微生物β-甘露聚糖酶的研究进展
2019-05-09张建新宋宜乐冯军厂常绪路
张建新,宋宜乐,冯军厂,常绪路*
(河南师范大学 水产学院,河南 新乡 453007)
β-甘露聚糖酶在动物生产、饲料、食品、医药、石油开采及生物技术等方面广泛应用[1],有关β-甘露聚糖酶的最初报道可以追溯至20世纪初到20世纪50年代[2],1960年研究学者DOETSEH等论述了能产β-甘露聚糖酶的细菌,之后到1965年,SHIBATA阐述了β-甘露聚糖酶的界定,为日后有关内容的研讨提供了理论依据[3-5]。70年代后,学者又进行了大量产β-甘露聚糖酶菌种的挑选、培育诱导,考察培养基对菌种产酶的影响。根据多年来大致的统计已有100余种能产此酶的微生物,探究最多的有细菌属的芽孢杆菌、枯草杆菌;真菌属的木霉菌、放线菌等。后来,人们进一步对不同来源的β-甘露聚糖酶进行纯化处理,对该酶的基础特性有了新的认识。
β-甘露聚糖酶不但是内切水解酶还是胞外诱导酶,其能被水解底物诱导,从而催化甘露多糖、异甘露多糖降解[1-4];还可将这些糖类中的β-1,4-甘露糖苷键降解,分解成甘露寡糖。除此之外,在含有纤维素成分原料的分解过程中β-甘露聚糖酶也有应用,已经在生物制胶、食物增甜剂、织物、水产动物日粮、医学药物、工具酶、石油开采等行业中有着相当广泛的应用[5]。
1 甘露聚糖
甘露聚糖是半纤维素(hemicellulose)中含量仅次于木聚糖的第二大复合物[6],它以β-1,4-糖苷键链接构成主链的基本骨架,由半乳糖、葡萄糖等以α/β-1,6-糖苷键链接成支链[7-9]。根据其构造分为线性甘露聚糖(linear mannan)、葡甘露聚糖(glucomannan)、半乳甘露聚糖(galactomannan)和半乳葡萄甘露聚糖(galactoglucomannan)[10]。它们在蔬菜、水果、豆制品、乳制品以及海鲜等食品的保鲜和防腐方面有很广泛用途。
甘露聚糖的水解产物是甘露寡糖,具有良好的生理调节功能,能有效降低人体胆固醇水平,减少便秘,降低血糖等,还可以利用肠内微生物调节肠道菌群环境。具有增强有益菌抑制有害菌的生长和代谢的作用,如促进双歧杆菌的生长,因此是一种很好的食品添加剂。
2 β-甘露聚糖酶的来源
甘露聚糖在自然条件下水解成甘露寡糖主要是β-甘露聚糖酶的作用。β-甘露聚糖酶在大自然中的来源非常丰富,根据不同来源可分为植物来源(如刺槐、豆类及椰果等)、动物来源(如海洋软体动物、蜗牛等低等动物)以及微生物来源等,其中微生物来源方面产β-甘露聚糖酶的报道最为常见[11-12]。在植物中,β-甘露聚糖酶常见于软木(如稻草)和发芽的植物种子(如咖啡、胡萝卜、西红柿等),果实(如番茄、椰子等)和贮藏器官(如莴苣等)。动物来源的β-甘露聚糖酶种类比植物少,并且主要由海洋软体动物(例如贻贝,皱纹盘鲍等)产生。β-甘露聚糖酶的最大来源是微生物,如细菌中的假单胞菌(Pseudomonas),枯草芽孢杆菌(Bacillus subtilis)和地衣芽孢杆菌(Bacillus licheniformis);真菌中的青霉属(Penicilliumsp.)和曲霉属(Aspergillussp.)。
此外,微生物来源的β-甘露聚糖酶不仅因为微生物资源的广泛,低成本,便利的培养条件和简单的操作提取,并且其活性高,通过在极端环境中筛选可以得到温度耐受性好的较大选择范围的酶源,较动植物来源的β-甘露聚糖酶具有更广的适用范围,目前在生产生活上得到了很大的推广与应用。
3 β-甘露聚糖酶的分子结构
1991年,Henrissat根据氨基酸序列同源性和疏水性氨基酸簇的分类将β-甘露聚糖酶分为糖苷水解酶家族5(GH5)和糖苷水解酶家族26(GH26)[13]。2008年ZHANGYL等[14]将其所研究酸热脂环酸杆菌(Alicyclobacillus acidocaldarius)Tc-12-31来源的胞内β-甘露聚糖酶划分为糖苷水解酶家族113(Glycosylhydrolasefamily113,GH113),这是β-甘露聚糖酶第一次被归入一个新的家族,编码320个氨基酸。2015年,一个新的糖苷水解酶家族134被鉴定,它起源于链霉菌、黑曲霉和米曲霉[15-17]。与过去五年报道的β-甘露聚糖相比,目前仅发现18~19 kDa的最小蛋白分子质量甘露聚糖酶,一般酶的蛋白分子质量为33~55kDa。2012年,ASPEBORGH等[18]将GH5家系划分为51个亚家族。GH5家族的甘露聚糖酶可分为GH5_7、GH5_8、GH5_10、GH5_17和GH5_36亚家族。这五个亚家族中β-甘露聚糖酶的来源既有真核生物又有细菌,其成员序列有很大差别,相似性较低(<20%)[19],属于GH26的β-甘露聚糖酶主要来源于细菌。GH5中的β-甘露聚糖大部分包含两个模块,独立的催化区和非催化区。其中,非催化结构域主要是碳水化合物结合结构域(carbohydrate binding element,CBM),与纤维素具有较好的特异性结合。研究表明,β-甘露聚糖酶的模块结构与甘露聚糖骨架的识别及其侧链的调节机制有关,但它们通常存在于酶的不同位置,具有CBM的β-甘露聚糖酶可显著提高甘露聚糖与纤维素复合物等不溶性物质的水解速率[20]。然而,也有一些酶,如GH26中的β-甘露聚糖酶,它们只具有催化结构域。
目前,已经鉴定出十多种β-甘露聚糖酶的三维结构,如低等动物中的贻贝、植物中的西红柿[21]和真菌中的里氏木霉(Trichodermareesei)。根据获得的三维结构,(β/α)8-TIM桶状结构(见图1)是GH5、GH26、GH113β-甘露聚糖酶共同具有的催化域结构,由8个α螺旋和8个β折叠交替组成[22]。GH134家族β-甘露聚糖酶的三维结构与上述三个家族不同,其结构类似于卵清溶菌酶结构。
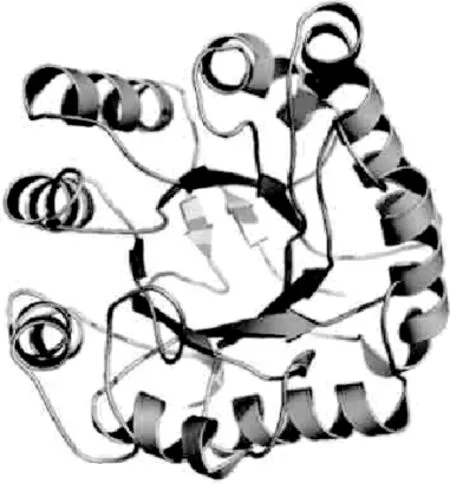
图1 (β/α)8-TIM桶状结构Fig.1 Barrel structure of(β/α)8-TIM
催化域结构的分子表面具有一个裂缝状的凹槽,其内部结构具有催化活性。有8个空间位置和功能方面十分保守的位点,分别是Arg、His、Asn、Glu、His、Tyr、Glu、Trp,对酶催化活性的作用特别重要,β-甘露聚糖酶的活性口袋就是由它们共同构成的[23]。两个Glu残基在所有β-甘露聚糖酶分子中是绝对保守的催化基团并且在催化过程中充当活性基团,其分别是起酸碱催化和亲核催化的作用。在活性位点周围还存在与底物结合的其他保守氨基酸。
4 β-甘露聚糖酶的催化机制
β-甘露聚糖酶是内切糖苷水解酶,其水解甘露糖骨架的β-1,4-D-甘露糖苷键。作用方式和在底物水解中获得的产物通常随酶和底物的来源而变化。例如,底物中葡萄糖和α-半乳糖的位置和含量以及酯酰化程度将影响水解模式和水解程度。结晶、不溶的甘露聚糖不易降解,说明底物自身的物理状态对此作用也会有影响。另外,不同的β-甘露聚糖酶对待水解的底物具有不同的要求。例如,源自瓜尔豆胶种子的β-甘露聚糖酶在底物中需要最少量的连续甘露糖残基为5个,酶的结合位点主要在甘露聚糖主链的甘露糖基上的第六位自由羟基而来源于黑曲霉的则要求底物中连续残基的个数最少为4[24]。甘露聚糖主要是杂多糖,它们的降解产物随着糖链和结构的复杂性而变化。通过β-甘露聚糖酶降解甘露聚糖后,分析该产物主要是聚合度为2~10的寡糖,产生很少或不产生单糖,酶和底物的来源对产物的聚合度有影响。研究表明,一些β-甘露聚糖酶也可以进行转糖苷基反应,这可能是β-甘露聚糖酶水解甘露聚糖获得的大多数产品是低聚糖,没有单糖或少量单糖的原因。如GUBITZ G M等[25]以Sclerotium rolfsii来源的β-甘露聚糖酶水解甘露聚糖,产物中主要包括二糖和三糖等低聚糖,当β-甘露聚糖酶作用于甘露糖四糖和甘露糖单糖(C14标记)时,发现甘露糖单糖与甘露寡糖结合;如果底物是甘露糖四糖和半乳糖单糖(C14标记),则产物主要是α-1,6-D半乳糖-β-甘露糖三糖。
β-甘露聚糖酶的催化机理与其他糖苷水解酶相似,基本上属于双取代反应机理[26]。这两个谷氨酸残基在催化反应中起重要作用,它们分别是酸碱催化基团和亲核催化基团。谷氨酸的α-羧基作为亲核催化基团,攻击底物中的异构碳原子以形成共价中间体以裂解糖苷键;另一个谷氨酸作为酸碱催化基团,首先α-羧基提供酸催化以提供质子稳定化,然后共价中间体提供碱催化,使水分子去质子化,最终完成水解过程,具体过程见图2[27-28]。

图2 β-糖苷水解酶双取代催化机制Fig.2 Disubstituted catalytic mechanism of β-glycoside hydrolase
GH134家族的β-甘露聚糖酶遵循立体翻转机制(www.cazypedia.org)[27]。质子化的Glu45作为广义酸起作用,为过渡态底物提供质子,使Asp57去质子化,作为产生去质子化的通用碱基,去质子化的水充当亲核攻击底物。与双取代机制相比,立体转换机制一步完成水解过程并改变异构体的构型,具体催化机制见图3[28-30]。

图3 β-糖苷水解酶翻转催化机制Fig.3 Inverting catalytic mechanism of β-glycoside hydrolase
5 β-甘露聚糖酶的应用
由于β-甘露聚糖酶对底物种类的作用特别广泛,所以β-甘露聚糖酶用途广泛。
在生物技术方面,在自然界许多多糖结构复杂,而β-甘露聚糖酶可以作用于β-甘露聚糖的糖苷键,然后根据酶解产物中物质的种类和比例进行测定,再反向推断多糖的部分或全部结构。因此可以作为分析天然多糖的工具酶。
在造纸工业中,与β-木聚糖酶等半纤维素降解酶组合,可以从纸浆中除去半纤维素并改善纸张质量。研究发现β-甘露聚糖酶和木聚糖酶在纸浆漂白时有协同作用,同时使用可使漂白效果大幅提升同时减少了因为碱的使用而造成对环境的污染,保护了生态环境[31]。
在环境治理方面,减少环境中的β-甘露聚糖物质可以降低消化道内容物的粘度,减少粪便排泄,减少环境污染[20,31],还可以吸附霉菌毒素,减少抗生素和其他化学药物的使用。
在纺织行业,苎麻作为纺织原料由来已久,但其中的胶质必须去掉,而传统的脱胶需要高温、高压、强酸、强碱等条件,这不仅使得纤维遭到损坏,也对环境造成了极大的污染。而β-甘露聚糖酶对于脱胶具有特异性,使得苎麻脱胶彻底、对纤维素损坏小、提高了纤维品质。另外,β-甘露聚糖酶可有效去除附着在纺织品上的多余染料,这样氯和碱的用量就会减少,保护了生态环境;此外,在纺织品的加工过程中,β-甘露聚糖酶还可以对布料进行软化、打磨、抛光,进一步提高纺织品质。
在饲料工业中,谷物及其副产品中普遍存在着一种抗营养因子β-D-甘露聚糖,其在单胃动物的消化道内形成凝胶状,使其消化道内容物具有较强的黏性,且由于家禽和猪等单胃动物,体内不能产生甘露聚糖酶和木聚糖酶等,而很多用作饲料的植物组织含有木聚糖和甘露聚糖等半纤维素,这使得动物饲料造成了极大的浪费。而甘露聚糖酶起到消除抗营养因子的作用,促进畜禽生长,减少养殖污染[32-33];另一方面,β-甘露聚糖酶水解产生的甘露寡糖可以起到益生元的作用,调节肠道微生态平衡,抑制有害菌的生长,促进有益菌的生长。
在农业生产中,甘露聚糖类物质作为半纤维素的第二大组分,广泛分布于自然界中,它是所有豆科植物细胞壁的主要组成成分。消除富含葡萄糖的β-甘露聚糖对豆类的干扰,提高豆粕的能量转化率[34-35]。
在制药行业,甘露低聚糖是一种水溶性的膳食纤维,能够降低血清中胆固醇和甘油三酯的含量。甘露低聚糖是双歧杆菌的增殖因子,能够调节肠道消化系统功能,改善肠道菌群结构,故甘露低聚糖具有保护肝脏、提高免疫力、促进肠道蠕动、降低胆固醇等生理功能;此外,β-甘露聚糖酶水解富含有甘露聚糖植物胶而成的甘露糖可用于治疗尿路感染。
在食品行业,β-甘露聚糖酶具有降解植物胶生成甘露低聚糖,作为益生元可有效降低人体胆固醇和血糖水平,增强人体免疫力和促进人体肠道益生菌的生长。还可以降解咖啡豆中的甘露聚糖,避免冻干过程中凝胶的形成,以利于甘露聚糖的充分利用,提高咖啡质量。同时,β-甘露聚糖酶在椰油的提取、增稠剂降解及果汁澄清等加工过程中也有重要作用。此外,甘露糖也有甜味可以替代蔗糖作为食品甜味剂。
除此之外,β-甘露聚糖酶在洗涤肥皂业、油气开采业、及生物质燃料等行业也有重要作用。
6 展望
微生物β-甘露聚糖酶的研究近年来取得了很大进展,已经可以进行工业化量产。但能够集大规模工业化生产,环境适应性强,活性高的的高品质β-甘露聚糖酶仍是当前之急需。基于此,不断提高微生物酶生产水平,研究β-甘露聚糖酶的分子结构及合成的调控机制,克隆和酶基因的高效表达,新型甘露聚糖酶的开发,促进工业应用是未来努力的主要方向。
