辣椒Whirly基因家族的鉴定及表达分析
2019-04-29吕艳艳魏小春赵艳艳原玉香王志勇杨双娟郑小兰张梦园姚秋菊张晓伟
吕艳艳,魏小春,赵艳艳,原玉香,王志勇,杨双娟,郑小兰,姜 俊,李 艳,张梦园,姚秋菊,张 强,张晓伟
(1.河南省农业科学院 园艺研究所, 河南 郑州 450002; 2.郑州大学 生命科学学院, 河南 郑州 450001;3.驻马店市农业科学院,河南 驻马店 463000)
Whirly基因家族的蛋白质成员在整个植物界中都存在,主要定位于线粒体和叶绿体中,具有和单链DNA结合的能力[1]。Desveaux等[2]在2000年从马铃薯中分离出的PBF-2转录因子是第一个Whirly基因家族成员,它和cDNA编码的蛋白质都以序列特异性的方式与诱导子响应元件 (Elicitor response element)单链形式结合。
Desveaux等[3]在2005年研究发现,拟南芥和马铃薯中的Whirly转录因子具有调节防御基因表达的作用,其可能在防御反应以外的过程中发挥作用,并可能在叶绿体和核中起作用。Whirly蛋白在细胞核内参与水杨酸(SA)依赖的抗病信号转导,Desveaux等[4]研究发现,拟南芥中StWhy1同源基因AtWHY1的DNA结合活性由SA诱导,并且是SA依赖的抗病性和SA诱导的系统获得性抗性(SAR)响应基因表达所需的。Whirly蛋白家族能够调节端粒结构稳定,拟南芥AtWHY1被鉴定为一个新的端粒末端结合蛋白,在调节拟南芥端粒的稳态中能够发挥作用,主要表现在:在AtWHY1缺陷型植株中端粒延长,端粒酶活性增加;在含有AtWHY1的转基因植物中端粒酶活性降低,端粒缩短[5]。Whirly蛋白在质体内能够维持质体基因组的稳定,调节质体基因的表达。Marechal等[6]发现,Whirly单链DNA结合蛋白在维持拟南芥质体基因组稳定方面起到重要的作用。Cappadocia等[7]的研究结果表明,Whirly蛋白与单链DNA的结合有助于准确修复断裂的双链DNA。
辣椒(CapsicumannuumL.)是茄科辣椒属的1年生或多年生草本植物,是世界上种植面积最广的香料作物,目前辣椒的全基因组测序和物理图谱已经完成[8]。目前国内外对Whirly基因家族的研究主要以拟南芥为对象,针对Whirly蛋白的结构和功能进行研究,而对于辣椒Whirly基因家族的研究还没有详细报道。为此,利用生物信息学的方法,对辣椒CM334基因组中Whirly基因家族的全部序列进行鉴定分析,并利用荧光定量PCR(qRT-PCR)检测了CM334中Whirly基因在逆境胁迫下的表达情况,为进一步研究Whirly基因家族在辣椒生长发育中的功能奠定基础。
1 材料和方法
1.1 植物材料
所用辣椒材料CM334由韩国首尔国立大学馈赠。辣椒培养条件参考文献[9-10]。当辣椒幼苗有6~8片真叶展开时按如下方法处理。
脱落酸(ABA)和茉莉酸甲酯(MeJA)处理:用脱落酸(100 μmol/L)和茉莉酸甲酯(200 μmol/L)分别喷洒辣椒叶片的正反面,以喷洒无菌水作为对照。
高温处理(Heat):对辣椒叶片进行高温处理,白天温度设置为(42±2)℃,晚上设置为(35±2)℃。对照组白天温度设置为28 ℃,晚上设置为26 ℃[11]。
低温处理(Cold):预先将光照培养箱的温度降至5 ℃(昼)/0 ℃(夜),随后将辣椒苗放入光照培养箱中培养。对照组培养温度与高温处理对照组相同。
疫病(YB)处理:将辣椒幼苗的根置于用Hoagland′s培养液配制的病原菌(来自河南农业大学毛庄实验基地)孢子悬浮液中,以无菌水作对照。28 ℃保湿24 h后,在温度28 ℃、相对湿度70%~90%条件下培养。
上述处理的辣椒植株在培养箱中培养的光照强度均为160 μmol/(m2·s),光周期均为12 h光/12 h暗。
1.2 辣椒Whirly基因家族成员的鉴定
将Whirly(pfam:PF08536)(http://pfam.xfam.org/)[12]的保守结构域在辣椒基因组数据库PGP(http://peppergenome.snu.ac.kr/,CM334蛋白)中进行Blast比对,获得的所有候选序列在NCBI(National Center for Biotechnology Information)中进行Blast 比对。利用在线分析工具ProtParam (http://web.sxpasy.org/protparam/)分析候选氨基酸序列的等电点及分子质量[13]。
1.3 基因结构和蛋白质基序分析
以候选Whirly家族成员的CDS序列和基因序列为参照,通过在线GSDS 2.0(Gene Structure Display Server,http://gsds.cbi.pku.edu.cn/)进行基因结构分析[14]。分析时将CDS序列和基因序列都转换成FASTA格式,还要保持同一基因2种序列的对应。用MEME Suite 4.12.0在线软件(http://meme-suite.org/tools/meme)对Whirly家族蛋白质基序进行鉴定。
1.4 系统进化树分析
Whirly家族蛋白质全长序列用ClustalW进行多序列比对,ClustalW比对参数如下。空位罚分:10;空位延伸罚分:0.2;蛋白质重量矩阵:gonnet;残留特定的罚分:开;亲水罚分:开;间隙分隔距离:4;末端间隙分割:关闭;负矩阵:关闭;延伸分歧截止:30%。比对后Whirly蛋白序列用MEGA 7.0软件,使用邻近法构建进化树,其中校验参数Bootstrap值设置为1 000次重复。
1.5 辣椒Whirly基因组织表达模式分析
在辣椒参考基因组(https://www.nature.com/articles/ng.2877)中获取CM334 RNA-seq数据,利用HemI绘制Whirly基因在辣椒不同组织和果实发育中的表达热图。
1.6 辣椒总RNA的提取及cDNA第1链的合成
总RNA提取采用TaKaRa MiniBeST Plant RNA Extraction Kit (TaKaRa, Inc., Dalian, China)试剂盒,按照说明书进行操作。cDNA第1链合成采用PrimeScriptTM1st Strand cDNA Synthesis Kit(TaKaRa, Inc., Dalian, China)。
1.7 辣椒Whirly基因家族表达分析
采用qRT-PCR分析,引物见表1。反应体系为20.0 μL,其中cDNA为2.0 μL,上、下游引物均为1.0 μL,2×SYBR®Premix Ex TaqTM(TliRNaseH Plus) 10.0 μL以及ddH2O 6.0 μL。设置程序:95 ℃预变性30 s;95 ℃变性5 s,55 ℃退火30 s,72 ℃延伸60 s,循环40次[9]。采用2-ΔΔCT法计算基因相对表达量[15]。
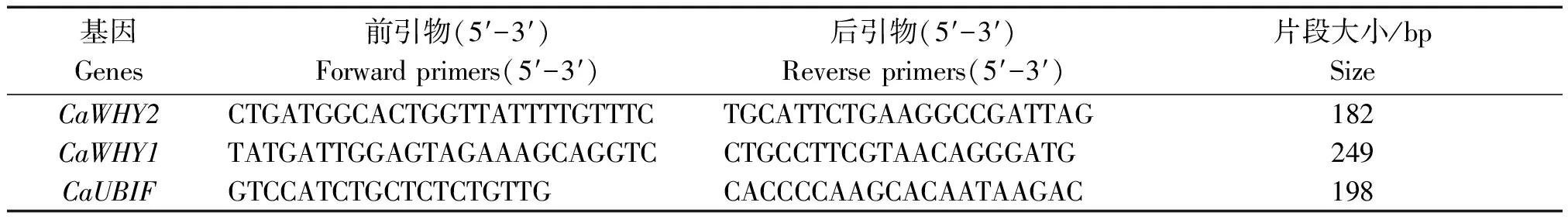
表1 qRT-PCR引物序列Tab.1 qRT-PCR primer sequences
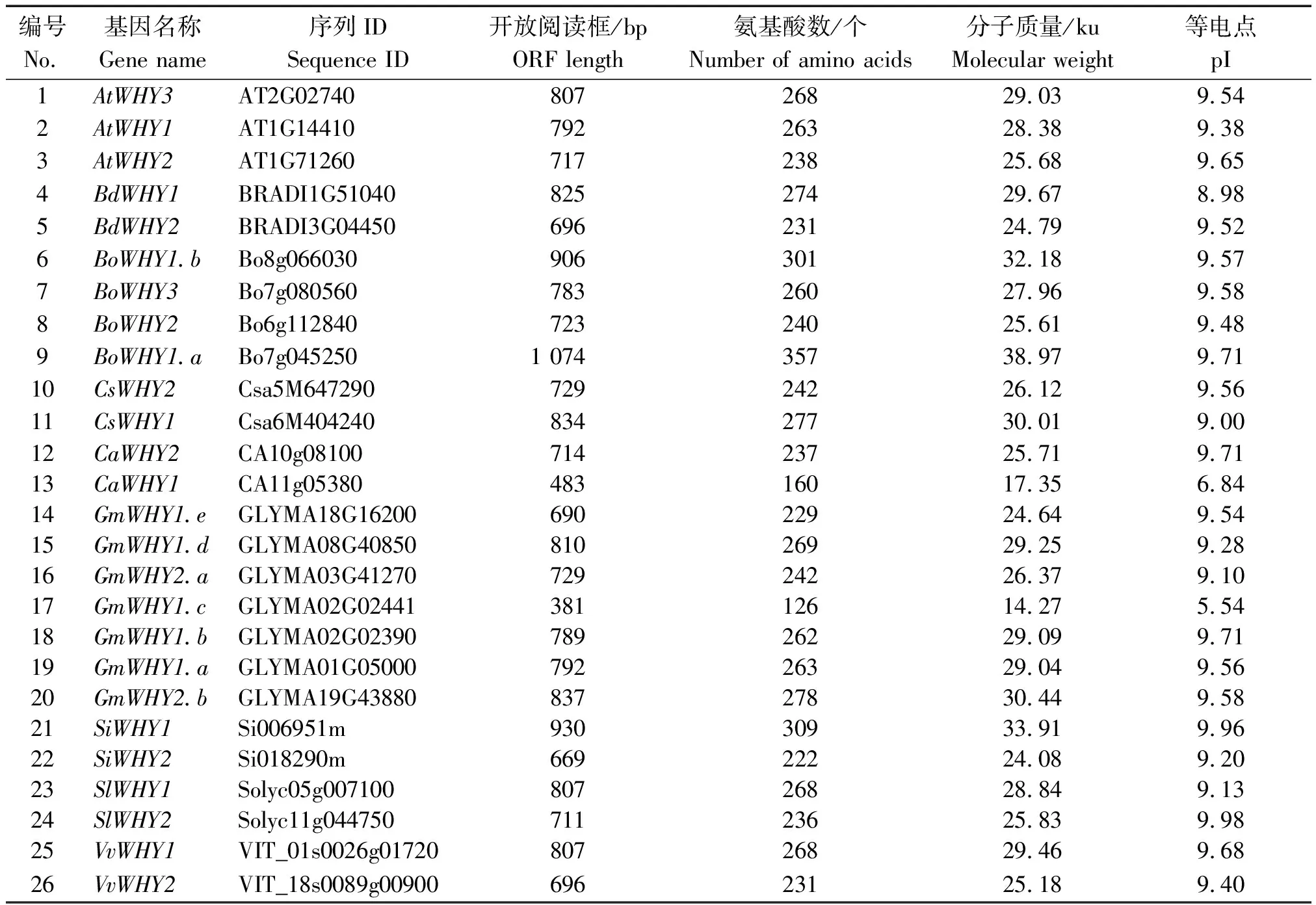
表2 Whirly基因家族成员鉴定结果Tab.2 Whirly family member identification list
2 结果与分析
2.1 辣椒Whirly基因家族成员的鉴定
以拟南芥的Whirly蛋白氨基酸序列为查询序列进行BlastP相似性检索,共鉴定了26个单子叶(短柄草、谷子)和双子叶(辣椒、拟南芥、甘蓝、黄瓜、大豆、番茄、葡萄)的Whirly基因家族成员(表2),其中辣椒CM334的Whirly基因有2个,分别为CaWHY1和CaWHY2。从表2可知,除了BdWHY1(pI=8.98)、CaWHY1(pI=6.84)、GmWHY1.c(pI=5.54)外,其他Whirly蛋白的等电点均在9.50左右。除CaWHY1和GmWHY1.c的分子质量分别为17.35,14.27 ku之外,其余Whirly蛋白的分子质量均在24.08~38.97 ku。
2.2 辣椒Whirly基因结构分析
通过GSDS基因结构分析可知,辣椒Whirly基因家族中CaWHY1有5个外显子、4个内含子,CaWHY2有7个外显子、6个内含子,CaWHY2和SlWHY2的基因结构完全一样(图1)。从图1还可以看出,除GmWHY1.c有2个内含子外,其他Whirly基因都有多个内含子(4~8个)。
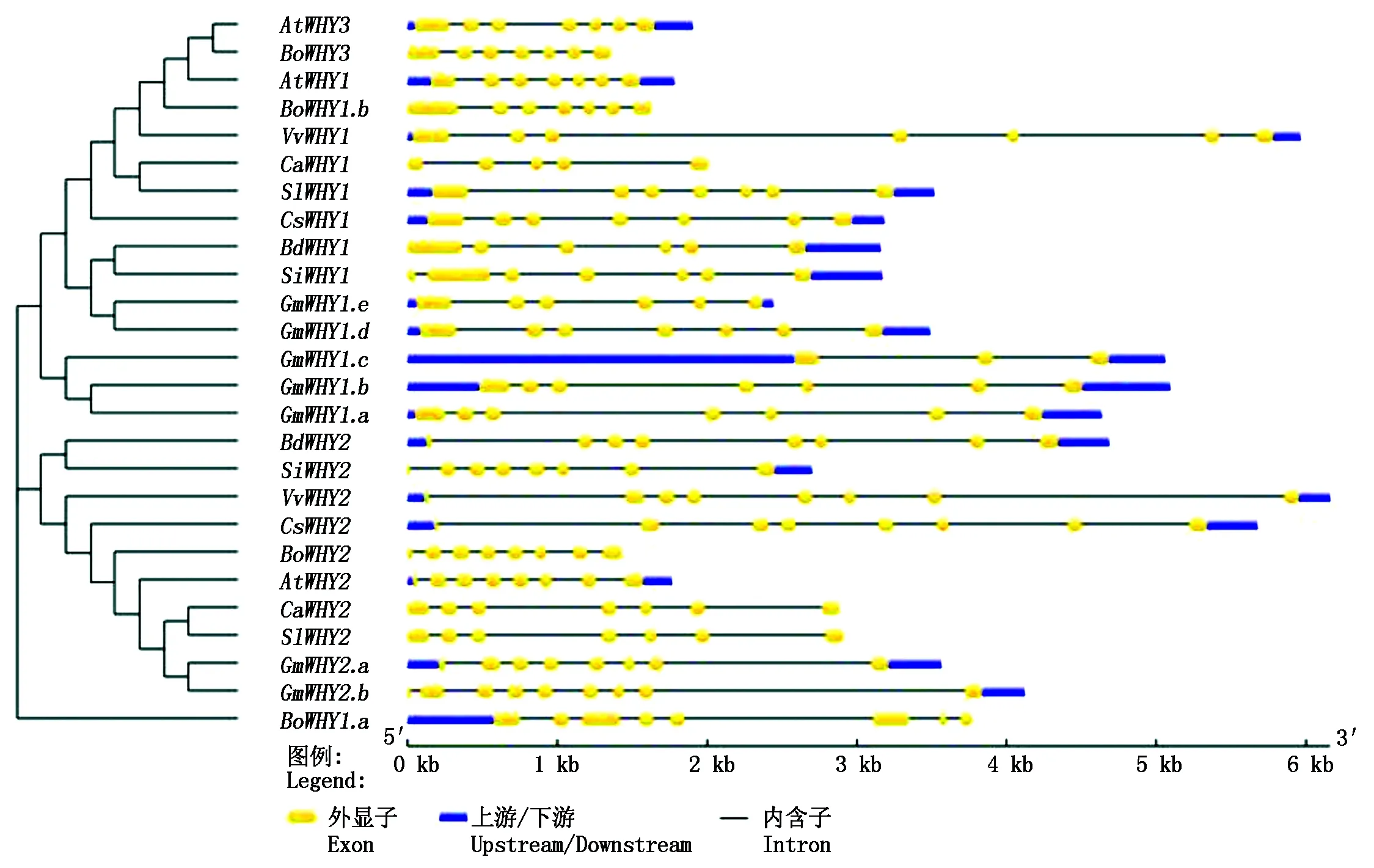
图1 26个Whirly家族基因结构分析Fig.1 Structure analysis of 26 Whirly family genes
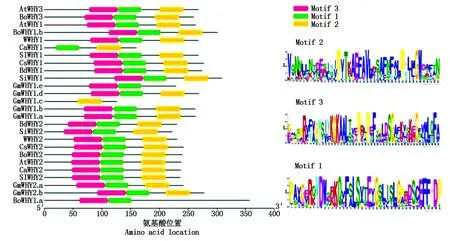
图2 26个Whirly蛋白保守基序分布Fig.2 Motif distribution of 26 Whirly proteins
2.3 辣椒Whirly蛋白保守基序分析
为了研究Whirly转录因子之间结构的多样性,利用MEME Suite 4.12.0 在线工具对26个不同物种的Whirly蛋白进行保守基序分析[16]。如图2所示,在Whirly家族中鉴定出3个保守基序。之前有研究报道表明,Whirly蛋白有3个结构域:N末端结构域、Whirly结构域和C末端多变区[17]。通过基序分析发现,Whirly基因家族在进化上结构具有保守性。
2.4 辣椒Whirly蛋白系统进化树分析
为更好地分析辣椒Whirly基因家族的进化,构建了辣椒、拟南芥、短柄草、甘蓝、黄瓜、大豆、谷子、番茄和葡萄的Whirly蛋白系统进化树(图3)。根据进化树的聚类结果,26个不同物种的Whirly蛋白明显分为3类:Class Ⅰ、 Class Ⅱ、Class Ⅲ。几乎所有的Whirly1属于Class Ⅰ,Whirly2属于Class Ⅱ,Whirly3属于Class Ⅲ。CaWHY1和SlWHY1聚在一起,CaWHY2和SlWHY2聚在一起,辣椒和番茄都属于茄科植物,其基因在功能上也有相似之处。同时,可以明显看出单子叶植物和双子叶植物分别聚在一起,表明单子叶和双子叶植物的Whirly基因在进化上有差异。
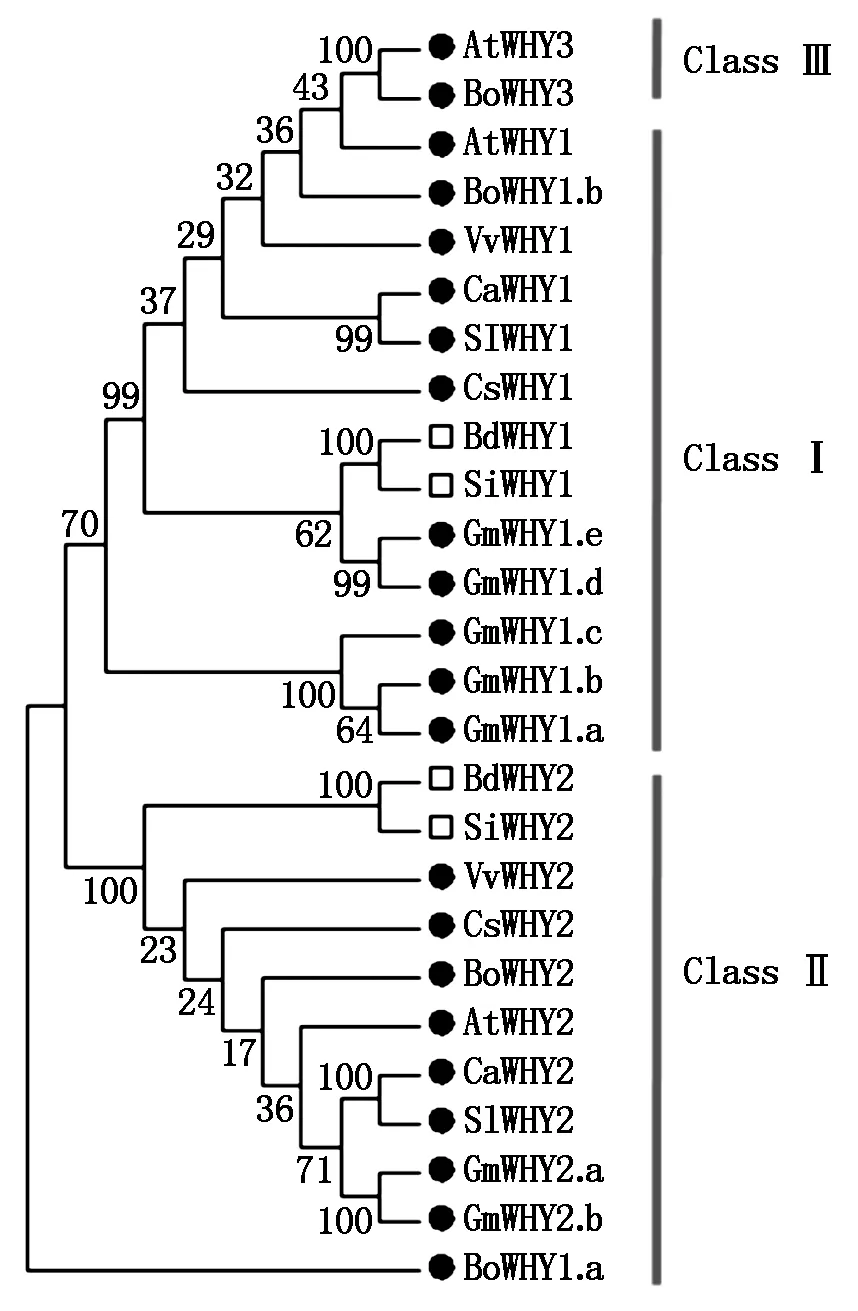
●.双子叶植物;□.单子叶植物。●.Dicotyledon;□.Monocotyledon.
2.5 辣椒Whirly基因时空表达分析
为了探究Whirly基因在辣椒不同组织及果实发育中的作用,利用CM334的RNA-seq数据绘制了Whirly基因家族在4个组织(根、茎、叶和果实)以及7个果实发育时期的热图(图4)。结果显示,CaWHY1和CaWHY2在不同组织中的表达量有差异,CaWHY2在根、茎和叶中均表达,CaWHY1在根、茎、叶中表达量却很低。同时,在果实发育中CaWHY1表达量也很低。

图4 Whirly基因在辣椒不同组织和果实发育中的表达模式Fig.4 Expression analysis of pepper Whirly genes in different tissues and fruit development
2.6 辣椒Whirly基因家族胁迫响应表达分析
利用qRT-PCR分析了辣椒CM334中的Whirly基因家族在逆境胁迫下的表达情况(图5)。辣椒的CaWHY1基因表达量在疫病胁迫下变化最为明显, 0~2 h急剧上升,2 h时达到最大值,为对照的4.5倍,随后表达量逐渐下降,24 h时下降到最小值,24~48 h又逐渐上升,48 h时其表达量为对照的1.8倍。CaWHY1在高温处理条件下,24 h时表达量达到最大值。在脱落酸、低温处理、茉莉酸甲酯胁迫下CaWHY1的表达量变化幅度相对不大。
CaWHY2在疫病和高温处理条件下的表达量变化最为明显,其中疫病胁迫下,在2 h 时其表达量达到最高值,为对照的2.3倍,随后逐渐降低,在12 h之后缓慢降低有趋于稳定之势,但最后表达量较对照有所下降。CaWHY2在高温处理条件下,2 h时其表达量上升至对照的1.7倍,6 h时又显著下降,之后逐渐上升,在24 h时表达量达到最大值,为对照的2.1倍,之后又逐渐下降,48 h时下降至对照的1.6倍。CaWHY2在脱落酸和低温处理条件下的表达模式相反,即当脱落酸胁迫下的表达量上升时,低温处理条件下的表达量下降,但脱落酸和低温处理条件下表达量的变化幅度不同。CaWHY2在茉莉酸甲酯处理条件下,0~2 h有不明显的上升,之后逐渐下降,24 h时降至最低值,约为对照的1/2,24~48 h逐渐上升,48 h时达到最大表达量,为对照的1.7倍。
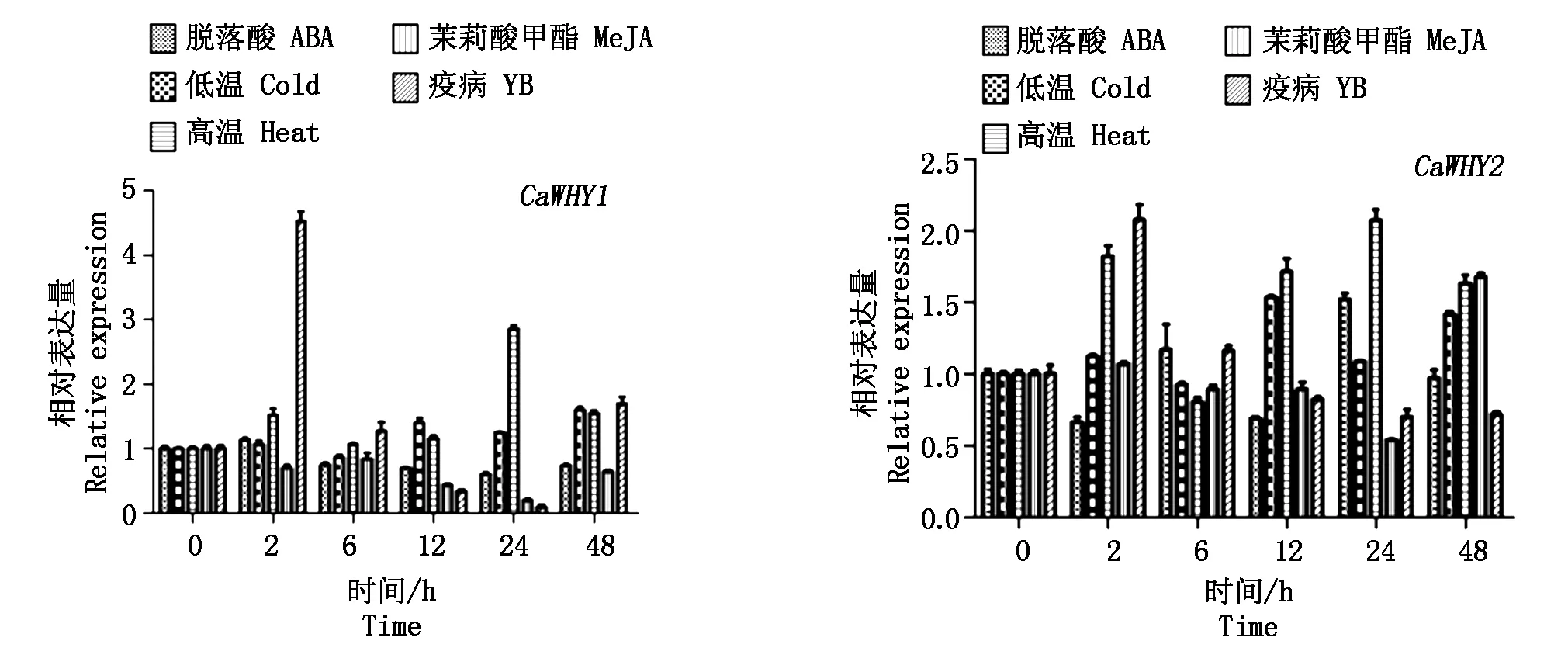
图5 辣椒Whirly基因家族响应胁迫的qRT-PCR分析Fig.5 qRT-PCR analysis of pepper Whirly gene family reacting to stress
3 结论与讨论
Whirly基因家族在双子叶和单子叶植物中均有发现。本研究在辣椒CM334中鉴定了CaWHY1和CaWHY22个Whirly基因,其中CaWHY2和番茄SlWHY2的基因结构完全一样,均有7个外显子、6个内含子,在进化树中CaWHY2和SlWHY2也是聚在一起,这可能是由于辣椒和番茄均属于茄科,亲缘关系较密切[18]。对蛋白质基序分析发现,Whirly蛋白在植物中高度保守,含有3个保守的基序。Whirly基因表达模式分析表明,CaWHY1和CaWHY2在不同组织和果实发育时期的表达水平存在差异。在脱落酸、低温处理、高温处理、茉莉酸甲酯和疫病处理条件下,CM334的Whirly基因均有表达。在疫病处理下,CaWHY2和CaWHY1表达量均增加,其中CaWHY1基因的表达量较CaWHY2增加更明显。CaWHY2基因在脱落酸和低温处理条件下表达量的变化趋势相反。
Whirly转录因子在植物的生长发育过程中起到很重要的作用[19]。另有研究证明,Whirly转录因子在叶绿体和细胞核中也发挥一定作用[20]。本研究对辣椒Whirly基因表达模式分析表明,Whirly基因家族在辣椒的生长发育中起调控作用。植物需要适应不利的环境条件,包括生物和非生物胁迫[21-22]。杨洋等[23]研究发现,Whirly转录因子在植物应对低温胁迫中发挥作用。本研究中,Whirly基因在各种胁迫处理下的表达量变化再次证明,Whirly基因在多种生物和非生物抗性反应中起重要作用。
Whirly基因在植物界中广泛存在,在其他物种尤其是拟南芥中Whirly基因的研究较为详细,但是有关辣椒Whirly基因的报道却较少。本研究鉴定了辣椒CM334中的Whirly基因,对其结构、进化关系、保守基序和表达模式进行了初步分析,并对其在逆境胁迫下的表达量变化进行了分析,为进一步研究Whirly基因在辣椒中的功能奠定了基础。
