Functional role of long non-coding RNA CASC19/miR-140-5p/CEMIP axis in colorectal cancer progression in vitro
2019-05-08XiaoDongWangJianLuYunShouLinChaoGaoFengQi
Xiao-Dong Wang, Jian Lu, Yun-Shou Lin, Chao Gao, Feng Qi
Abstrac t BACKGROUND Long non-coding RNAs (lncRNAs) are widely involved in tumor regulation.Nevertheless, the role of the lncRNA cancer susceptibility 19 (CASC19) in colorectal cancer (CRC) has yet to be fully clarified.AIM To explore the effect of CASC19 on proliferation and metastasizing ability of CRC cells.METHODS CASC19 expression in human CRC tissues, pair-matched adjacent normal colon tissues, and CRC cells was detected using quantitative real-time PCR (qRT-PCR).CASC19 expression, as well as its relation to overall survival, was extrapolated by Kaplan-Meier survival analysis together with multivariable Cox regression assay.In vitro experiments were performed to confirm whether CASC19 regulates CRC cell invasion, migration, proliferation, and apoptosis.RESULTS CASC19 expression was markedly upregulated in CRC tissues and CRC cell lines(P < 0.05). qRT-PCR revealed that CASC19 expression was higher in 25 tissue samples from patients with aggressive CRC compared with the 27 tissue samples from patients with nonaggressive CRC (P < 0.05). Higher CASC19 expression was associated with poorer patient prognoses. Furthermore, in vitro experiments demonstrated that CASC19 overexpression enhanced CRC cell invasion,migration, and proliferation. CASC19 overexpression enhanced the expression of cell migration inducing hyaluronidase 1 (CEMIP) and epithelial-mesenchymal transition markers. MiR-140-5p was found to be able to bind directly to CASC19 and CEMIP. Overexpression of miR-140-5p reversed the effect of CASC19 on cell on different terms, provided the original w ork is properly cited and the use is non-commercial. See:http://creativecommons.org/licen ses/by-nc/4.0/
Key words: Colorectal cancer; Long non-coding RNA; MicroRNA; Proliferation;Metastasis
INTRODUCTION
Colorectal cancer (CRC) is a tumor that is increasingly common in the modern world[1]. Tumor metastasis is one of the most important causes of poor prognosis for patients with CRC. At the time of diagnosis, approximately 20%-25% of patients with CRC are found to have liver metastasis. At the same time, liver metastasis occurs in up to 40%-50% of patients after resection of primary CRC[2]. Although current methods for the diagnosis and therapy of CRC have achieved remarkable progress,tumor metastasis remains an important factor affecting the survival of patients[3]. In recent years, gene therapy has become an intense focus of research. Transporting tumor suppressor genes or non-coding RNAs via nanocarriers may be a new option for cancer therapeutics[4]. Therefore, a thorough understanding of the molecular pathophysiological pathways underlying CRC is crucial to developing an effective therapeutic strategy.
Non-coding RNAs include microRNAs (miRNAs) and long non-coding RNAs(lncRNAs). MiRNAs bind to the 3'-untranslated regions (3'-UTR) of the message RNA(mRNA) of the target genes, resulting in mRNA degradation and inhibition of the translation process. LncRNAs are RNAs that are longer than 200 nucleotides. The existing literature primarily investigates the regulatory roles of lncRNAs in several biological processes[5,6]. Dysregulation of lncRNAs is observed in various types of cancers, including breast cancer[7], oesophageal cancer[8], hepatocellular carcinoma[9-11],lung cancer[12], gastric cancer[13], and CRC[14-18]. LncRNA dysregulation has been discovered to be closely related to cancer progression. For example, overexpression of the lncRNA n335586 contributes to cell migration and invasion in hepatocellular carcinoma[19], w hile the lncRNA CASP5 facilitates the migration and invasion of human glioblastoma cells[20]. The regulatory mechanism of lncRNAs is still not clearly understood, and its possible role in cancer has been hyp othesized to be as a competing endogenous RNA (ceRNA) for sponge miRNAs. For instance, the lncRNA UCA1 may adsorb microRNA (miRNA/miR)-182, thereby affecting the expression of its downstream target gene PFKFB2 and promoting glioma metastasis[21]. The lncRNA PVT1 enhances colon cancer metastasis by altering the miR-30d-5p/RUNX2 axis[22].CRC progression has recently been discovered to be associated with endogenous lncRNA sponges. The cancer susceptibility 19 (CASC19) is a 324 bp lncRNA that is located on chromosome 8q24.21. Several lines of evidence suggest that the expression of CASC19 is overregulated in CRC, and this may play an oncogenic role in CRC progression[23-25]. How ever, the mechanism by w hich CASC19 regulates CRC progression is not fully understood.
The cell migration ind ucing hyaluronid ase 1 (CEMIP) gene is located on chromosome 15q25 and encodes a 150 k Da protein. CEMIP is originally described as an inner ear protein and its mutation leads to hearing loss[26]. CEMIP has traditionally been linked to hyaluronic acid d epolymerization[27]. Recent find ings indicate that CEMIP may be involved in tumor d evelop ment and may p romote tumor cell proliferation and metastasis. For instance, the high expression of CEMIP is associated w ith a p oor prognosis of prostate cancer[28], gastric cancer[29], and CRC[28,30-34]. These rep orts suggest that CEMIP contributes to cancer heterogeneity and may be a potential therapeutic target.
Our present study d emonstrated that CRC possesses a characteristic alteration in CASC19 exp ression p rofile that is related to CRC progression. Overexp ression of CASC19 promotes CRC p rogression. In ad dition, mechanism analysis show ed that CASC19 p ositively regulates CEMIP expression via sponge miR-140-5p, thereby exerting a carcinogenic effect in CRC progression. Future therap ies targeting the CASC19/miR-140-5p/CEMIP axis may be beneficial in CRC.
MATERIALS AND METHODS
Patients and tissue specimens
This study included 52 patients w ho w ere pathologically diagnosed as having CRC and received surgical treatment betw een January 2015 and December 2016 at Tianjin Med ical University General Hospital. Dissected tumor and adjacent normal colonic mucosal tissues (as samples taken from areas w ell close to the section margin) were immediately frozen in liquid nitrogen and stored at -80 °C. All the patients did not receive neoadjuvant therapy. All patients provided w ritten informed consent. This stud y was approved by the Medical Ethics Committee of Tianjin Medical University General Hosp ital (Ethical No. IRB2015-YX-018) and w as cond ucted in strict compliance with the Declaration of Helsinki.
Cell culture and transfection
Healthy colon epithelial cell line (FHC) and CRC cell lines (HCT-116, SW480, Lovo,and SW620) w ere purchased from the Chinese Academy of Sciences (Shanghai,China). All cells were maintained in DMEM medium supplemented with 10% FBS in a controlled environment of 5% CO2at 37 °C. Cell medium was replaced every 2 d.Before transfection, a total of 2.5 × 104cells per well were seeded onto 6-well plates and incubated for 24 h, then the culture medium was discarded and 100 nmol/L of CASC19 overexpressing plasmid (CASC19-p; Genechem, Shanghai, China), 200 nmol/L of siRNA mixture, 100 nmol/L of miR-140-5p mimic (Ribobio, Guangzhou,China), or 200 nmol/L of miR-140-5p inhibitor (Ribobio, Guangzhou, China) w as used for cell transfection. The pc DNA 3.1 negative control (CASC19-p NC),pcNDA 3.1-CASC19 (CASC19-p), CASC19 siRNAs (siCASC19), CEMIP siRNAs(siCEMIP), and pcNDA3.1-CEMIP (CEMIP-p) w ere purchased from Genechem(Shanghai, China). MiR-140-5p mimic, miR-140-5p inhibitor, mimic NC, and inhibitor NC were obtained from Ribobio (Guangzhou, China).
Luciferase reporter system
The luciferase reporter system was constructed by Genechem (Shanghai, China) as previously described[35]. Briefly, site-directed mutations were introduced into the CASC19 or CEMIP binding site of miR-140-5p (Quik Change Lightning Site-Directed Mutagenesis Kit, Stratagene, United States). The 3'-UTR fragment of wild-type (Wt)and mutant (Mut) CASC19 or CEMIP was then sub-cloned into the p GL3 luciferase vector (Promega, United States) by PCR, respectively. The miR-140-5p mimic was cotransfected w ith the vector into 293T cell lines for 12 h in 96-well plates using an enhanced infection solution (containing 5 μg/mL polybrene) (Genechem, Shanghai,China). The 293T cells were then cultured for 24 h and lysed to analyze the luciferase activity. Renilla (Promega, United States) activity was used as the internal control.
Cell cycle assay
A flow cytometric technique was used to perform cell cycle analysis as detailed in a prior study[36]. In brief, HCT-116 and SW480 cells were harvested after transfection,and cell fixation w as d one w ith 70% p re-cooled ethanol at -20 °C overnight.Subsequently, cell staining was carried out using propidium iodide (PI, 50 μg/mL;Solarbio, Tianjin, China) and RNase A (0.1 mg/mL, Sigma, United States) at room temperature for 30 min. Then, the cells were analyzed w ith a flow cytometer (BD FACSCato II Flow Cytometer, BD Biosciences, Franklin Lakes, United States). All data pertaining to cell cycle studies are described in terms of the percentage of cell distributions in different phases (G0/G1, S, and G2/M).
Cell proliferation assay
The cell counting kit-8 (CCK-8) assay w as used to assess the ability of SW480 and HCT-116 cells to proliferate. Cells (2 × 103) w ere added to each well of a 96-well plate with 100 μL of culture medium. CCK-8 solution (10 μL; Dojindo Laboratories, Japan)was applied into each well from day 0 to day 5. The cells were left to incubate for 3 h at 37 °C. A microplate luminometer (Bio-Rad Laboratories, United States) w as then used to quantify absorbance values at 450 nm.
Wound-healing and transwell assays
The abilities of cells to migrate and invade were assessed by wound-healing assay and Transwell assay with Matrigel (BD Bioscience, United States). Briefly, for the woundhealing assay, transfected cells were allowed to proliferate in 6-well plates until a confluent monolayer was achieved. The cell layer was gently scratched with a 200 μL pipette tip to inflict a wound. PBS was used to rinse the cells to remove debris and wound healing was allowed to take place for the next 24 h. The area of open wound covered by cells was described in terms of “percent wound closure (%)” [(Scratch distance0h- Scratch d istance24h)/Scratch distance0h× 100%, the distances w ere measured with Image J]. For the transwell assay, 200 μL of medium without serum containing 1 × 105cells was applied onto the upper chamber, while 600 μL of medium with 10% fetal bovine serum (FBS) was added to the bottom chamber. The cells were left to incubate for 24 h at 37 °C. The membranes were then removed and cells were fixated with 4% paraformaldehyde before undergoing a 10-min staining period with 0.1% crystal violet. An inverted microscope was used to count the total cell number.
Apoptosis assay
For the apoptosis assay, 200 μL of PBS was used to suspend cells. PI and fluorescein isothiocyanate (FITC) conjugated Annexin V (BD Biosciences, Franklin Lakes, NJ,United States) were then added into the suspension and left to incubate for 20 minutes at room temperature. A flow cytometer (FACSVerse, BD Biosciences, San Diego, CA,United States) was used to analyze cells.
RNA extraction and quantitative real-time PCR (qRT-PCR)
Total RNA was extracted with an RNA extraction kit (QIAGEN, Shanghai, China) and 1 μg of total RNA was added to a 20 μL system. The GoScript Reverse Transcription system (Promega, United States) was used for reverse transcription of CASC19 and CEMIP, and miR-140-5p was reverse transcribed by the stem-loop method (QIAGEN,Shanghai, China). GAPDH and U6 served as internal controls. The primer sequences are as follows: forward, 5'-GAGGAAGGCAGCACAATGATG-3' and reverse, 5'-CTTG CCAGTGTCTTCTCCTGA-3' for CASC19; forw ard, 5'-GCTCTGGGATTTAAGG CAGC-3' and reverse, 5'-ATTGGAGCCATGGACTGTGA-3' for CEMIP; forward, 5'-CAGTGGTTTTACCCTATGGTAG-3' and universal primer, 5'-TGGTGTCGTGGA GTCG-3' for miR-140-5p; forward, 5'-TGCACCACCAACTGCTTAGC-3' and reverse 5'-G G C A T G G A C T G T G G T C A T G A G-3' f o r G A P D H; f o r w a r d, 5'-CTCGCTTCGGCAGCACA-3' and reverse, 5'-AACGCTTCACGAATTTGCGT-3' for U6. All PCR reactions lasted 40 cycles, and the relative fold change of gene expression was derived using the 2-ΔΔCtmethod.
Subcellular fractionation
Cellular localization of CASC19 was determined by subcellular fractionation using the PARIS Kit (Thermo Fisher Scientific, United States). Briefly, cytoplasmic and nuclear RNA were first extracted. The expression of CASC19 in the cytoplasmic and nuclear fractions w as determined by qRT-PCR. GAPDH and U6 served as the cytoplasmic and nuclear controls, respectively.
RNA immunoprecipitation (RIP) assay
RIP assay (Magna RIPTM RNA Binding Protein Immunoprecipitation Kit, Millipore,United States) w as utilized to verify the binding association between miR-140-5p and endogenous CASC19 in CRC cells. Briefly, harvested HCT-116 and SW480 cells w ere resuspended in cell RIP lysis buffer and kept on ice for 20 min. The RIP lysate w as then centrifuged and 100 μL of the supernatant w as removed and added to each beadantibod y [anti-Argonaute 2 (Ago2) or negative control Ig G] comp lex in RIP immunoprecipitation buffer. All tubes were left to incubate overnight in a rotator at 4°C. The next day, immunoprecipitation tubes were centrifuged briefly and placed on a magnetic separator to collect the bead-antibody-RNA complexes. Protein K w as used to purify the RNA. Purified RNA was then analyzed by RT-PCR.
Western blot analysis
Specimens and CRC cells were lysed in RIPA to obtain total protein, and 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) was used to separate 40 μg of each p rotein sample w ith the proteins subsequently reapp lied to polyvinylidene fluoride membranes (Millipore, United States). These membranes containing blotted proteins w ere exposed to 0.5% bovine serum albumin to block endogenous reactions, followed by overnight probing at 4 °C with anti-CEMIP(1:1500, Novus, United States), anti-Vimentin (1:1500, CST, United States), anti-Ncadherin (1:1500, CST, United States), anti-E-cad herin [1:1500, Cell Signaling Technology (CST), United States], anti-poly AD-ribose polymerase (PARP) (1:2000,CST, United States), anti-caspase 3 (1:2000, CST, United States), anti-caspase 7 (1:2000,CST, United States), and anti-β-actin (1:3000, CST, United States) antibodies. The next day, membranes were rinsed and subjected to 2-h room temperature incubation with diluted secondary horseradish peroxidase (HRP)-marked antibodies (1:2500, CST,United States). An enhanced chemiluminescence detection kit (Millipore, United States) was used to detect immunoreactive protein bands. Images were acquired with a G-Box system (Syngene, United States) and analyzed w ith Image J (National Institute of Mental Health, United States).
Immunohistochemistry assay
Immunohistochemistry assay was used to determine CEMIP expression in CRC tissues and adjacent normal colonic mucosal tissues as described in our previous study[37]. Briefly, tissue samples were fixed in 4% paraformaldehyde, embedded in paraffin, and sliced into 4 μm sections. Dehydration was carried out with xylene and a gradient of ethanol solution before endogenous peroxidase blocking was carried out with a 3% H2O2solution. Antigen retrieval was performed with heated sodium citrate solution (92 °C-95 °C, 10 nmol/L, p H 6.0) for 5 min and 1% goat serum was added to the sections at room temperature for 2 min to block the sections. Rabbit anti-human CEMIP antibody (1:150; CST, United States) was applied onto the sections and left overnight to incubate at 4 °C. After incubation with the secondary antibody for 1 h at RT, freshly prepared diaminobenzidine was then added to the sections that were then stained w ith hematoxylin. A light microscope (Leica Microsystems, Wetzlar,Germany) w as used to visualize slides and brown particles in the cytoplasm or cytomembranes were taken to indicate positive staining.
Statistical analysis
All data are expressed in terms of the mean ± SD. Continuous data w ere analyzed using the t-test or the one-w ay analysis of variance (ANOVA). The relationship between CASC19 expression and clinicopathological features was determined by onew ay analysis of variance, the Chi-squared test, or the Fisher's exact test. The prognostic value of CASC19 in CRC patients was explored by Cox regression analysis.Kaplan-Meier method s and log-rank tests w ere used to carry out survival analyses.The correlations betw een CASC19, miR-140-5p, and CEMIP w ere analyzed using Spearman or Pearson correlation analysis. All statistical analyses w ere carried out with the use of GraphPad Prism (GraphPad Software, Version 5.0, United States) and SigmaPlot softw are (SPSS 22.0, United States). P-values < 0.05 w ere consid ered statistically significant. All results are representative of three repeated experiments.
RESULTS
CASC19 is upregulated in CRC tissues and CRC cell lines, and high expression of CASC19 indicates a poor prognosis
CASC19 expression was analyzed in 52 pairs of CRC tissues and adjacent normal colonic mucosal tissues using qRT-PCR. CASC19 exhibited significantly increased expression in CRC tissues in contrast to adjacent healthy colonic mucosal tissues (P <0.05) (Figure 1A). Consistently, the CASC19 expression in CRC cell lines w as markedly elevated in comparison to FHC cells (P < 0.05) (Figure 1B). For further in vitro studies, HCT-116 and SW480 cell lines were used.
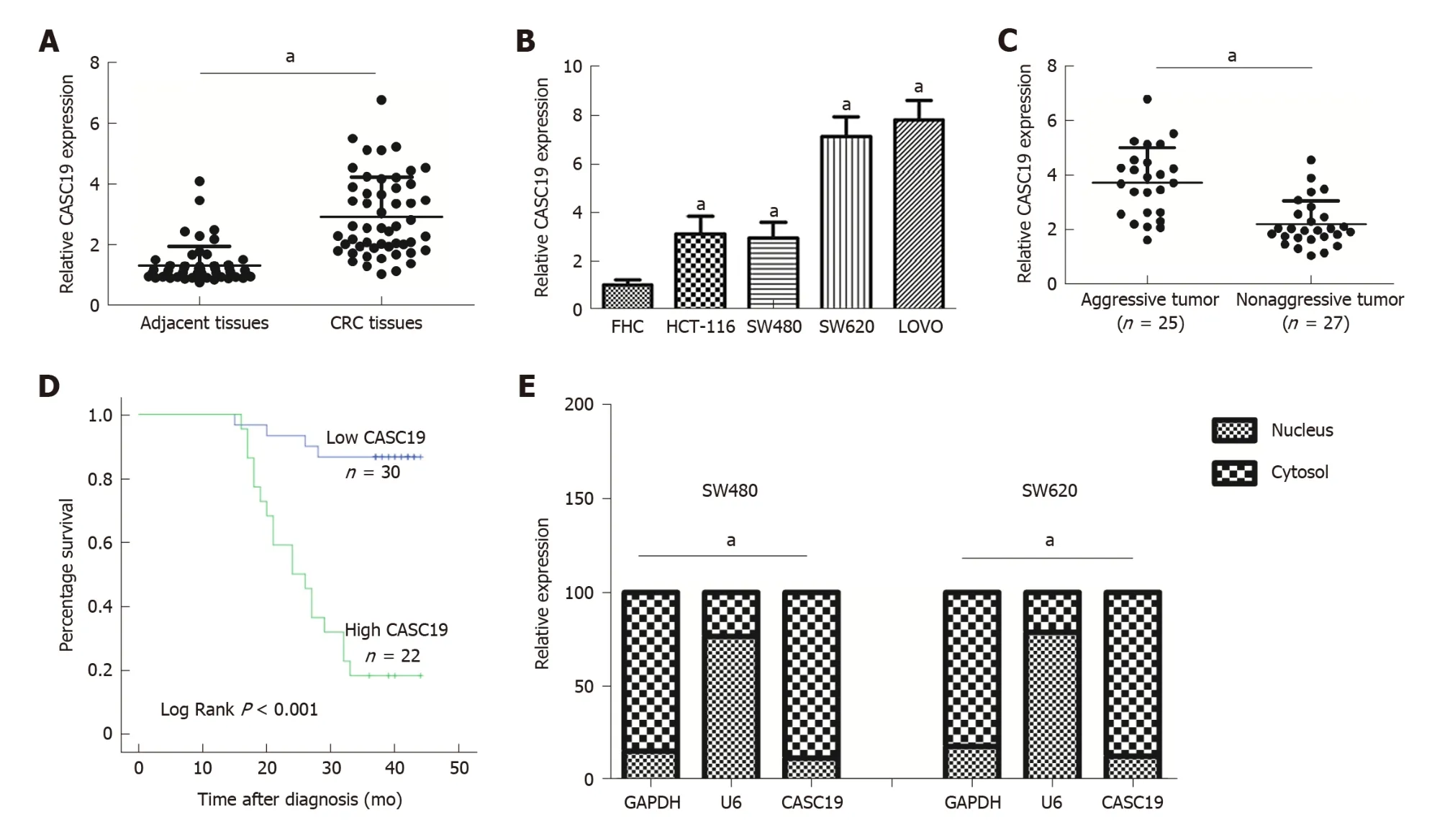
Figure 1 Differential expression of cancer susceptibility 19 in colorectal cancer tissues and cell lines. A: Differential cancer susceptibility 19 (CASC19)expression in CRC tissues and adjacent normal colon mucosa tissues. B: Differential CASC19 expression in colorectal cancer (CRC) cell lines and FHC cells. C:Differential CASC19 expression in aggressive tumor tissue samples and nonaggressive tumor tissue samples. D: Kaplan-Meier analyses of the correlations between CASC19 and overall survival of 52 patients with CRC. E: Cellular localization of CASC19 in CRC cells. Data are reported as the mean ± SD. a P < 0.05. CASC19:Cancer susceptibility 19; CRC: Colorectal cancer.
We also focused on determining the association between expression of CASC19 and clinical phenotypes of patients w ith CRC. Participants w ere stratified into an aggressive tumor group (n = 25) and a non-aggressive tumor group (n = 27) based on whether the patient had or not lymph node or liver metastasis. Results showed that aggressive CRC tumors had higher CASC19 expression (P < 0.05) (Figure 1C). Next,we calculated the average of relative CASC19 expression levels in tumor tissues, and the average value (2.9310) was determined to be the cutoff point between “High CASC19” and “Low CASC19”. Results showed that 22 samples were included in the high CASC19 group w hile another 30 w ere includ ed in the low CASC19 group.Subsequent correlation analysis revealed that CASC19 expression was associated with liver metastasis (P < 0.01), lymphatic metastasis (P < 0.01), and TNM stage (P < 0.05)(Table 1) (Supplementary Figure 1). Significant associations w ere not present between CASC19 expression and other clinical parameters such as tumor size, local invasion,age, and gender (P > 0.05) (Table 1). Kaplan-Meier survival curves demonstrated that patients w ho possessed raised expression of CASC19 experienced shorter survival durations in contrast to those who had low er CASC19 expression (log-rank P < 0.001)(Figure 1D). We then carried out multivariable Cox regression analysis to further determine the impact of clinicopathological features and expression of CASC19 on the overall survival times of patients w ith CRC. The find ings ind icated that CASC19 expression [the exp onent of B (Exp (B) = 8.893, 95% confidence interval (CI): 2.596-30.464, P = 0.001] was an independent prognostic factor for CRC (Table 2).
Moreover, the results of subcellular fractionation confirmed that CASC19 w as predominantly localized in the cell plasma (Figure 1E). Taken together, our findings demonstrated that CASC19 w as a highly expressed lncRNA in CRC and is highly involved in CRC metastasis.
CASC19 promotes proliferation and inhibits apoptosis of CRC cells in vitro
To exp lore the role of CASC19 in CRC cells, SW480 and HCT-116 cells w ere transfected w ith CASC19-p or siCASC19. CASC19 was increased significantly in the CASC19-p group (CASC19 overexpression) in contrast to the control group (P < 0.05)and w as substantially attenuated in the siCASC19 group (CASC19 knockd ow n) (P <0.05) (Figure 2A). CASC19-p NC or siCASC19 NC did not alter CASC19 expression in CRC cells (Figure 2A).
Ad d itionally, the CCK-8 assay w as used to assess the impact of CASC19 on the proliferation of SW480 and HCT-116 cells. Figure 2B demonstrates that HCT-116 and SW480 cells exp erienced augmented p roliferation in the p resence of CASC19 overexpression (P < 0.05). Conversely, knockdow n of CASC19 inhibited HCT-116 and SW480 cell proliferation (P < 0.05).
Interestingly, high CASC19 expression in CRC cells led to marked ly d ecreased casp ase-3, caspase-7, and PARP (P < 0.05 for all) (Figure 2C). Overexp ression of CASC19 d ecreased the apop totic rate of CRC cells (P < 0.05) (Figure 2D), w hile CASC19 knockd ow n enhanced casp ase-7, casp ase-3, and PARA p rotein levels.Furthermore, CASC19 knockdow n marked ly elevated the p ercentage of ap optoticcells (P < 0.05 for all) (Figure 2D). Cell cycle assay rev ealed that CASC19 overexpression led to a higher proportion of SW480 and HCT-116 cells that w ere in the S phase. This proportion w as significantly reduced in cells that had suppressed CASC19 levels. It can be conclud ed that CASC19 p romoted CRC cell grow th, w hile knockdown of CASC19 expression could impede CRC cell growth.
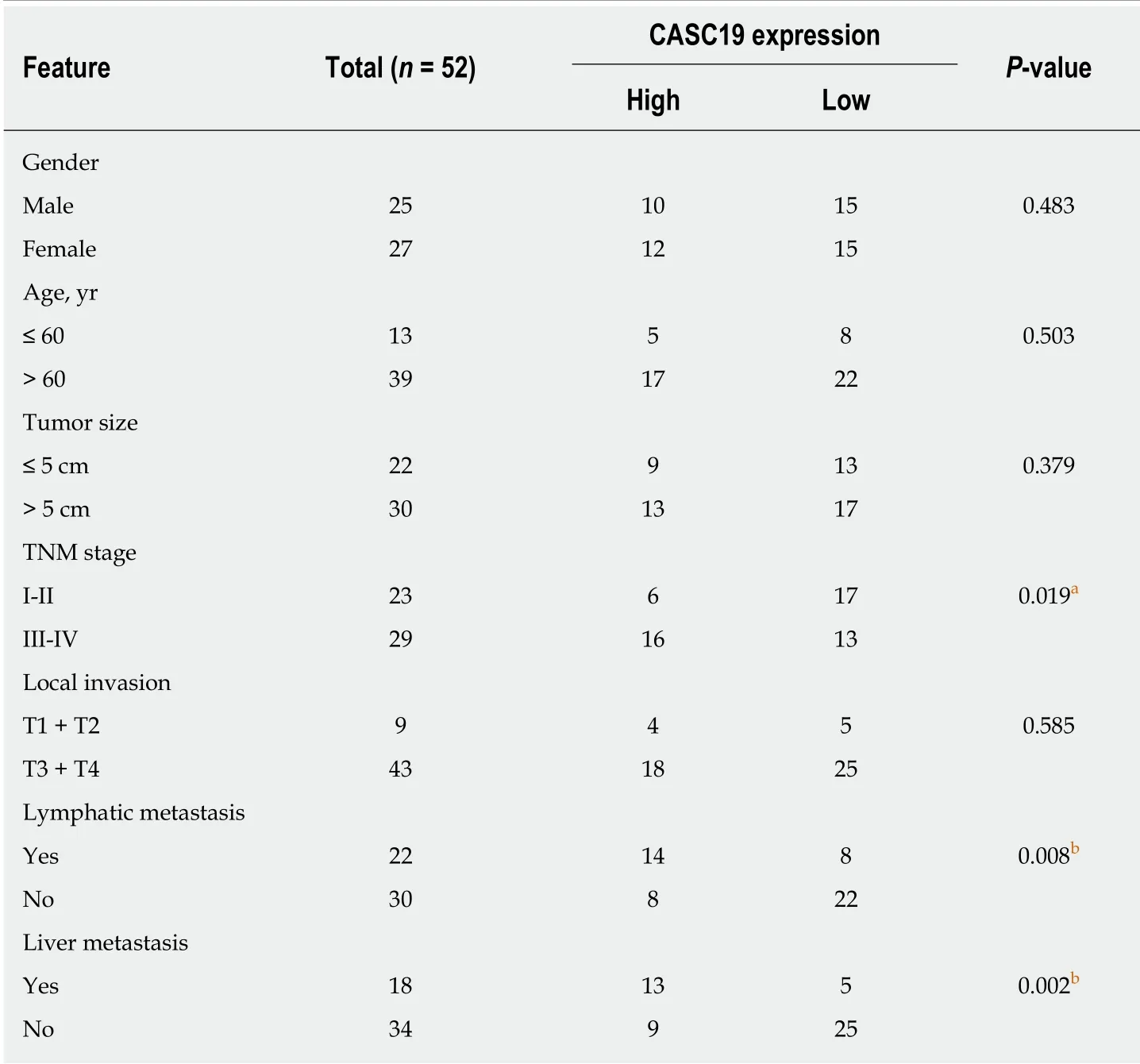
Table 1 Associations of cancer susceptibility 19 expression with clinicopathologic features of patients with colorectal cancer
CASC19 promotes CRC cell migration and invasion in vitro
The w ound-healing and transw ell assays w ere used to d etermine the imp act of CASC19 exp ression on CRC cell migration and invasion, resp ectively. Cells transfected with CASC19-p completed wound healing at a faster rate in comparison to cells that w ere transfected w ith CASC19-p NC (P < 0.05) (Figure 3A). HCT-116 and SW480 cells that w ere induced to overexp ress CASC19 w ere observed to und ergo higher invasion rates (Figure 3B). Whereas, the migration and invasion abilities w ere inhibited in cells that had CASC19 knockdown (Figure 3A and B) (P < 0.05).
Given the crucial function of epithelial-mesenchymal transition (EMT) in the malignant transformation and invasive properties of cancer cells, we sought to clarify the impact of CASC19 on EMT marker expression in CRC cell lines. E-cad herin w as downregulated (P < 0.05), w hile Vimentin and N-cadherin were upregulated (P < 0.05 for all) in HCT-116 and SW480 cell lines that had upregulated CASC19 (Figure 3C). In contrast, E-cad herin exp ression w as increased (P < 0.05), and the exp ression of Vimentin and N-cadherin was decreased (P < 0.05 for all) in HCT-16 and SW480 cells that had suppressed CASC19 levels (Figure 3C). Together, these d ata indicate that CASC19 promotes CRC cell metastasis.
CASC19 promotes CEMIP expression by targeting miR-140-5p
To und erstand how CASC19 affects the CRC cell migration and p roliferation, a ce RN A n etw o r k an aly sis w as car r ied ou t u sin g star Base v 2.0(http://starbase.sysu.edu.cn/ind ex.php)[38,39]. Interestingly, CASC19 w as positively correlated w ith CEMIT expression. We examined the protein and m RNA levels of CEMIP in CRC cell lines and tissues. CEMIP expression w as augmented in CRCtissues in contrast to healthy normal colonic mucosa at both the mRNA and protein levels (P < 0.05 for all) (Figure 4A and B). Similarly, the mRNA levels of CEMIP in CRC cells lines was upregulated in contrast to levels found in FHC cells (P < 0.05)(Figure 4C). Furthermore, Spearman's correlation analysis revealed CASC19 expression was positively correlated CEMIT m RNA expression in CRC tissues(Pearson r = 0.809, P < 0.001) (Figure 4D). Additionally, we examined the effect of CASC19 on CEMIP expression in SW480 and HCT-116 cell lines. The mRNA and protein levels of CEMIP were upregulated after CASC19 overexpression in both cell lines, while suppression of CASC19 expression decreased CEMIP protein and mRNA levels in the two cell lines (P < 0.05 for all).
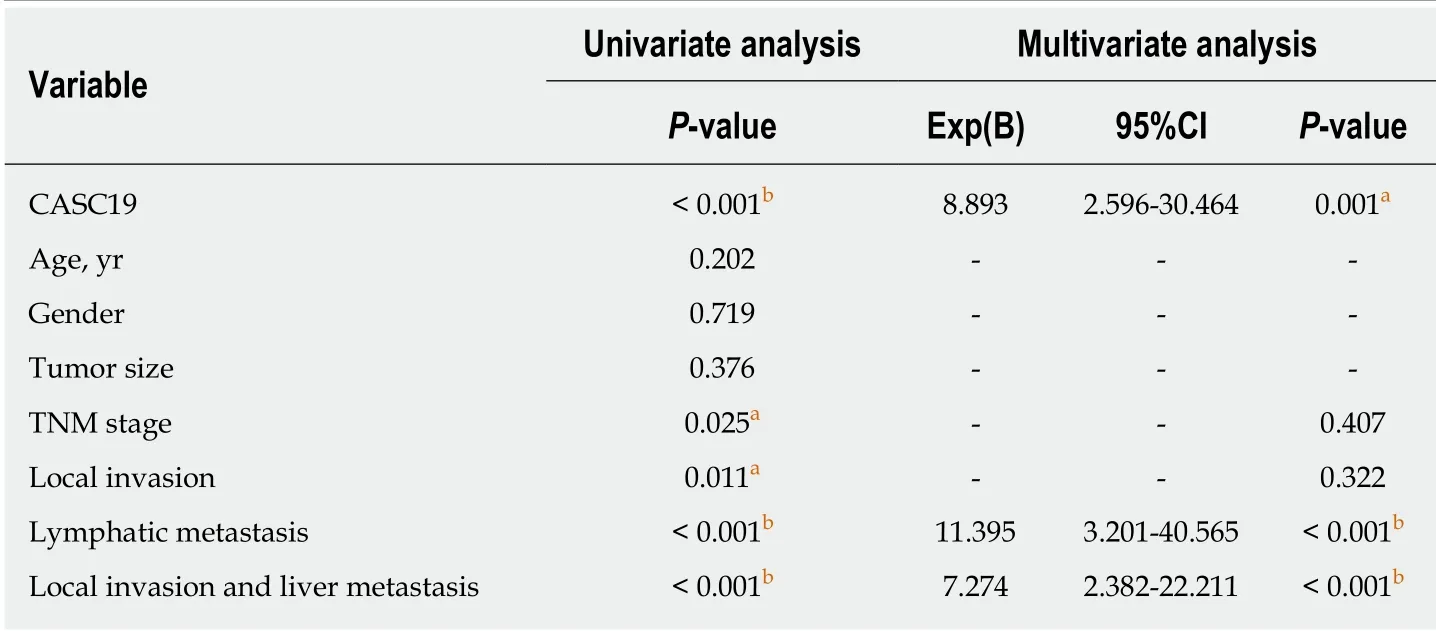
Table 2 Univariate and multivariate analyses of variables influencing survival of 52 patients with colorectal cancer
It has been demonstrated that lncRNAs are able to bind directly to miRNA and function as ceRNAs[40]. We next predicted miR-140-5p to be a potential target of CASC19 by using starBase v 2.0 program (http://starbase.sysu.edu.cn/index.php)[38,39](Figure 4F). We went on to examine the expression of miR-140-5p in CRC cell lines and tissues. CRC tissue samples were found to possess a much lower miR-140-5p level in contrast to adjacent healthy colonic mucosal tissues (P < 0.05) (Figure 4G).
Besides, miR-140-5p was suppressed in CRC cell lines in contrast to FHC cells (P <0.05) (Figure 4G). CASC19 and miR-140-5p expression appeared to negatively correlate in CRC tissues, based on Spearman's correlation analysis (Pearson r = -0.660,P < 0.001) (Figure 4H). The anti-Ago2 RIP assay was performed to verify the presence of direct bind ing betw een CASC19 and miR-140-5p. Endogenous CASC19 w as specifically enriched in miR-140-5p mimic transfected CRC cells in contrast to the NC mimic group (P < 0.05) (Figure 4I). A luciferase reporter assay was carried out to determine if CASC19 could directly target miR-140-5p. The luciferase activity was reduced in wild-type CASC19 3'-UTR and miR-140-5p mimic co-transfected 293T cells in comparison to wild-type CASC19 3'-UTR and miR-140-5p mimic NC co-transfected 293T cells (P < 0.05) (Figure 4J).
Interestingly, CEMIP is a potential target gene of miR-140-5p [predicted by Tar get Scan (h ttp://w w w.tar getscan.or g/v er t_71/) an d m iRBase(http://www.mirbase.org/)] (Figure 4K). Spearman's correlation analysis showed that CEMIP correlated negatively with miR-140-5p in CRC tissues (Pearson r = -0.657,P < 0.001) (Figure 4L). CEMIP is a direct target of miR-140-5p, as revealed by luciferase reporter assays (Figure 4M).
Furthermore, it w as suggested that the CEMIP p rotein exp ression w as d ow nregulated in siCASC19 and NC inhibitor co-transfected CRC cells, a phenomenon which could be reversed with the addition of miR-140-5p inhibitor (P <0.05) (Figure 4N). CEMIP expression was markedly raised after transfecting CRC cells with CASC19-p and NC mimic. Moreover, the effects of CASC19 were reversed by the addition of miR-140-5p mimic to CRC cell lines (P < 0.05) (Figure 4O). Collectively,these finding indicate that CASC19 is able to function as a ceRNA and indirectly regulate CEMIP expression by sponging miR-140-5p.
MiR-140-5p reverses the promoting effect of CASC19 on the proliferation and metastasizing ability of CRC cells
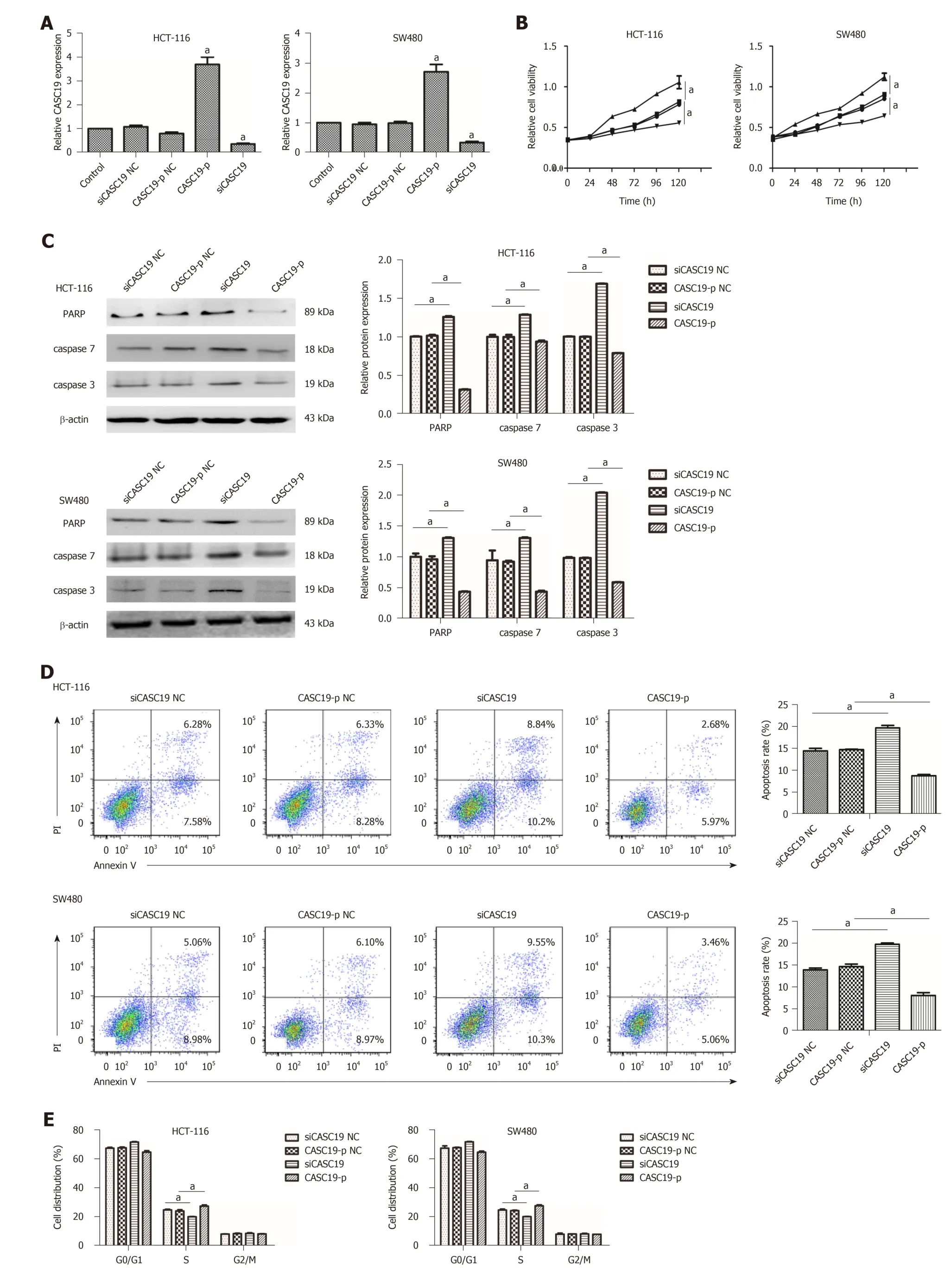
Figure 2 Role of cancer susceptibility 19 in the proliferation and migration of colorectal cancer cells. A: Expression of cancer susceptibility 19 (CASC19) in colorectal cancer (CRC) cells transfected with siCASC19 or CASC19-p and their NCs was quantified by quantitative real-time PCR. B: Growth curves of HCT-116 and SW480 cell lines that were transfected with CASC19-p or siCASC19 were analyzed using the Cell Counting Kit-8 assay. C: Western blot was used to determine the expression of caspase 3, caspase 7, and PARP in HCT-116 and SW480 cells that were transfected with either CASC19-p or siCASC19. The grayscale value ofproteins was measured with Image J. D: Cell apoptosis rate was detected by flow cytometry after transfection with CASC19-p or siCASC19. E: The proportion of cells in each cell cycle phase was detected using flow cytometry after cells were transfected with CASC19-p or siCASC19. Data are reported as the mean ± SD. a P < 0.05.
To confirm w hether CASC19 exerts its function through sponging miR-140-5p in CRC tissues, CCK-8 and transw ell assays w ere p erformed. Results d emonstrated that knockdown of CASC19 or CEMIP expression significantly inhibited the proliferation and invasion of HCT-116 and SW480 cells (P < 0.05) (Figure 5A-D). The add ition of miR-140-5p inhibitor reversed this effect. Moreover, miR-140-5p inhibitor w as able to promote the proliferation and invasion abilities of cells. Conversely, overexpression of CASC19 or CEMIP in the cell lines promoted the proliferative and invasive capacities of HCT-116 and SW480 cells, w ith this effect reversed by miR-140-5p mimic.Furthermore, miR-140-5p alone significantly inhibited the proliferative and invasive cap acities of both cell lines (P < 0.05) (Figure 5A-D). Lastly, the exp ression of Ecad herin and vimentin in transfected CRC cells w as analyzed. Consistent w ith the p revious results, the effect of knockd ow n or overexp ression of CASC19 on the expression of the markers was reversed by miR-140-5p inhibitor or miR-140-5p mimic(Figure 5E and F). These d ata suggested that miR-140-5p is able to counteract the effect of CASC19 on the growth and metastasizing abilities of CRC cells in vitro.
DISCUSSION
Studies have confirmed that dysregulation of lncRNAs is closely related to the occurrence and development of various tumors. LncRNAs may become potential targets for the development of future chemotherapeutic medications[41,42]. Therefore,studying the clinical significance and function of lncRNA along with its possible mechanisms of action may contribute towards the gene therapy in cancer. Recently,researchers have demonstrated that lncRNAs are involved in the progression of CRC.For instance, the lncRNA SUMO1P3 drives CRC growth and metastasis[16,43,44]. Lymph node metastasis and poor CRC prognosis have been linked to the upregulation of the lncRNA BANCR[43]. The lncRNA HAND2-AS1 sponging miR-1275 suppresses CRC progression by upregulating KLF14[44]. Previous studies demonstrate that the lncRNA CASC19 may promote the progression of several cancers, and it has been suggested that CASC19 may function as a biomarker for CRC poor prognosis[24,45]. Therefore, it is necessary to study the function of CASC19 in CRC. In our study, it was observed that the expression of CASC19 w as greatly raised in both CRC cell lines and tissue samples. This result is consistent with previous results, which suggests that CASC19 may be an important lncRNA that regulates CRC heterogeneity. Given that the cellular location of CASC19 w as related to lncRNA function, w e examined the subcellular distribution of CASC19 using cellular fractionation. Cellular localization show ed that CASC19 w as mostly located in the cellular plasma, indicating that CASC19 may function to sponge miRNA.
Heterogeneity is a key property that is responsible for malignant processes such as initiation, migration, and metastasis of cancer. One example is the lncRNA HOTAIR that may promote the progression of a variety of cancers by affecting the proliferation,migration, and invasion of various cancer cells[46]. Knockdown of the lncRNA LCPAT1 inhibits autophagy in lung cancer[47]. The lncRNA PVT1 may promote ovarian cancer progression[48]. These data suggest that lncRNAs are emerging as crucial cancer regulators. Correlation analysis show ed that high expression of CASC19 w as associated with CRC metastasis. Thus, we speculated that CASC19 may be correlated with CRC progression. As we expected, in vitro experiments showed that CASC19 promoted CRC progression.
It has been demonstrated that lncRNAs may function as ceRNAs by sponging miRNAs[49]. For example, HOTAIR promotes renal cell carcinoma progression via absorbing miR-138[50]. The lncRNA SNHG7 sponges miR-216b to upregulate GALNT1,leading ultimately to the promotion of CRC cell proliferation and liver metastasis[51].Previously, miR-140-5p w as observed to be expressed abnormally in several malignancies including CRC[52], breast cancer[53], ovarian cancer[54], glioma[55], and gastric cancer[56]. Similar to previous studies, our study revealed that CASC19 may promote CRC by targeting miR-140-5p. The current investigations revealed that CASC19 directly targets miR-140-5p. Imp ortantly, there w as an end ogenous interaction betw een these tw o molecules in CRC cells. Moreover, inhibiting proliferation and invasion and promoting apoptosis of cancer cells are known to involve miR-140-5p[55,57]. These studies suggest that miR-140-5p may play a tumor suppressive role in different tumors. However, literature is scarce regarding its role in CRC. Our results demonstrated that miR-140-5p was markedly reduced in CRC cell lines and tissue samples. In addition, miR-140-5p reversed the function of CASC19 in CRC cell lines. These results suggest that CASC19 sponging of miR-140-5p may be a mechanism regulating the heterogeneity of CRC. Given that miRNAs are know n to simultaneously control the exp ression of a broad range of genes, our w orking hyp othesis w as that miR-140-5p may target d ifferent genes in different cell lines,resulting in varying phenotypes. Therefore, we examined the target genes predicted and found out that CEMIP may be a target gene of miR-140-5p in CRC.
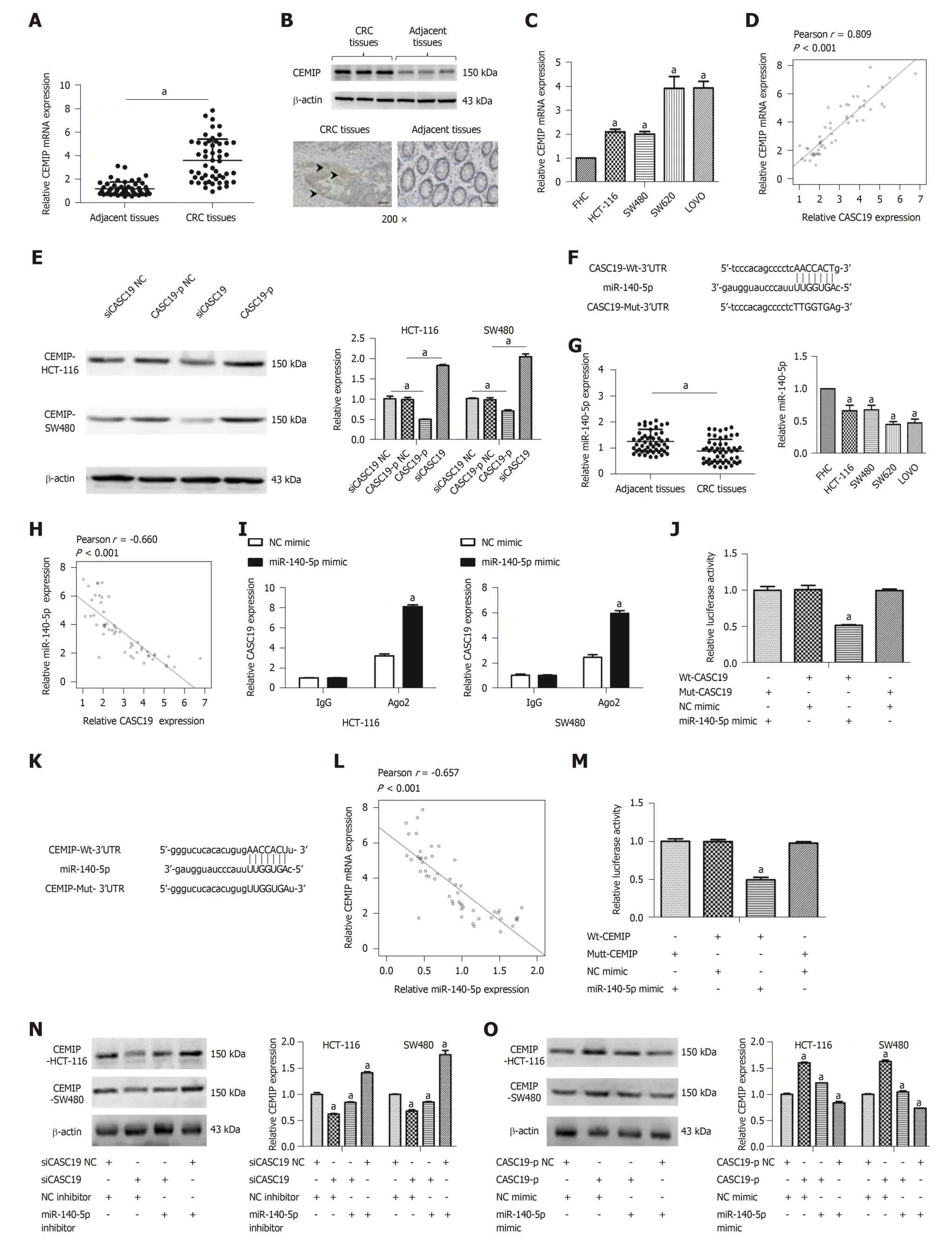
Figure 4 Cancer susceptibility 19 acts as a competing endogenous RNA by sponging miR-140-5p to regulate cell migration inducing hyaluronidase 1 expression. A: Differential mRNA expression of cell migration inducing hyaluronidase 1 (CEMIP) in colorectal cancer (CRC) tissues and adjacent healthy colonic mucosa tissues was determined by quantitative real-time PCR (qRT-PCR). B: Differential protein expression of CEMIP in CRC tissues and adjacent healthy colonic mucosa tissues was analyzed using Western blot and immunochemistry. C: Differential mRNA expression of CEMIP in CRC cell lines and FHC cells was determined by qRT-PCR. D: Cancer susceptibility 19 (CASC19) and CEMIP correlated positively in CRC tissues, based on Pearson's correlation curve. E: Protein expression of CEMIP in SW480 and HCT-116 cells was determined by Western blot after transfection with CASC19-p or siCASC19. The grayscale value of protein was measured with Image J. F: Predicted binding sites of miR-140-5p in the CASC19 sequence. G: Differential expression of miR-140-5p in CRC tissues and adjacent healthy colonic mucosa tissues, CRC cell lines, and FHC cells was determined by qRT-PCR. H: CASC19 and miR-140-5p correlated negatively in CRC tissues, as determined by Pearson's correlation curve. I: RNA-IP was performed in SW480 and HCT-116 cells and CASC19 expression was determined by qRT-PCR. J:Luciferase reporter assay verified miR-140-5p to be a direct target of CASC19. K: Predicted binding sites of miR-140-5p in the CEMIP mRNA sequence. L: CEMIP and miR-140-5p correlated negatively in CRC tissues, based on Pearson's correlation curve. M: Luciferase reporter assay verified CEMIP to be a direct target of miR-140-5p. N: CEMIP expression in SW480 and HCT-116 cells transfected with miR-140-5p mimic or siCASC19 was determined by Western blot and qRT-PCR. O: The expression of CEMIP in SW480 and HCT-116 cells transfected with miR-140-5p inhibitor or CASC19-p was determined by Western blot and qRT-PCR. Data are reported as the mean ± SD. a P < 0.05. CASC19: Cancer susceptibility 19; CRC: Colorectal cancer; CEMIP: Cell migration inducing hyaluronidase 1.
Tumor malignancy is driven by its almost limitless capability for proliferation and metastasis. CEMIP w as initially know n as an enzyme to depolymerize hyaluronic acid. Further stud ies reveal CEMIP overexpression to be a common phenomenon in several tumors. For instance, CEMIP drives breast cancer progression and predicts poor p rognosis in pancreatic ductal adenocarcinoma[31,32]. In CRC, tumor metastasis and invasion are able to be suppressed through the miR-216a-mediated attenuation of CEMIP[58]. CEMIP therefore may be associated with the progression of CRC. Previous studies have demonstrated that CEMIP may interact with the glycogen phosphorylase kinase β-subunit (PHKB) to promote cancer cell survival by breakdown of glycogen[59].In CRC tissues, the tumor cells are exp osed to a nutrient d ep rivation and hypoxia environment[60]. Energy deprivation is one of the main courses of cell death. It has been demonstrated that glycogen accumulation contributes to cell survival w hen cancer cells expose to hypoxia[61-63]. At the same time, glycogen lysis is an important process for cell survival. CEMIP overexpressing tumor cells have relatively low glycogen content, w hile the survival rate of these cells is relatively high. CEMIP may play a vital role in CRC survival and metastasis. Many differing miR-140-5p targets,such as YES1[56], Wnt1[64], and VEGFA[65], have been documented to be associated w ith cell invasion and migration. In our study, w e observed that CASC19 functions as a ceRNA, controlling the availability of miR-140-5p that can be acted on by the CEMIP gene. The current investigations focused on CEMIP, which was found to have strong associations with CASC19 and cancer cell survival. Our results strongly suggest that CASC19 may regulate CEMIP expression via sponging of miR-140-5p.
In conclusion, CASC19 overexp ression augmented CRC cell proliferation and metastasizing abilities in vitro. Our study is the first to report a novel mechanism for CASC19 in CRC. CASC19 exerted its effects partially through the CASC19/miR-140-5p/CEMIP axis. Investigation of this CASC19/miR-140-5p/CEMIP pathway allows a deeper understanding of the pathogenesis of CRC, w ith the members of this pathway serving as useful targets for future p reventive and therapeutic innovations in the management of CRC.
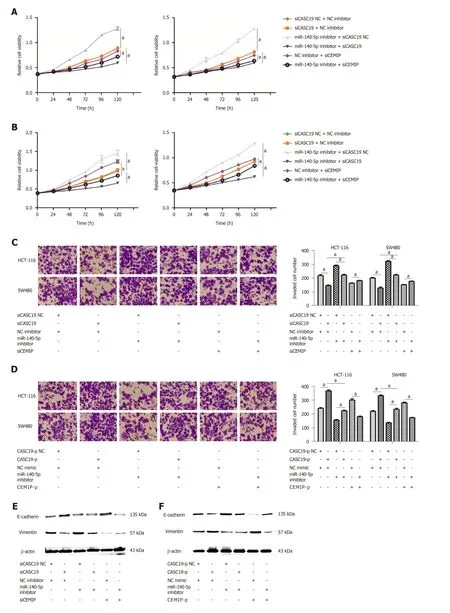
Figure 5 MiR-140-5p suppresses cancer susceptibility 19-induced enhancement of colorectal cancer cell proliferation and metastasis. A: Growth curves of SW480 and HCT-116 cell lines after being transfected by either miR-140-5p inhibitor or siCASC19 were determined by the Cell Counting Kit-8 (CCK-8) assay. B:Growth curves of SW480 and HCT-116 cell lines after being transfected by either miR-140-5p mimic or CASC19-p were analyzed by the CCK-8 assay. C: The number of invading HCT-116 and SW480 cells was determined by transwell assay after being transfected by either miR-140-5p inhibitor or siCASC19. Scale bars = 20 μm. D:The number of invading HCT-116 and SW480 cells was determined by transwell assay after being transfected with either miR-140-5p mimic or CASC19-p. Scale bars= 20 μm. E: The expression of E-cadherin and vimentin in CRC cells was determined by Western blot after being transfected by either miR-140-5p inhibitor orsiCASC19. F: The expression of E-cadherin and vimentin in CRC cells was determined by Western blot after being transfected with either miR-140-5p mimic or CASC19-p. Data are reported as the mean ± SD. a P < 0.05. CASC19: Cancer susceptibility; CRC: Colorectal cancer; CEMIP: Cell migration inducing hyaluronidase 1.
ARTICLE HIGHLIGHTS
Research background
There is increasing evidence that long non-coding RNAs (lncRNAs) play an important role in tumor progression. The lncRNA cancer susceptibility 19 (CASC19) is highly expressed in colorectal cancer (CRC) tissues, and may be associated w ith colon cancer progression. How ever,there has been no experimental evidence regarding the function of CASC19 in CRC.
Research motivation
Investigation of the functions of the lncRNA CASC19 may suggest p otential molecular mechanisms of CRC carcinogenesis and progression, and may further offer the potential for future preventive and therapeutic innovations in the management of CRC.
Research objectives
We measured the lncRNA CASC19 expression in CRC tissues and pair-matched adjacent nontumor tissues, and investigated biological functions and the possible molecular mechanisms of CASC19 in CRC.
Research methods
We detected CASC19 expression in CRC tissues and pair-matched adjacent non-tumor tissues using quantitative real-time PCR. The biological behavior of CASC19 in vitro was then assessed by overexpression and knockdow n of the expression of CASC19. In further molecular mechanism studies, we confirmed the possible molecular mechanisms by which CASC19 plays a role by overexpression and knockdown of the expression of downstream molecules of CASC19.
Research results
The lncRNA CASC19 was upregulated in CRC tissue, which was related to tumor stage and metastasis. The lncRNA CASC19 had a role of promoting proliferation, metastasizing ability,and epithelial-mesenchymal transition, and inhibiting apoptosis by regulating microRNA 140-5p/cell migration inducing hyaluronidase 1 (miR-140-5p/CEMIP) expression in CRC cells.
Research conclusions
The lncRNA CASC 19 is overexpressed in CRC tissues and cell lines, w hich could promote CRC progression through regulating miR-140-5p/CEMIP.
Research perspectives
Members of the CASC19/miR-140-5p/CEMIP axis may serve as useful targets for future preventive and therapeutic innovations in the management of CRC.
杂志排行

World Journal of Gastroenterology的其它文章
- Precision surgical approach with lymph-node dissection in early gastric cancer
- Update on hepatocellular carcinoma: Pathologists' review
- Application of artificial intelligence in gastroenterology
- Clinical significance of programmed death 1/programmed death ligand 1 pathway in gastric neuroendocrine carcinomas
- Seven-senescence-associated gene signature predicts overall survival for Asian patients with hepatocellular carcinoma
- Chronic functional constipation is strongly linked to vitamin D deficiency