青海春小麦品种高原363成株期抗条锈病基因遗传模型分析
2019-04-29侯璐
侯 璐
(青海大学 农林科学院,青海省农林科学院,青海省农业有害生物综合治理重点实验室,农业部西宁作物有害生物科学观测实验站,青海大学省部共建三江源生态与高原农牧业国家重点实验室,青海 西宁 810016)
小麦条锈病是世界范围内的病害,该病害是由于小麦条锈菌(Pucciniastriiformisf. sp.Tritici)引发的。通常该病最容易发生在低温并且潮湿的环境内,一般来讲,只要是植株仍旧是绿色,也就是苗期到成熟期之内,都是有可能得病的[1]。经过实践与相关研究证明,最具有性价比并且环保的方式来防治此病害就是利用和种植具有抗病的小麦品种[2-3]。但是自然界中小麦条锈菌群体的毒性结构复杂,不断出现的新的小麦条锈菌毒性小种,容易使正在推广的生产中的小麦品种由原来的抗病转变为感病,因此,导致小麦条锈病一直多发而从未消减过,对此只能不断地寻找新的抗条锈病小麦材料来挖掘新的抗病基因,从而达到多样化的抗条锈病基因并能合理的布局,进一步对小麦抗条锈病基因遗传特点的研究也是一项防治小麦出现此类病害的重要基础研究。
我国有些地区在夏季时会流传此病害,青海省正是其中之一[4]。在青海省,春小麦和冬小麦交错种植,且小麦的种植呈垂直分布,每年以晚熟春麦上的条锈病菌源为主的大量越夏菌源,可以一直保持到早期播种的冬小麦出苗以后,从而为我国东部冬小麦生产区提供条锈病菌。青海省近几年种植的春小麦品种主要是“阿勃”,这个品种是高感此病害的品种,青海省在生产上表现抗病的品种较少,这种情况为我国小麦条锈病越夏区菌源的治理带来很不利的影响。
为了增加抗条锈病基因的多样性,我国科研工作者做了许多工作。在对我国农家品种和生产品种的抗条锈病性研究中,徐世昌等所在研究组做了大量系统的抗病性鉴定和研究工作[5-10]。在对不同省份的小麦品种的抗条锈性研究上:曹世勤等[11]对甘肃省的 26 个春麦品种(系)做了结合苗期和成株期测试的抗病性评价,并且在2003-2013年这10 a持续地完成了对4 291份冬小麦和710份春小麦的抗条锈性鉴定[12]。李永平等[13]对临麦系列春小麦品种进行结合苗期测试和成株期测试的抗病性评价分析;王万军等[14]对86份贵协系小麦种质资源进行了成株期抗条锈病鉴定,发现9份资源表现了相对稳定的成株期抗病性。在对春小麦种质资源的抗条锈病性研究方面,周新力等[15]对80份引自美国的春小麦种质资源进行了系统的苗期和成株期抗条锈性评价;姚强[16]评价了青海、西藏等地区的120份春小麦品种(系)的成株期抗条锈性,侯璐等[17-18]先后对搜集的共483份春麦资源进行了结合苗期测试和成株期抗病性测试的系统抗条锈性鉴定。在对抗条锈性春小麦种质资源中条锈病基因遗传分析研究方面:侯璐等[19-20]先后分析了8份抗病春小麦种质资源的苗期抗条锈病基因的遗传特点,姚强等[21]分析了青春39中苗期抗条锈病基因的遗传规律;而目前国内针对春小麦品种成株期的抗小麦条锈病基因的遗传特点研究仍旧较少,仅张调喜等[22-23]做了春小麦品种墨波(Mobo)和青春38的成株期抗条锈病基因的遗传解析。
青海农科院植保所植物病理研究组从2010-2016年的大田条锈病成株期感病情况调查,发现春小麦品种高原363一直表现高抗条锈病,是很好的抗源材料[16],本研究主要探究高原363的成株期抗条锈病基因的抗性遗传特点和规律,以对其的有效合理利用提供理论参考。
1 材料和方法
1.1 试验材料
高原363; Taichung 29 (T29)(中国农业科学院植物保护研究所的徐世昌先生提供最初种子);高原363作为父本,和T29杂交自交后获得F2∶3群体。
1.2 试验方法
2015年和2016年分别于青海省东海市互助县和西宁市的小麦条锈病抗性鉴定自然发病圃进行了抗病性鉴定。
2个亲本均是种1行,1 m的行长,每行间隔0.3 m;在2015年使用单粒点播的方式种植F2,1行总共播种10粒; 2015年种植的无论是互助县还是西宁市的F2单株群体,均收获到了相应的F3家系群体的种子。2016年将F3家系群体于相同的地方继续种植;在每块试验地中每播种20行就插种1行T29作为感病诱发对照组,并且在鉴定圃四周亦种植3行T29作为发病诱发行,按照节气,于每年的4月上旬进行播种,T29一般在7月上旬充分发病,因此,在这个时间段内调查亲本和群体,2015年调查F2群体中的每个单株, 2016年对每个F3家系进行调查。将反应型和严重程度均详细记录反馈。不同次数的调查之间间隔为7 d。一共进行2次调查,使用Hou等[24]0~9级的记录方法来记录反映型,田间病害严重度(DS,%)按0~100的调查方法分级记录[15],选用2次调查中发病最严重的数据作为最终统计。
2个F2∶3群体在各个环境中的严重度和反应型的频率分布条形图由软件 Excel 2016 统计并绘制;并且使用植物数量性状主基因+多基因混合遗传模型分析软件[25]对数据进行解析和分析。使用期望最大化以及极大似然算法估算分布参数。适宜遗传模型的选择标准,一般为AIC值较小以及适应性检验结果比较优的模型。使用最小二乘法来估计一阶遗传参数和二阶遗传参数,估计得出的二阶遗传参数包括了主基因和多基因的遗传方差以及遗传率等数据。
2 结果与分析
2.1 条锈病抗性统计分析
2015年与2016年在青海两地的试验中,感病亲本T29的病害反应型均为9级,严重度为95%,说明试验地条锈病发病比较充分,而高原363的反应型为3级,严重度为10%;F2∶3群体在2个环境中反应型和严重度大小的频率分布如图1,2。由图中发现,F2∶3群体在2个环境下反应型和严重度均呈现出了双峰分布的现象,而且整体上的连续性分布现象并未出现,但是不同的区段内却又出现了连续性的现象。对此分析可能是因为由主效基因和微效多基因共同控制的复杂遗传表现的现象。

图1 高原363/T29 F2∶3群体在互助试验地所测试病害反应型和严重度的频率分布Fig.1 Analysis of frequency distributions of Gaoyuan 363/T29 F2∶3 populations the infection type and disease severity tested in Huzhu

图2 高原363/T29 F2∶3群体在西宁试验地所测试反应型和病害严重度的频率分布Fig.2 Analysis of frequency distributions of Gaoyuan 363/T29 F2∶3 populations the infection type and disease severity tested in Xining
2.2 遗传模型的选择
分析结果如表 1,得到11 种遗传模型的AIC值,包括0 对主基因(A)、1 对主基因(B)和2 对主基因(C) 3类。选取 AIC 值最小的3个或者与最小AIC值比较接近的遗传模型为候选模型,获得候选模型的U12、U22和 U32、nW2和Dn统计量,选择模型时考虑显著的检验个数问题,将显著检验个数最少的模型选择为最优遗传模型。
2.2.1 高原363的成株期抗条锈性基因遗传模型的选择 根据互助测试得到的分析结果如表1,由严重度数据进一步分析得到, C-5、C-6和 C-2的AIC值最低,分别为1 582.23,1 584.72和1 586.23;适合性检验结果发现,该3个模型都是只有1个统计量达到显著水平,C-5 模型的 AIC 值最小,故作为最适合遗传模型。当用反应型数据分析时,C-1、C-5和C-6的AIC 值最低,分别为542.79,586.18和588.08;结合适合性检验结果分析,C-6模型达到显著的检验个数最少,为1个,因此,将其作为最适合的遗传模型。
在西宁测试的分析结果如表1,用严重度数据分析时,C-6、C-5和C-3的 AIC 最低,分别为639.34,639.94和641.48;在适合性检验结果中得知,3个模型均未表现出显著水平的统计量,C-6即为最适合的遗传模型,因为其AIC值最小。当用发病反应型数据分析时,模型C-1和C-6和C-5的 AIC 值最低,分别为198.56,250.07和252.07;适合性检验结果分析中得知,3个模型的统计量中均只有一个达到了显著水平,因为C-1模型的AIC值最小,即为最适合的遗传模型。
由以上分析结果可知,2个地点中选用严重度数据或反应型数据分析时,最适宜的遗传模型均是表现出了2对主基因的遗传方式,主基因的作用方式不同,其中C-1模型为加性-显性-上位性作用,C-5模型为完全显性作用,C-6模型为等显性作用。
2.2.2 最适遗传模型相关参数估计 反应型和严重度的一阶和二阶遗传参数由最优遗传模型估计(表2)。高原363/T29 F2群体在互助测试,对严重度数据进行分析时,高原363的成株期抗条锈病基因作用方式表现为C-5模型,也就是加性效应均表现出正向效应的2对完全显性主效基因表现的作用;当使用反应数据分析时,发现高原363的成株期抗条锈病基因作用方式如同C-6模型表现出的模式,也就是2对等显性主基因作用的结果,并且2对基因的加性效应只有一个呈现出正向效应。在西宁的测试中发现,以严重度数据进行分析得出的为C-6模型,也就是2对等显性主基因作用,并且这2对基因的加性效应只有一个呈现出正向效应;而以反应型数据为分析得出的结果呈现出C-1模型,也就是2对呈现出加性-显性-上位性的主基因作用的结果,除了其显性效应均是负向效应,基因间加性×加性互作为负向,其他互相作用效应均显示为正向效应。在互助县的试验地条件下,主基因的遗传率分别为94.10%,87.64%,而在西宁市的试验地条件下主基因的遗传率为93.93%,99.26%,由此证明2个不同环境下成株期高原363的抗条锈基因都表现很强的遗传能力,并且在2个环境下主基因的遗传率差距并不大。说明主基因具有较高的遗传稳定性,受环境因素影响小。

表1 高原363/T29 F2 群体在2个试验地病害反应型和发病严重度在各遗传模型下的AIC 值Tab.1 AIC values of genetic models by Gaoyuan 363/T29 F2 populations the infection type and disease severity tested in two environments
注:MG.主基因;A.加性效应;D.显性效应;I.互作;N.负向;E.相等;下划线表示 AIC 值较小,加粗数字表示属于最优遗传模型。
Note: MG.Major gene; A.Additive;D.Dominance;I.Interaction;N.Negative; E.Equal; Minimum AIC value in each cross is underlined, data in bold belonged the most fitted genetic model.
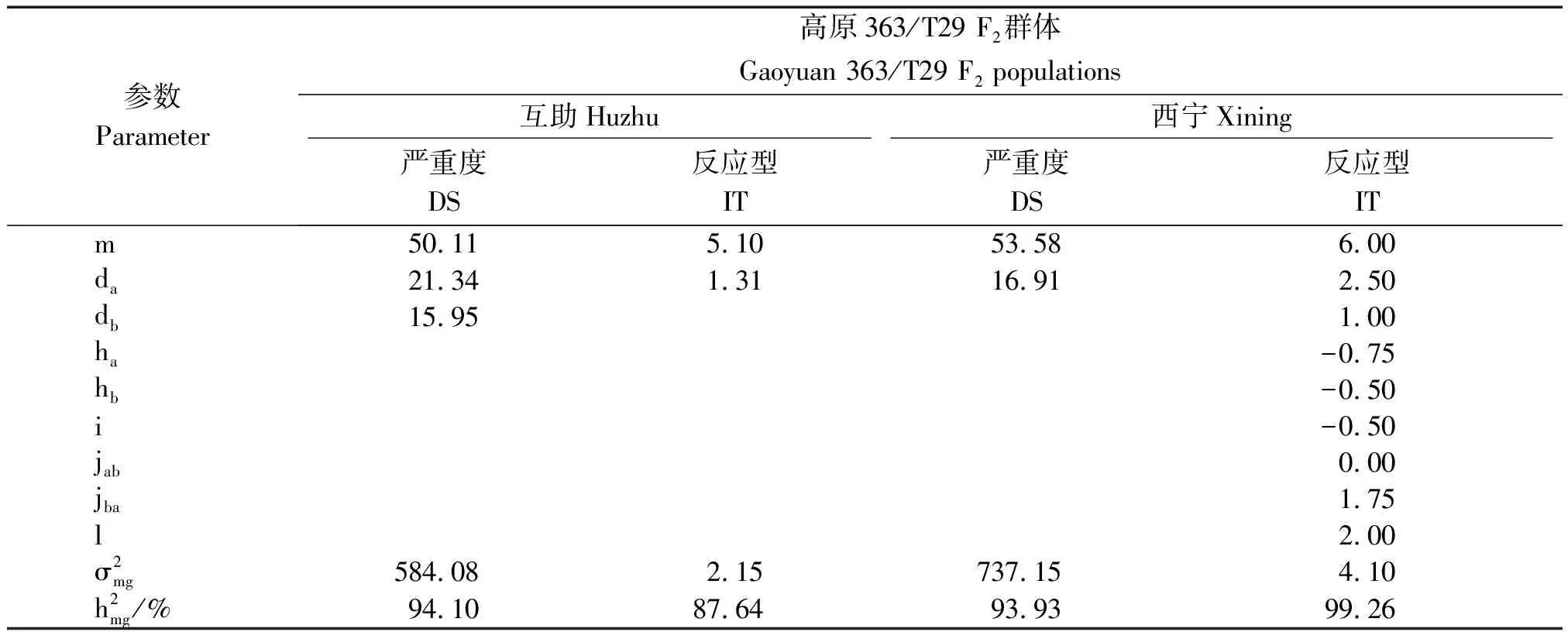
表2 高原363/T29 F2群体在最适遗传模型下估计的遗传参数值Tab.2 Genetic parameters of Gaoyuan 363/T29 F2 populations estimated from the best models


3 讨论
作为青海省的春小麦主栽品种,高原363由中国科学院西北高原生物研究所1975年以高原2D巨穗单体和B3-1-1特多实矮秆突变体为基础,应用人工染色体工程引入外源优异基因,利用F2单体优株复合杂交法综合、固定累加多种属多品种的优秀基因,并且与常规选育方法相结合而培育出,不仅仅具有耐旱、耐青干的特点以及抗倒伏的特点,更是具有抗条锈、叶锈以及杆锈的特点[26]。本研究组从2010-2016年7 a连续观察,发现成株期高原363对条锈病的抗性一直很高。研究其抗病遗传机制对科学利用这一抗病资源具有重要意义。
目前,国内发表的主要是依据孟德尔遗传规律结合病害反应型数据分析的方法对小麦成株期抗锈病基因的遗传分析,仅能分析出1~3对成株期抗锈病基因的作用规律[27-31]。本研究组报道了对春小麦品种墨波和青春38的成株期抗条锈基因的遗传解析[22-23],也是利用本研究中使用的混合遗传模型来进行分析,和本研究中的情况相仿,也就是杂交群体的抗病性分离结果并不符合纯数量性遗传的连续性分布,在鉴定结果中发现中间型较多,在人为抗病与感病划分时容易出现较大的误差,特别是在该研究小领域中,对小麦条锈病的病害严重度的结果还未进行过人为的抗病、感病的划分,而直接应用植物数量性状主基因+多基因混合遗传模型分析方法分析,既避免了人为划分抗、感病的误差,而且可以将病害严重度与发病反应型的数据结果结合到一起分析,使研究分析更丰富,结果更可靠。
本研究选取了高原363与T29 春小麦品种以及它们的杂交后代群体,在西宁市与互助县2个试验地点各进行了2年的田间成株期条锈病抗性鉴定研究,结果最终显示,高原363含有2对成株期抗条锈病主效基因,该2对主基因在不同环境或用不同调查参数(严重度或反应型)时表现的作用方式不一样。根据高原363中的成株期抗条锈病基因的遗传特点,可以使用分子标记的方法定位抗病基因,找到紧密连锁抗病基因的标记,在分子标记辅助选择育种的过程中可以起到作用,也可直接通过抗病基因聚合以进一步增强寄主的抗病性并提高抗性持久性。
