能发育塔礁。本区发育的台地边缘礁主要分布在东沙隆起向白云凹陷的自然延伸区域,最为典型特征是分布面积大,台缘礁及台缘之下的混积陆棚厚度变化明显。该种模式生物礁仅在白云北坡近东沙隆起区分布[5-10],且主要发育在珠江组下段,厚度相对较大,为海侵初期台地边缘发育的生物礁。
3.2.2 古隆起斜坡礁发育模式
古隆起斜坡(图3(b))要求海底向海平缓倾斜,水体逐渐变深,从近岸高能波浪作用带向下逐渐过渡为深水低能环境,其间没有明显的坡折。该种模式下生物礁发育主要受海平面的变化:早期低位域时期,生物礁在缓坡带底部发育,之后海平面上升,生物礁表现为沿着缓坡追补生长的特点,当海平面上升速度大于生物礁生长速度时,生物礁逐渐被海水淹没并停止生长。
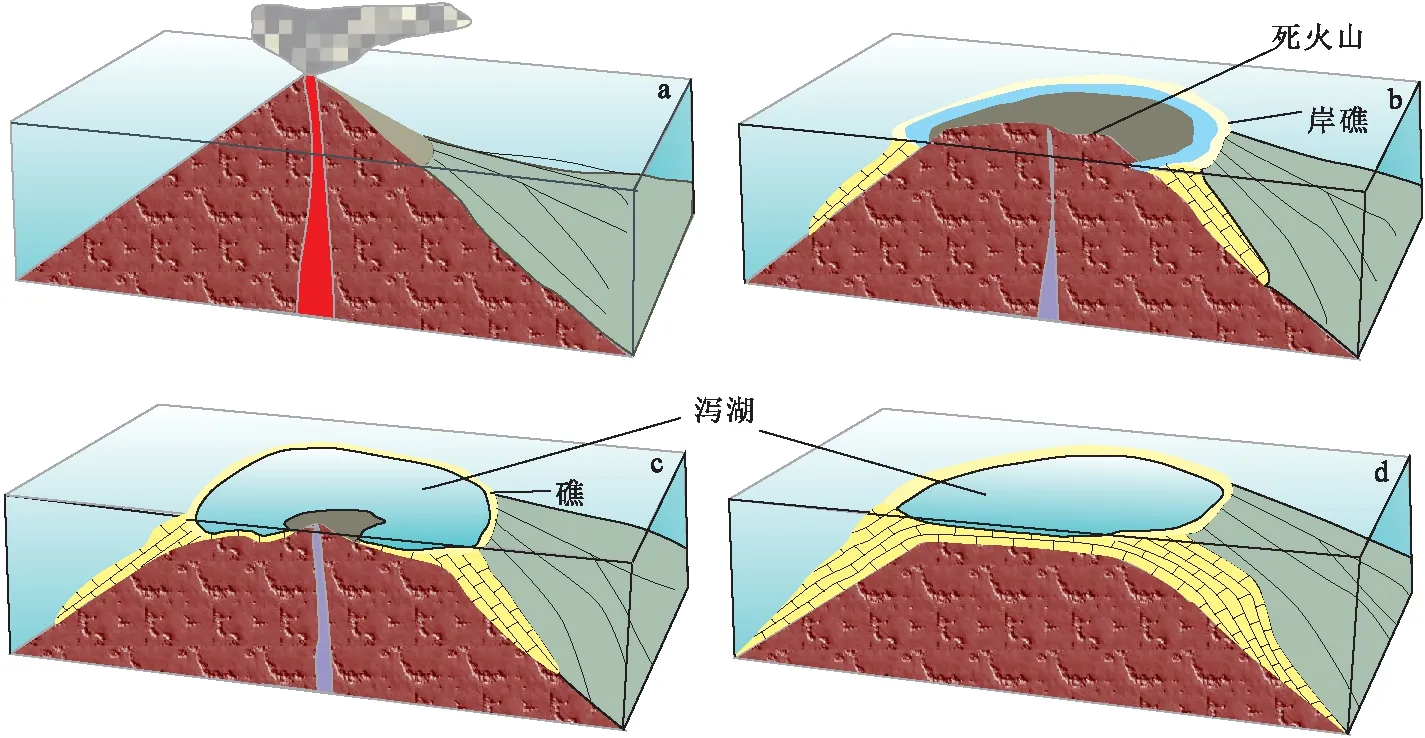
图4 火山周缘礁发育模式Fig.4 Model for isolated carbonate platform associated with volcanism(a)火山喷发;(b)岸礁发育;(c)环礁发育;(d)珊瑚鸟形成

图5 研究区碳酸盐岩与生物礁分布图Fig.5 The structure sketch of Carbonate and reefs in the study area
3.2.3 古隆起顶部礁发育模式
古隆起顶部礁具有面积相对较小、厚度相对较大(多于100 m)的特点,白云-荔湾深水区古隆起顶部礁多发育在小面积的孤立的古隆起之上。根据构造活动及古隆起的形成机制,可将古隆起顶部划分为①与基底古隆起相关的古隆起顶部礁(图3(c));②与构造后期抬升相关的构造高部位礁(图3(d));③与古火山相关的火山周缘礁(图3(e))。
古隆起顶部礁和构造高部位礁均发育在古隆起内地势相对平缓的位置,为台地内部点礁或台地边缘堡礁,该类生物礁外形呈丘形,其顶界为平行-亚平行强反射轴,上覆海侵层披覆于其上,底界受基底隆起影响,为中-弱振幅的强反射轴,内部杂乱-前积-空白反射结构,可以隐约见到生物礁的垂向建造(图3(d), 图3(f)),而礁体范围不断变小,反应海平面上升,礁体生长至死亡的过程[19-20]。
火山周缘礁的发育与火山活动密切相关,分布范围限于火山岛周缘(如图2(c)),通常该发育模式经历5个过程[19-20]:①受岩浆上涌影响,火山岛形成,形成可供生物礁发育的隆起背景;②相对海平面上涨,构造活动减弱,在火山岛两翼具备生物礁发育的清水条件,岸礁发育,开始形成碳酸盐岩建隆;③相对海平面继续上升,火山口几乎淹没于海平面之下,生物礁在之前形成的碳酸盐岩建隆之上持续发育,形成环礁,内部出现局限潟湖沉积;④相对海平面继续上升,火山口被全部淹没,环礁持续向上建造,其内部潟湖环境水体能量较低,且由于生物礁生长,水体中碳酸钙过饱和,潟湖内部碳酸盐岩沉淀,火山岛消失,珊瑚岛形成;⑤当海水上涨不再适合生物礁体生长时。整个礁体淹没,上覆碎屑沉积直接超覆在礁体之上,生物礁停止发育(图4)。
4 分布及其发育主控因素分析
4.1 碳酸盐岩与生物礁的分布
本次研究在统计白云-荔湾深水区16口钻井灰岩厚度统计及高密度二维、三维地震剖面识别的基础上,理清了白云-荔湾深水区碳酸盐岩及生物礁分布特征。总体而言,白云-荔湾深水区碳酸盐岩主要分布于南部隆起带顺鹤隆起-云开低凸起、兴荔凸起-云荔低凸起及东沙隆起向白云凹陷延伸带之上,发育层段位于T70~MFS18.5之间。生物礁发育时期有一定规律性,鹤山凹陷及番禺低隆起生物礁主要发育在恩平组沉积时期,顺鹤-云荔低隆起带生物礁发育在T60层位上下即23 Ma左右,白云凹陷及流花-东沙隆起带生物礁主要发育在珠江组沉积时期。
研究区薄层碳酸盐岩广泛分布,分布面积约20 000 km2,其中流花-东沙隆起带碳酸盐岩分布面积约2 000 km2,番禺低隆起碳酸盐岩分布面积约600 km2,顺鹤隆起-荔湾凹陷-兴荔凸起碳酸盐岩分布面积约13 000 km2,白云坳陷-云荔低凸起碳酸盐岩分布面积约2 700 km2。灰岩最大厚度分布于东沙-流花隆起带,厚度多超过300 m,顺鹤-云荔低隆起薄层灰岩(5 m~10 m)广泛分布,由于缺乏钻井资料,确切厚度有待证实。大型生物礁多呈孤立、零星的分布,本次识别出可靠生物礁37个(图5),其中台地边缘礁共7个,孤立塔礁共8个,基底古隆起顶部礁共10个,古隆起斜坡礁共2个,与构造后期抬升相关的构造高部位礁共3个,火山周缘礁共7个。
4.2 碳酸盐岩发育主控因素
4.2.1 构造演化与古地理条件
生物礁发育背景与盆地演化背景密切相关[15-19],笔者对白云-荔湾深水区生物礁发育在时间和空间上的规律性与该区构造演化及沉积充填背景进行了详细分析。
研究区新生代沉积充填从老到新经历了断陷期文昌组的半地堑河湖相沉积、断拗期恩平组的河湖相煤系沉积、过渡期珠海组的浅海陆架三角洲,拗陷期珠江组-韩江组的深水陆坡-深水扇沉积和深水陆坡欠补偿沉积的粤海组、上新统万山组及第四系[6](图6)。
断陷期文昌组河湖沉积充填了规模巨大的文昌组陆相断陷湖盆沉积,不利于碳酸盐岩及生物礁发育。断拗期恩平组大型湖盆沉积充填开始于晚始新世,研究区西北面的番禺低隆起及凹陷主体为湖相沉积环境,而白云凹陷南部开始与海相通,开始呈现典型的海相特征。已钻井在恩平组晚期地层中发现栎类花粉-柯式双沟粉-双束松粉组合中常绿栎类花粉多占明显优势,反应恩平组晚期为较暖湿气候,综合分析恩平组晚期在海水清-浅-温暖的条件下是利于生物礁生长和发育的。
过渡期浅海陆架三角洲发育在珠海组时期,在珠海组沉积初期白云凹陷的北坡、西南云开低隆起带及南部隆起带与东沙隆起带都暴露在地表之外,只有南部隆起带中间部分区域与外海相通,此时的深水区还是局限海湾的沉积环境;在珠海组沉积中期,白云深水区的沉积和沉降中心从NE向转为NNE向,盆地发育另外一次重要的构造沉降的迁移,随着海侵规模逐渐扩大,研究区主体处于滨浅海环境,同时,珠海组陆架坡折带雏形开始形成。此后,珠海组沉积晚期陆架坡折带稳定发育在南部隆起带附近,受古珠江三角洲供源主体沉积作用、稳定的构造沉降和大的海退背景控制,珠海组晚期沉积时的陆架坡折带由NW向SE方向迁移,直至23.8 Ma左右,珠海组开始最大珠海组开始最大一次海退进积[6]。始新世末至渐新世早期全球的气候变冷事件[21-24]以及现有文献资料[24-25]对于珠江口盆地珠海组的孢粉学特征是亚热带、温带孢粉植物群的表述,反应珠海组沉积时期为凉干气候,综合分析认为南海北部深水区在珠海组时期,在稳定的构造沉降和大的海退背景控制下,不利于大型碳酸盐岩建隆的发育,这与研究区在珠海组并没有发现大型生物礁具有很好的吻合性。
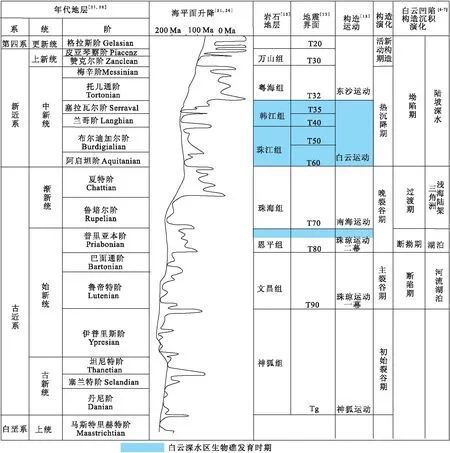
图6 研究区构造演化与全球海平面变化的关系Fig.6 The relationship between tectonic evolution and global sea level change in the study area
拗陷期珠江组-韩江组陆坡深水沉积始于23.8 Ma的白云运动,此时,珠海组陆架坡折带由南部隆起带向白云凹陷北坡珠江组陆架坡折带迁移[2],并发育了一系列火山及局部隆起带,这些火山及古隆起正是白云深水区珠江组深水环境大型孤立碳酸盐岩建隆发育的基础。伴随着大规模的海侵,在东沙隆起带发育大面积厚度较大的碳酸盐岩镶边台地,台地内部或边缘发育大型生物礁。
粤海-第四系为陆坡深水欠补偿期沉积[6],研究区的沉积速率大大降低,只有少量的细粒或泥质沉积物进入到白云-荔湾深水区,沉积以泥质为主,含少量细砂,不利于碳酸盐岩发育。
4.2.2 生物礁发育主控因素分析
国内、外学者研究表明[10-12,17]海相碳酸盐台地及其生物礁发育应具备2个条件:①清水环境;②浅水环境,但不同地区影响水体变化的因素各具差异,研究区古构造演化及古地理条件具有其独特特征,因此碳酸盐岩发育的主要控制因素也具独特性,总结起来主要表现为构造运动和海平面升降变化综合作用影响碳酸盐岩发育(图6)。
首先,构造运动对白云深水区碳酸盐岩及生物礁发育的控制表现为3个方面:
1)研究区古新世-始新世神狐组-恩平组沉积期(新南海扩张前)主要为碎屑岩沉积,此时碳酸盐岩的发育非常局限;渐新世晚期23.8 Ma附近-中新世珠江组沉积期(新南海扩张期)为适宜碳酸盐岩发育的沉积期,中新世中期韩江组沉积期至今(新南海扩张后)主要为淹没碳酸盐岩台地的深水陆坡泥质沉积[6,13-15]。因此,新南海扩展及其开启之后大规模的海侵,为研究区区海相沉积以及碳酸盐岩及生物礁发育提供了必要的构造背景。
2)研究区碳酸盐岩及生物礁分布明显受远离物源分布的古隆起或火山建隆控制,而古隆起及火山建隆的分布受构造作用控制。
3)白云凹陷持续沉降,特别是南海北部陆架坡折带在23.8 Ma~21 Ma期间[6-7],由南部隆起带向白云凹陷北坡迁移,白云深水区开始由浅海陆架沉积向深水陆坡沉积演变,是研究区碳酸盐岩及生物礁短暂发育并淹没消亡的主要构造因素。
其次, 海平面升降的影响:全球海平面变化与构造运动的叠加效应引起相对海平面周期性升降,是碳酸盐岩台地发育、演化与消亡的最直接的影响因素[10,12,16]。不同体系域生物礁发育模式明显不同。海侵体系域形成于海平面主体上升时期,古隆起斜坡礁即发育在此种沉积背景。伴随着每一次大规模的海侵作用,碳酸盐台地大范围的消减,相对高地貌的古隆起顶部礁得以保存和继续发育。高位体系域形成于海平面主体下降时期,主要发育高能动荡台地边缘礁。
5 结论与建议
笔者基于地震资料首次总结了白云-荔湾深水区的生物礁类型及识别特征,同时根据生物礁发育构造背景建立了研究区可能存在的生物礁发育模式,这也是首次在珠江口盆地(东部)生物礁低勘探程度区从生物礁发育背景出发建立生物礁发育模式;研究区处于水深超过300 m的区域,其构造演化和古地理背景的独特性决定了该区生物礁具有在构造高部位零星发育的特点,我们从构造演化和古地理环境出发首次分析了珠江口盆地(东部)深水沉积背景下碳酸盐岩及生物礁发育的主控因素,期望能为该区碳酸盐岩油气勘探提供指导作用。
白云-荔湾深水区目前油气勘探主要集中在白云凹陷及周边珠江组碎屑岩储层中[5,7],鉴于碳酸盐岩储层非均质性较强,储层精细刻画难度大等因素,要实现此类油藏的油气勘探难度较大,因此建议在油气优势汇聚区(具体包括白云坳陷内部、云荔低隆起和东沙隆起区)对已经识别出的碳酸盐岩建隆或生物礁开展储层精细研究,寻找有利碳酸盐岩储层发育区,进而推动有利目标上钻。
