生产类群、性别及包装方式对冷藏牦牛肉肌浆蛋白氧化的影响
2019-05-05李思宁唐善虎
李思宁,唐善虎*
(西南民族大学生命科学与技术学院,四川 成都 610041)
在肉品的加工贮藏过程中,脂肪和蛋白质氧化是导致肉品质劣变的一个重要原因[1]。由于蛋白质在细胞成分中含量高并能与活性氧高速反应,因此蛋白质是发生氧化的主要成分[2]。肌肉中的蛋白质可以通过活性氧直接修饰,也可通过脂肪氧化产物或HClO、H2O2等非自由基修饰。蛋白氧化会导致分子结构和完整性发生显著变化,改变氨基酸侧链性质、生成羰基衍生物、产生蛋白质聚合物等[3-4],由此影响肉制品加工的营养和感官特性[5-8]。
牦牛是生存在高海拔地区的唯一牛种,由于长期的封闭繁衍和对环境生态的适应性,不同生产区域牦牛的生产性能和肉质出现了差异,形成了不同生产类群,但还不足构成具有显著外貌和生产性能差异的品系。牦牛肉的肌浆蛋白约占牦牛肉总蛋白质量的30%~35%,大部分生化代谢及氧化还原反应都在肌浆蛋白中发生,与糖酵解相关的酶、与肉色有密切关系的肌红蛋白也都存在于肌浆蛋白质内[10-11]。肌红蛋白和参与代谢的酶类的氧化,会进一步导致肉颜色及肉质的劣变[12]。
现阶段,有关肉类蛋白质的氧化研究主要集中在肉中肌原纤维蛋白在氧化剂或模拟氧化体系[13-14]、不同包装[15-17]、不同加工处理[18-20]条件下结构和功能性的变化;而有关肌浆蛋白氧化对肉类功能特性方面的影响研究报道较少。马纪兵等[21]研究了冷藏过程中非真空和真空包装牦牛肉蛋白质条带的变化情况,发现随着冷藏时间的延长,这两种包装方式的牦牛肉肌浆蛋白表现出不同程度的降解。Sher[22]研究了反复冻融对鸡胸肉品质的影响,发现冻融次数的增加导致蛋白氧化加剧,表现为羰基含量升高,巯基含量降低,肌浆蛋白降解。Marcos等[23]研究发现200 MPa以上的压力会诱导肌浆蛋白溶解度和持水力降低、色泽改变及肌浆蛋白组成变化。Tokur等[18]发现在4 ℃冷藏条件下,2 岁龄细唇鲻鱼肌浆蛋白羰基含量下降,4 岁龄和6 岁龄的羰基含量无显著变化;肌浆蛋白条带的变化与细唇鲻鱼的年龄及β-巯基乙醇是否存在有关,主要表现在100、50 kDa及37 kDa附近蛋白条带的改变。目前鲜见有关牦牛生产类群和性别影响牦牛肉低温冷藏过程肌浆蛋白氧化特性的研究报道。本研究选取四川甘孜州4 个不同生产类群的公、母牦牛的背长肌,经普通包装或真空包装处理后,在4 ℃下冷藏,通过测定不同冷藏时间下肌浆蛋白质氧化、紫外吸收光谱及电泳指标,探讨牦牛肉中肌浆蛋白质的氧化稳定性及蛋白质降解情况,为冷藏牦牛肉品质的控制提供参考依据。
1 材料与方法
1.1 材料与试剂
选取自然放牧状态下,甘孜州色达、新龙、稻城及理塘4 个生产类群的健康无病的3 岁龄公牦牛各3 头、母牦牛各3 头,共计24 头牦牛集中屠宰,采集背长肌,装入洁净保鲜袋中密封,放入-18 ℃冰箱急冻2 h,在4 ℃条件下运回实验室。
乙二胺四乙酸、2,4-二硝基苯肼、5,5’-二硫代双(2-硝基苯甲酸)、磷酸氢二钠、磷酸二氢钠、五水硫酸铜、酒石酸钾钠、硫氰化钾、溴酚蓝、β-巯基乙醇、尿素(均为分析纯) 成都市科龙化工试剂厂;丙烯酰胺、十二烷基硫酸钠(sodium dodecyl sulfate,SDS)、三羟甲基氨基甲烷、N,N’-甲叉双丙烯酰胺、N,N,N’,N’-四甲基乙二胺、过硫酸铵、甘氨酸、考马斯亮蓝R-250、标准牛血清白蛋白(均为分析纯) 美国Sigma公司;聚丙烯酰胺凝胶电泳(polyacrylamide gel electrophoresis,PAGE)标准蛋白(11~245 kDa) 上海生化试剂公司。
1.2 仪器与设备
5804离心机 德国Eppendorf公司;紫外-可见分光光度计 上海翱艺公司;FSH-2A高速组织匀浆机 金坛市华城海龙实验仪器厂;HH-6恒温水浴锅 国华电器有限公司;Mini电泳槽、Versa Doc 1000凝胶成像系统 美国Bio-Rad公司;DYY-12型电泳仪 北京市六一仪器厂。
1.3 方法
1.3.1 实验设计
甘孜州色达(SD)、新龙(XL)、稻城(DC)及理塘(LT)4 个不同生产类群的公(male,M)、母(female,F)牦牛肉切割成厚度1 cm、每份质量50 g的肉块,经过普通包装(2丝聚乙烯复合包装膜)或真空包装(10 cm×20 cm的双面20丝聚乙烯复合真空包装袋)处理后,在4 ℃条件下冷藏。从第0天开始,每隔3 d取一次样,肉样变质(色泽及气味改变)作为取样的终点。所有样品提取肌浆蛋白后,测定蛋白氧化指标、紫外吸收光谱及SDS-PAGE。
1.3.2 肌浆蛋白的提取及测定
参考Tokur等[18]的方法,并略作修改。取牦牛肉5 g,剔除脂肪和结缔组织,切成2 mm×2 mm×2 mm大小的肉粒,添加5 倍体积的50 mmol/L磷酸盐缓冲液(pH 7.0),13 000 r/min匀浆60 s,5 000 ×g、4 ℃离心20 min,上清液经4号滤纸过滤,沉淀再加提取液提取,重复上述操作,合并两次离心的上清液,即为肌浆蛋白。采用双缩脲法测定蛋白含量[24]。
1.3.3 羰基含量的测定
羰基含量的测定采用Levine等[25]的方法,并略作修改。1 mL 5 mg/mL的蛋白溶液与1 mL 10 mmol/L的2,4-二硝基苯肼(用2 mol/L盐酸溶解)混匀,以2 mol/L盐酸溶液作对照;将反应体系置于黑暗中反应1 h,每隔10 min漩涡1 次;反应完毕后,加入1 mL、质量分数20%的三氯乙酸溶液,摇匀后12 000 ×g、4 ℃离心15 min,弃上清液;沉淀用1 mL乙醇-乙酸乙酯混合物(1∶1,V/V)洗涤3 次,再溶于3 mL 6 mol/L的盐酸胍溶液(37 ℃水浴15 min);12 000 ×g离心15 min除去不溶物质,取上清液,在波长370 nm波长处测定吸光度(A)。羰基含量按公式(1)计算。
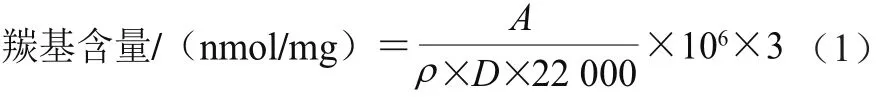
式中:ρ为蛋白质量浓度/(mg/mL);D为比色光径(1 cm);22 000为摩尔吸光系数/(L/(mol·cm));3为稀释倍数。
1.3.4 总巯基含量的测定
梁诚趁着夜色离开家,来到一间公寓,拿钥匙开门。一个女人坐在沙发上看着电视吃着薯片。看见开门的梁诚,女人开心地叫了一声:“老公,你来啦!”
总巯基的测定参考Ellman[26]的方法。取1.5 mL 5 mg/mL蛋白溶液与10.0 mL Tris-甘氨酸缓冲液(含8 mol/L尿素、0.086 mol/L Tris、0.09 mol/L甘氨酸、4 mmol/L乙二胺四乙酸,pH 8.0)混匀;将以上处理样品分别加50 μL 5,5’-二硫代双(2-硝基苯甲酸)(用0.1 mol/L的Tris-盐酸溶解,pH 7.6)试剂,剧烈振荡后在25 ℃条件下水浴1 h,12 000 ×g离心10 min,同时以0.1 mol/L Tris-盐酸作为对照,取上清液在412 nm波长处测定吸光度(A)。蛋白中巯基含量用公式(2)计算。
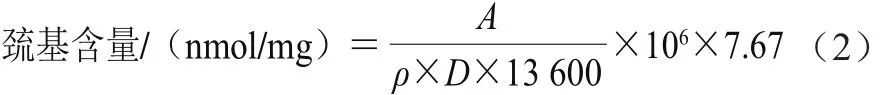
式中:ρ为蛋白质量浓度/(mg/mL);13 600为摩尔吸光系数/(L/(mol·cm));D为比色光径(1 cm);7.67为稀释倍数。
1.3.5 表面疏水性的测定
表面疏水性的测定参考Chelh等[27]的方法。取1 mL 5 mg/mL蛋白溶液,加入200 μL 1 mg/mL溴酚蓝溶液,振荡混匀10 min,7 000×g离心15 min,上清液稀释10 倍后,在595 nm波长处测定吸光度(A)。空白样为未加样品的磷酸盐缓冲液。以溴酚蓝结合量表示表面疏水性,按公式(3)计算。
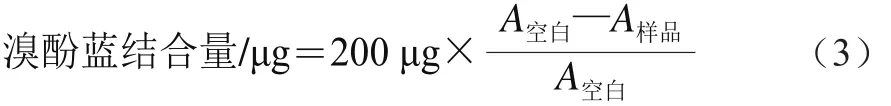
1.3.6 紫外吸收光谱测定
将肌浆蛋白稀释到0.4 mg/L,在200~300 nm波长处进行紫外扫描,记录吸收光谱。
1.3.7 牦牛肉肌浆蛋白SDS-PAGE
牦牛肉肌浆蛋白的SDS-PAGE参考Laemmli[28]的方法。1 mg/mL待测蛋白质和5×样品缓冲液以体积比3∶1混合,沸水浴5 min,冷却后上样,上样体积为10 μL。分离胶质量分数12%、浓缩胶质量分数4%。电泳后用质量分数为0.2%的考马斯亮蓝R-250染色液染色,用含体积分数7.5%甲醇和7.5%醋酸的脱色液脱色至背景清晰。使用凝胶成像仪拍照,分析蛋白聚集与降解变化。
1.4 数据处理
2 结果与分析
2.1 冷藏过程中牦牛肉肌浆蛋白羰基含量的变化
羰基的产生作为蛋白氧化的标志之一,是由易受自由基攻击的带有—NH—或者—NH2的氨基酸残基以及肽键的断裂生成。
由表1可知,普通包装组肌浆蛋白羰基在冷藏前3 d迅速增加,第3天显著高于第0天的羰基含量(P<0.05);至第6天,羰基含量显著下降(P<0.05);冷藏第9天,羰基含量达到最大值。肉类产品在低温保藏过程中,羰基含量会上升[29-30],但是随冷藏时间的延长,羰基含量可能会下降,如通过Strecker反应降解[31]。有研究报道,新生成的羰基会攻击蛋白质中的亲核物质,发生羰氨缩合反应,导致羰基含量降低[32]。在普通包装组中,生产类群之间肌浆蛋白羰基含量差异显著(P<0.05);同一生产类群的公、母牦牛肉之间肌浆蛋白羰基含量差异均不显著(P>0.05)。
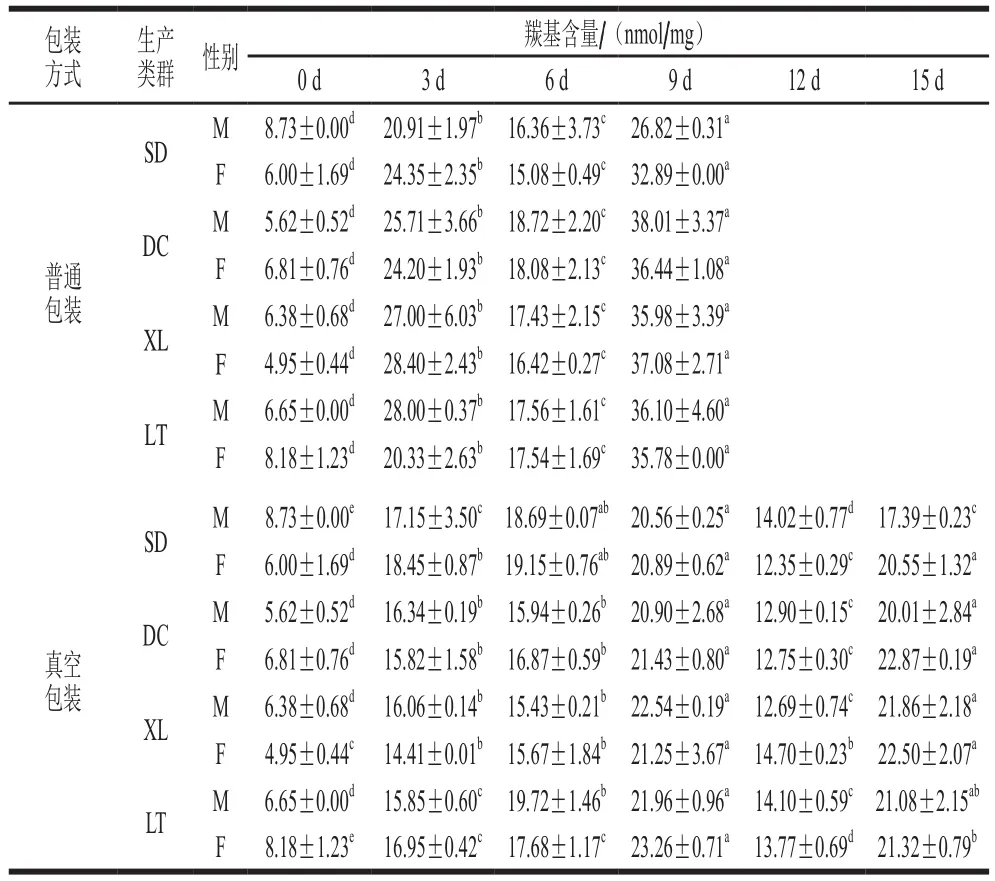
表1 不同冷藏时间的牦牛肉肌浆蛋白羰基含量Table 1 Carbonyl contents of sarcoplasmic proteins in yak meat at different storage times
冷藏0~9 d,真空包装组肌浆蛋白羰基含量显著增加(P<0.05);除SD-M和LT-M组外,其余组在第3、6天羰基含量差异不显著(P>0.05)。第12天,羰基含量下降,表明羰基可能发生了降解或聚合或被进一步氧化成了其他的产物;至第15天时,羰基含量又增加;除SD-M和LT-F外,其余组羰基含量在第15天均与第9天无显著差异(P>0.05),说明蛋白氧化加剧。在真空包装组中,生产类群之间肌浆蛋白羰基含量差异显著(P<0.05);同一生产类群的公、母牦牛肉之间肌浆蛋白羰基含量差异均不显著(P>0.05)。
2.2 冷藏过程牦牛肉肌浆蛋白总巯基含量的变化
半胱氨酸残基是蛋白质中最容易受到攻击的氨基酸残基,总巯基含量可以用来评价半胱氨酸残基的氧化状态。肌肉蛋白中的巯基基团发生氧化,生成二硫键、次磺酸、亚磺酸和磺酸[33]。Eaton[34]研究发现,在不同的氧化环境下,蛋白质巯基可以被可逆地氧化为二硫键和次磺酸(早期氧化过程),也可以被不可逆地氧化为亚磺酸和磺酸。
由表2可知,随冷藏时间的延长,普通包装组肌浆蛋白总巯基含量在0~3 d无显著变化(P>0.05),说明冷藏前期,包装袋内的活性氧使部分巯基被可逆地氧化成了二硫键和次磺酸;3 d后,总巯基含量显著下降(P<0.05),在此阶段巯基被氧化的速度大于被还原的速度,生成了不可逆的氧化状态,形成了非二硫键的含硫化合物。在普通包装组中,生产类群之间肌浆蛋白总巯基含量差异显著(P<0.05)。同一生产类群的公、母牦牛肉之间肌浆蛋白总巯基含量差异均不显著(P>0.05)。
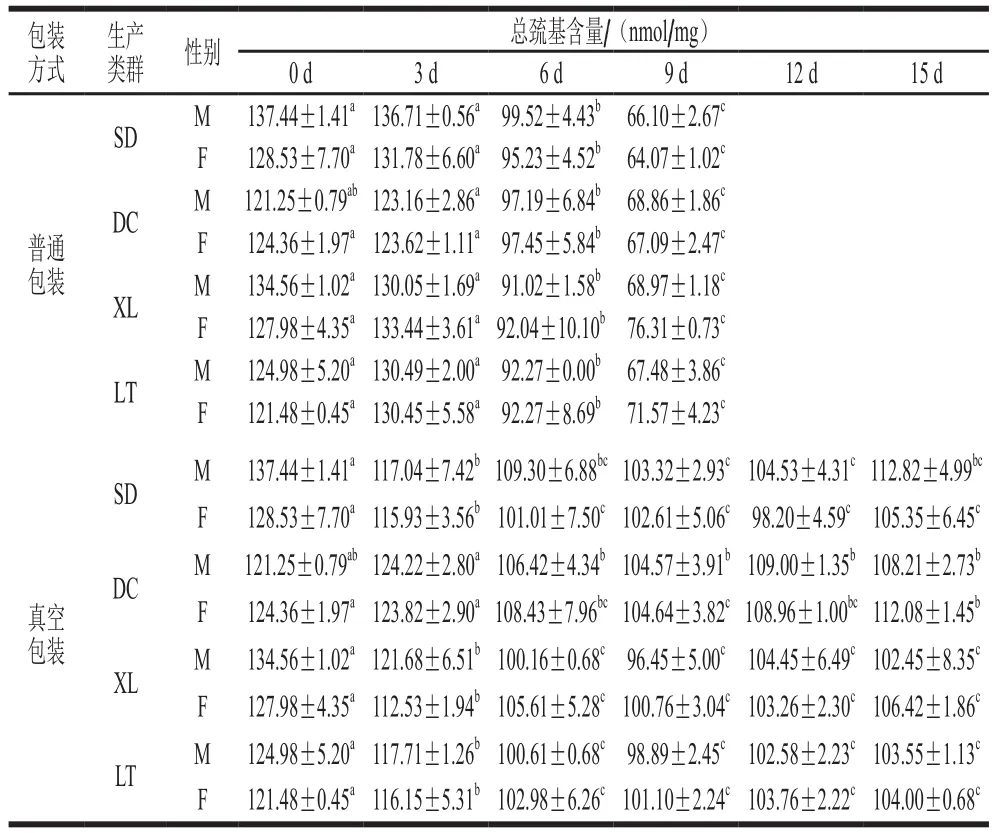
表2 不同冷藏时间的牦牛肉肌浆蛋白总巯基含量Table 2 Sulfhydryl contents of sarcoplasmic proteins in yak meat at different storage times
冷藏0~6 d,真空包装组肌浆蛋白总巯基含量下降(P<0.05),包装内残留的氧使暴露在蛋白质表面的半胱氨酸发生了氧化,导致巯基含量减少。6~15 d,总巯基含量基本趋于稳定(P>0.05),可能是包装内的氧分压较低,巯基被氧化速度约等于被还原的速度,表现出巯基含量没有发生大的改变。在真空包装组中,生产类群之间肌浆蛋白总巯基差异显著(P<0.05);同一生产类群的公、母牦牛肉之间肌浆蛋白总巯基含量差异均不显著(P>0.05)。
2.3 冷藏过程牦牛肉肌浆蛋白表面疏水性的变化
表面疏水性反映的是蛋白质表面疏水性氨基酸的相对含量,疏水性的改变与蛋白质结构变化密切相关。
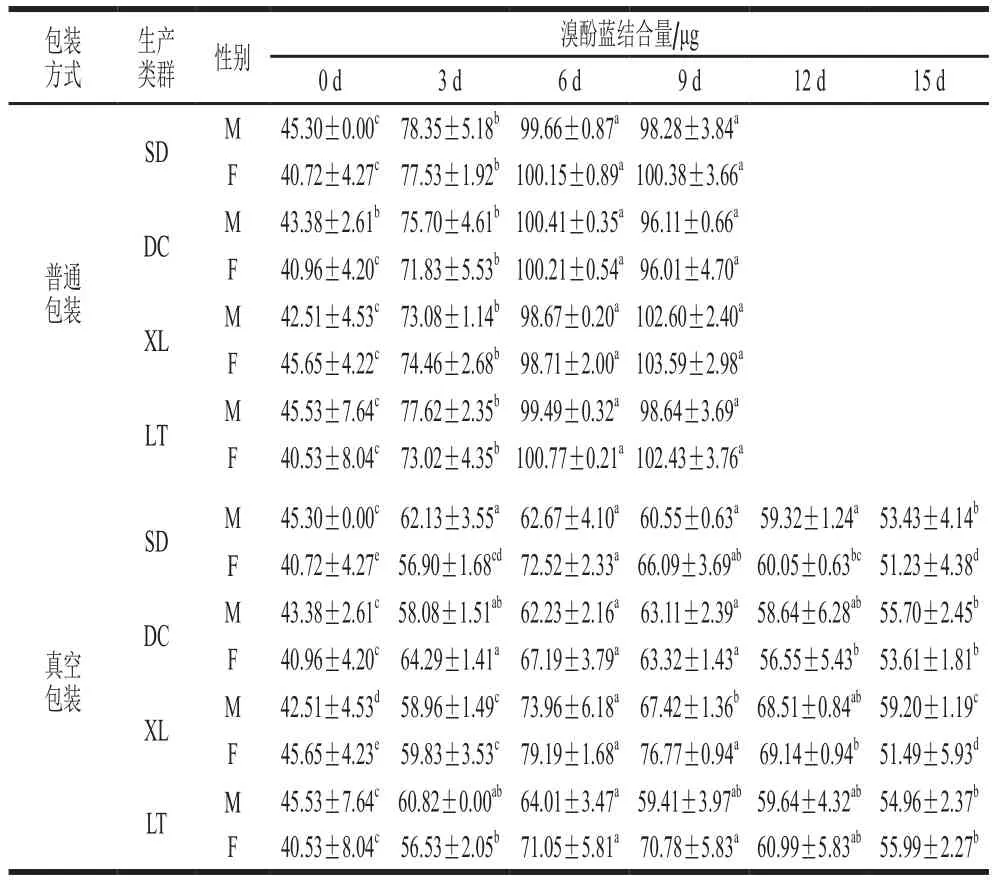
表3 不同冷藏时间的牦牛肉肌浆蛋白表面疏水性Table 3 Surface hydrophobicity of sarcoplasmic proteins in yak meat at different storage times
由表3可知,随冷藏时间延长,普通包装组肌浆蛋白疏水性呈上升趋势。氧化引起肌浆蛋白构象改变后,肌浆蛋白中更多的疏水性氨基酸残基暴露,导致表面疏水性增加。与第6天相比,第9天肌浆蛋白表面疏水性没有显著增加,可能是随着氧化继续进行,蛋白表面的疏水基团由于疏水相互作用发生了聚集[35]。在普通包装组中,生产类群之间肌浆蛋白疏水性差异显著(P<0.05);同一生产类群的公、母牦牛肉之间肌浆蛋白疏水性差异均不显著(P>0.05)。
真空包装组肌浆蛋白疏水性在0~6 d呈增加的趋势,随后疏水性开始下降。6 d后肌浆蛋白表面疏水性开始缓慢下降,可能是由于暴露的疏水性氨基酸残基发生聚集或形成了其他疏水性物质。在真空包装组中,生产类群之间肌浆蛋白疏水性差异显著(P<0.05)。同一生产类群的公、母牦牛肉之间肌浆蛋白疏水性差异均不显著(P>0.05)。
2.4 冷藏过程牦牛肉肌浆蛋白紫外吸收光谱
二阶导数紫外分光光度法是一种用于测定几种化合物定性和定量的分析方法,如鉴别氨基酸、揭示蛋白质结构的变化[36]。
由图1A可知,冷藏第0天,普通包装组肌浆蛋白在245 nm及285 nm波长处各有1 个波峰,在235、280 nm及290 nm波长处各有1 个波谷;280 nm和290 nm波长处的吸收峰分别归属于酪氨酸和色氨酸残基,285 nm波长处的波峰归属于色氨酸和酪氨酸的共同作用[37]。由图1B可知,与第0 天相比,第3天的二阶导数紫外光谱在280~290 nm波长处的吸收峰均减弱,这可能是酪氨酸、色氨酸被氧化后含量降低造成的;245 nm波长处明显的吸收峰表明肌浆蛋白中K带的存在,即存在共轭的不饱和键。由图1C、D可知,在第6、9天,样品均在295 nm波长处有明显的正吸收,其余波段有很低强度的紫外吸收,第9天的吸收峰强度大于第6天的。在整个冷藏过程中,肌浆蛋白构象发生了很大程度的变化,尤其在冷藏第3天改变最明显。在氧自由基的攻击下,芳香族氨基酸、带有—NH—或者—NH2的氨基酸被氧化及疏水性氨基酸暴露都会导致肌浆蛋白构象发生变化。而相同冷藏时间,普通包装组的不同生产类群及性别的牦牛肉肌浆蛋白紫外吸收谱图波形一致。
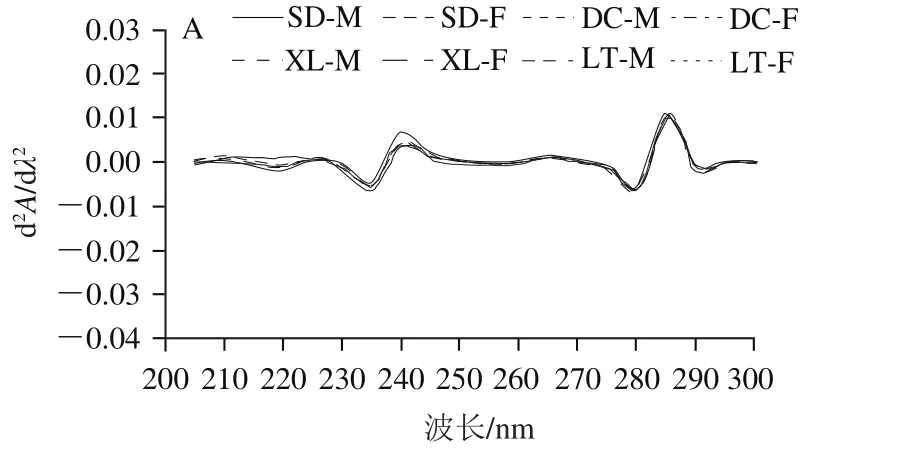
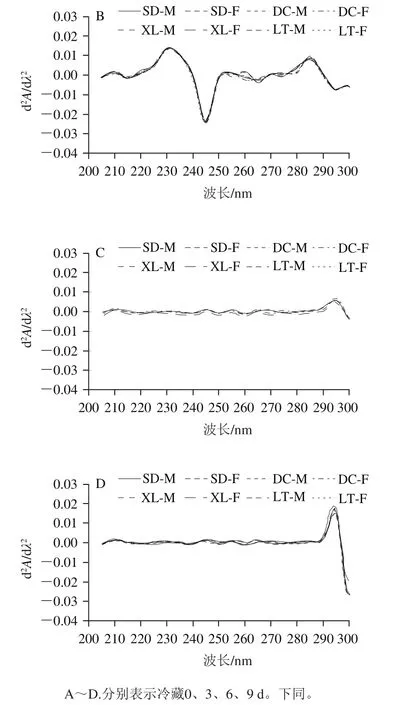
图1 不同冷藏时间的普通包装组肌浆蛋白紫外二阶导数紫外光谱图Fig. 1 Second-order derivative UV spectra of sarcoplasmic proteins in air-packaged yak meat at different storage times
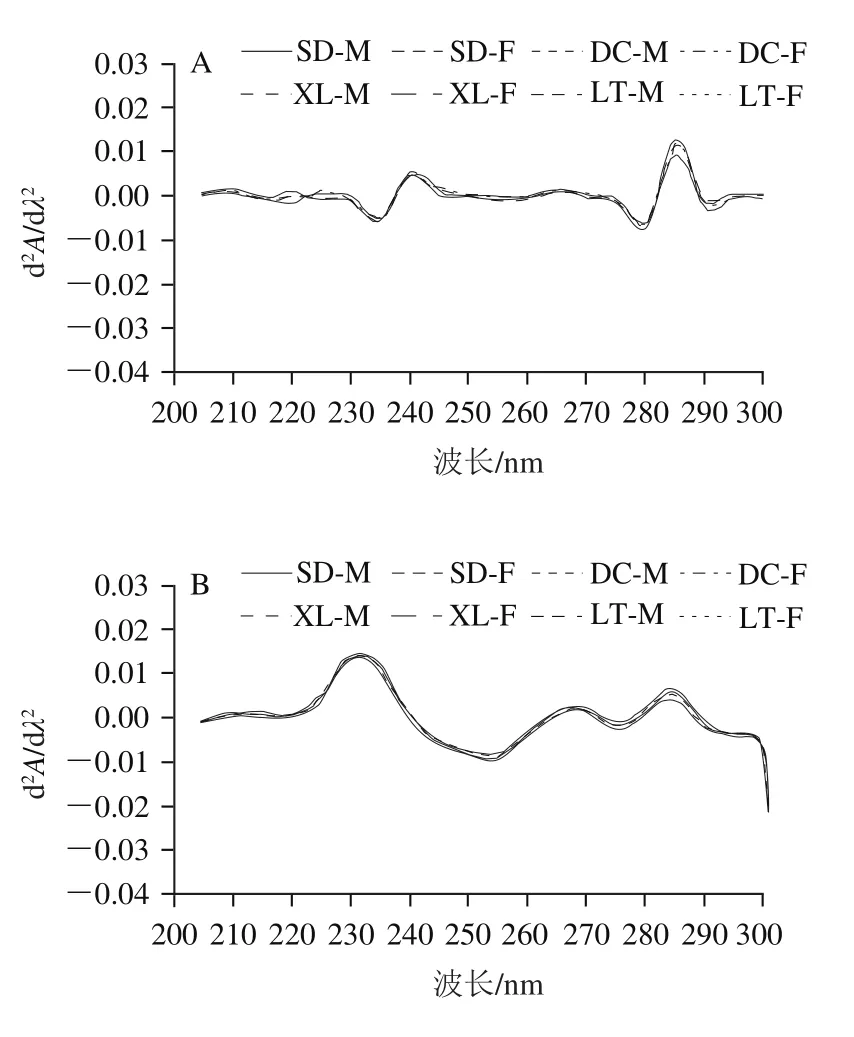

图2 不同冷藏时间的真空包装组肌浆蛋白紫外二阶导数紫外光谱图Fig. 2 Second-order derivative UV spectra of sarcoplasmic protein in vacuum-packaged yak meat at different storage times
与第0天相比,第3天的二阶导数紫外光谱在280~290 nm波长处的吸收峰均减弱(图2A、B)。由图2B可知,真空包装组肌浆蛋白在贮藏第3天的紫外波谱与普通包装组第3天相似,区别在于245 nm处的波谷红移到了255 nm。由图2C~E可知,冷藏第6、9、12天的波谱与普通包装组第6天基本一致。由图2F可知,与第12天相比,第15天肌浆蛋白的波谱发生了改变,295 nm处的波峰蓝移到了290 nm处,且吸收峰明显增强,表明蛋白质氧化程度加剧。所处的微环境疏水时,特征谱峰会发生蓝移[38]。冷藏6~12 d,肌浆蛋白的紫外吸收谱图无明显改变,说明真空包装抑制了蛋白质氧化。在整个冷藏过程中,肌浆蛋白构象在冷藏第3天变化最明显。相同冷藏时间,真空包装组的不同生产类群及性别的牦牛肉肌浆蛋白紫外吸收谱图波形一致。
2.5 冷藏过程牦牛肉肌浆蛋白SDS-PAGE
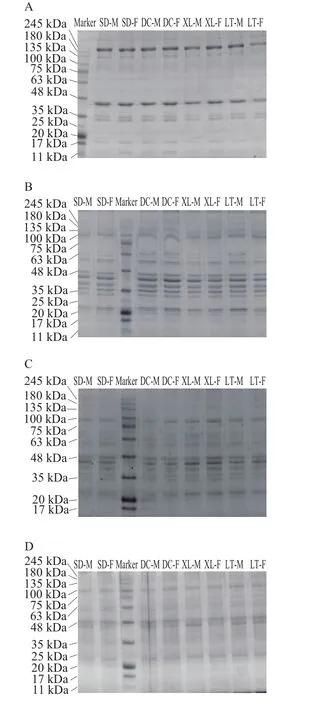
图3 不同冷藏时间的普通包装组肌浆蛋白SDS-PAGE图Fig. 3 SDS-PAGE patterns of sarcoplasmic proteins in air-packaged yak meat at different storage times
由图3A~D可知,随冷藏时间的延长,普通包装组的牦牛肉肌浆蛋白分子质量逐渐降低,所有蛋白条带都发生了一定程度的弱化。冷藏第0天,245 kDa附近的蛋白条带是区别于其他冷藏时间的明显条带。第3天,245 kDa附近的蛋白条带消失,降解成了20~75 kDa及135~180 kDa的小分子蛋白质。第6天和第9天电泳图中100~135 kDa的蛋白条带明显弱化或消失,其他条带也发生了弱化;在第9天,35~48 kDa的条带基本消失。在整个冷藏过程中,100 kDa附近的两条条带一直存在,可能为糖原磷酸化酶(97 kDa)和β-半乳糖苷酶(116 kDa)[39-40]。肌红蛋白(17 kDa附近)在冷藏0~3 d时存在,第3天时条带已发生弱化,6 d后全部消失。Grune等[41]发现氧化的增加可以通过蛋白酶促进蛋白质降解。Ryu等[42]发现肌浆蛋白条带的弱化伴随着析出的肌浆蛋白结合到肌原纤维蛋白上,因此肉色由红色转变为苍白色。相同冷藏时间,普通包装组的不同生产类群及性别的牦牛肉肌浆蛋白SDS-PAGE图谱中蛋白条带一致。
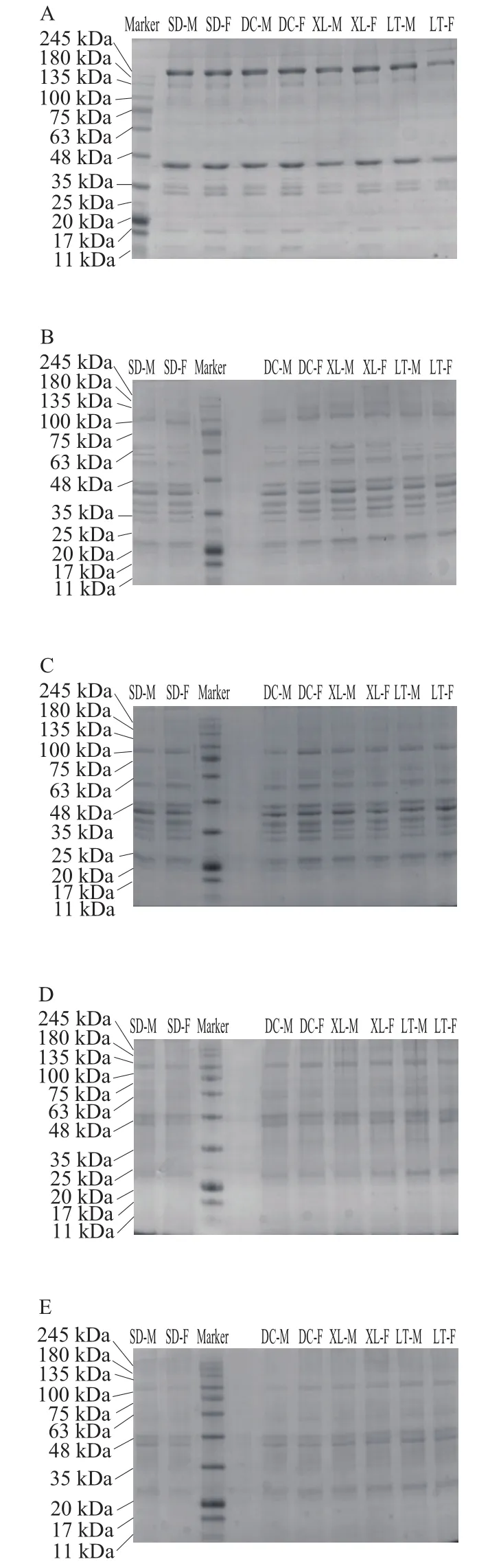
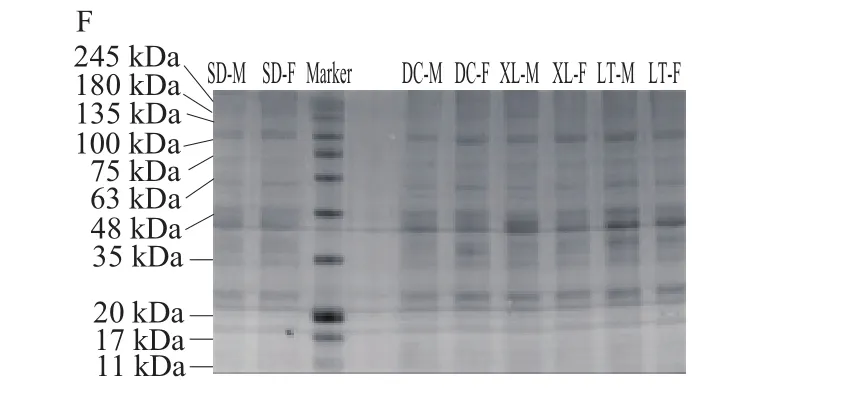
由图4A~F可知,随冷藏时间的延长,真空包装组牦牛肉的肌浆蛋白分子质量降低,所有蛋白条带都发生了一定程度的弱化。与第0天相比,冷藏第3天真空包装组的肌浆蛋白245 kDa附近的蛋白条带消失,电泳图谱与相同冷藏时间的普通包装组一致。冷藏第6天,肌浆蛋白电泳图中100~135 kDa的蛋白条带消失,降解成了分子质量在20 kDa附近的小分子蛋白质,且63 kDa附近的条带发生了弱化。第9、12、15天的肌浆蛋白电泳图基本一致,与其他时间的图谱相比,所有条带均发生了弱化。相同冷藏时间,真空包装组的不同生产类群及性别的牦牛肉肌浆蛋白电泳图谱中蛋白条带一致。本实验牦牛肉的肌浆蛋白条带的分布规律和分子质量与马纪兵等[21]报道的甘南牦牛肉有较大差异,这可能是牦牛品种差异造成的。
3 结 论
在整个冷藏过程中,采用普通包装或真空包装的4 个生产类群的公、母牦牛肉肌浆蛋白的氧化程度均增加,具体表现为:随冷藏时间的延长,2 种包装的羰基含量均呈波浪式上升,总巯基含量下降;普通包装表面疏水性显著增加(P<0.05),真空包装表面疏水性在0~6 d增加,随后降低;紫外吸收光谱改变,冷藏第3天光谱改变最为明显;245 kDa附近及100~135 kDa蛋白条带消失或弱化。相同包装方式的公、母牦牛肉之间肌浆蛋白羰基、总巯基含量和表面疏水性差异不显著(P>0.05)。相对于普通含氧包装,真空包装可以有效抑制肌浆蛋白氧化,并且有效延缓蛋白的氧化速率。对于相同包装组,不同生产类群之间肌浆蛋白的氧化程度差异显著(P<0.05),表明不同生产类群牦牛的生产性能和肉质确实出现了差异;但肌浆蛋白氧化与牦牛的性别无关(P>0.05)。
综上所述,牦牛肉在低温贮藏及加工过程中,应从包装方式及牦牛的生产性能方面着手采取措施,以便有效抑制蛋白质的氧化,减少肉质在颜色、食用品质方面的劣变。
