松果菊苷对大鼠Leydig细胞衰老模型AXL和wnt2b基因的影响
2019-04-26孙静庞云瑞单博颖薛亚然牛嗣云郑立双郗昕
孙静 庞云瑞 单博颖 薛亚然 牛嗣云 郑立双 郗昕
(1河北大学医学院组织学胚胎学教研室,河北 保定 071000;2保定市第一医院药学部;3河北大学附属医院中心实验室)
根据甲基化芯片筛查结果〔1〕,从中选出甲基化基因AXL和wnt2b,通过荧光定量聚合酶链式反应qRT-PCR、甲基化特异性PCR(MSP)检测技术,证明了肉苁蓉能够调控睾丸组织AXL和wnt2b基因的表达。本研究拟探讨松果菊苷对大鼠睾丸间质(Leydig)细胞衰老模型AXL和wnt2b基因的影响。
1 材料和方法
1.1睾丸间质(Leydig)细胞的原代培养和纯度鉴定 取用2周龄Wistar雄性大鼠幼崽,无菌取出双侧睾丸组织,浸泡于预冷的PBS中。剥离睾丸组织中白膜,分离得到新鲜、结构完整的睾丸组织。无菌胶原酶Ⅰ消化后,差异贴壁法分离、培养Leydig细胞。
1.2Leydig细胞衰老模型建立与鉴定 依据“自由基氧化损伤”理论〔2〕建立间质细胞衰老模型,将新鲜配制的过氧化氢(H2O2)和FeSO4加入已培养2 d的细胞中,使浓度分别为50 μmol/L和100 μmol/L,作用时间分别为8 h(即各加入的体积为2 μl),成功建立了细胞衰老模型,并用 β-半乳糖苷酶染色成功鉴定细胞衰老状态〔3〕。
1.3噻唑盐(MTT)比色法筛选松果菊苷作用最适浓度 在培养2 d的细胞中加入不同浓度的松果菊苷进行药物处理,浓度梯度设置为10、20、40、80、160和320 μmol/L,持续作用1、2、3 d,上机检测后计算细胞增殖活力,选择最佳作用于Leydig细胞的浓度。
1.4细胞分组 正常组(培养128 h),衰老组(氧化损伤8 h,继续培养72 h)和松果菊苷组(氧化损伤8 h,松果菊苷用药继续培养72 h)。
1.5qRT-PCR TRIzol法提取Leydig 细胞总RNA,以焦碳酸二乙酯(DEPC)水为对照孔,每孔加入2 μl,设置复孔,用酶标仪进行核酸定量RNA浓度,然后反转录成cDNA。三组Leydig细胞的内参均选择β-actin ,荧光染料(SYBR)进行实时定量检测。借助统计软件SPSS19.0,采用 2-△△ct计算方法统计分析三组的Ct值。引物序列:内参β-actin上游引物:CCCATCTATGAGGGTTACGC,下游引物:TTTAATGTCACGCACGATTTC;AXLF上游引物:GGTGGCTGTGAAGACGATG,下游引物:CTCAGATACTCCATGCCACT;wnt2b上游引物:GCTGGACCAAACCTGAACG,下游引物:CAAGAAGTATCGGGAAGCA。
1.6MSP 根据细胞基因组DNA快速提取试剂盒,提取各组Leydig细胞DNA,核酸定量DNA浓度,一步法完成DNA样本变性和亚硫酸盐转化,扩增甲基化基因AXL和wnt2b。3 % 琼脂糖凝胶电泳,借助ChampGel 5000系统自带Lane1软件,统计分析三组甲基化百分比。引物序列:wnt2b甲基化上游引物:GGGAAAGTAGTTTAGTTGTTTTTG,甲基化下游引物:AAACTCCTTTAACCACACTACCCTC;wnt2b未甲基化上游引物:CGAGGTGGCAAACATCCTATATTAAG,未甲基化下游引物:CTTTGAAGGCTCCACTCCTGCACACT;AXL甲基化上游引物:AAGTATAAGAGTTTTAACTTAAGGTGGG,甲基化下游引物:ACTTACGCTACACCGTACAAAAA;AXL未甲基化上游引物:GGTGGCTGTGAGTTAGTTGAGACGATG,未甲基化下游引物:CCAAACTCAGATACTCCATGTTCTTCC。
1.7统计分析 应用SPSS19.0软件行方差分析、LSD-t检验。
2 结 果
2.1MTT 筛选松果菊苷作用浓度结果 当松果菊苷浓度达到 80 μmol/L,无论作用24 h 或 48 h,细胞活力开始下降,320 μmol/L时下降最明显;松果菊苷作用 72 h 后,不同浓度间细胞活力无差异,其中相同作用浓度,随着作用时间延长,72 h时细胞活力最好(P<0.05)。综合而言,72 h时细胞作用浓度为 20 μmol/L时,细胞增殖活力最佳。见表1。
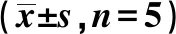
表1 Leydig 细胞的增殖活力
与320 μmol/L比较,1)P<0.05,与160 μmol/L比较:2)P<0.05;与80 μmol/L比较:3)P<0.05
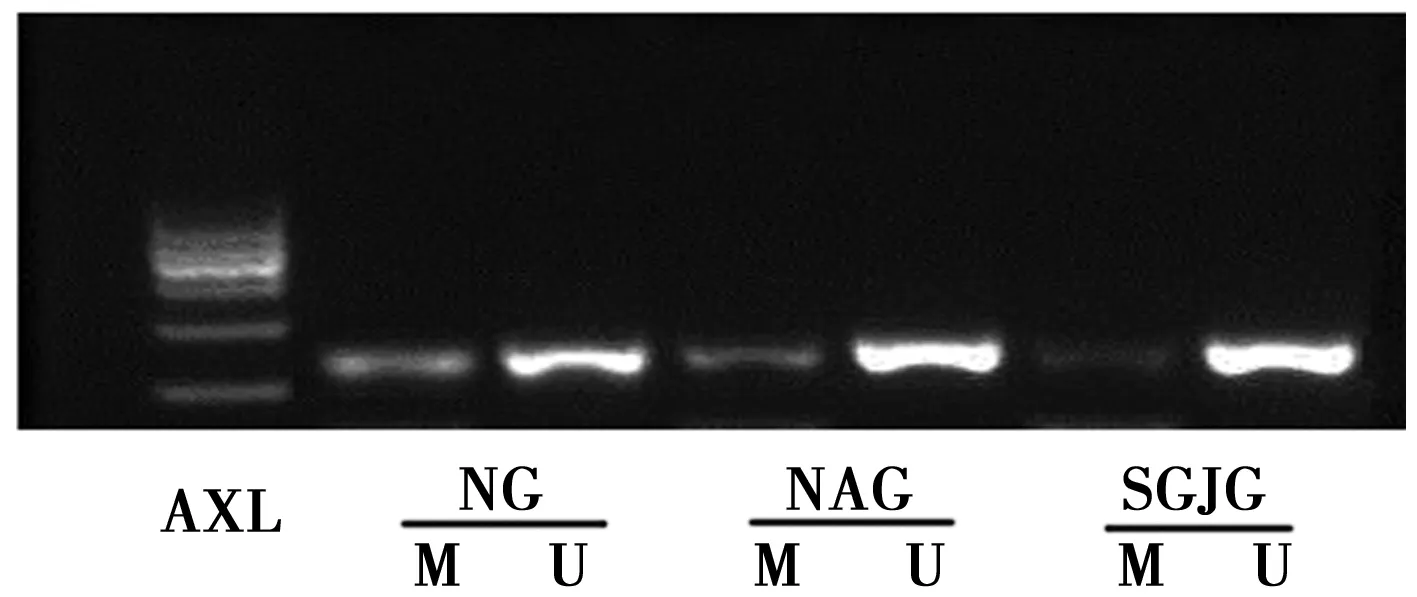
2.2松果菊苷对衰老Leydig细胞AXL基因的影响 衰老组Leydig细胞 AXL mRNA 的表达水平显著高于正常组,但甲基化水平明显低于正常组(P<0.05)。松果菊苷用药后AXL mRNA 表达水平明显低于衰老组(P<0.05),但AXL 的甲基化水平与衰老组差异无统计学意义(P>0.05)。见表2,图1。
2.3松果菊苷对衰老Leydig细胞wnt2b基因的影响 衰老组Leydig细胞 wnt2b mRNA 表达水平明显高于正常组(P<0.05),甲基化水平显著低于正常组(P<0.05)。松果菊苷组用药后wnt2b甲基化水平明显升高(P<0.05),但与衰老组Leydig细胞相比,二者wnt2b mRNA 表达水平无统计学意义(P>0.05)。见表3,图2。
表2 三组 AXL mRNA 的表达量
与衰老组比较:1)P<0.05,下表同
NG:正常组 AG:衰老组 SGJG:松果菊苷组 nc:阴性对照 M:甲基化 U:未甲基化,图2同图1 不同组AXL基因DNA甲基化水平变化
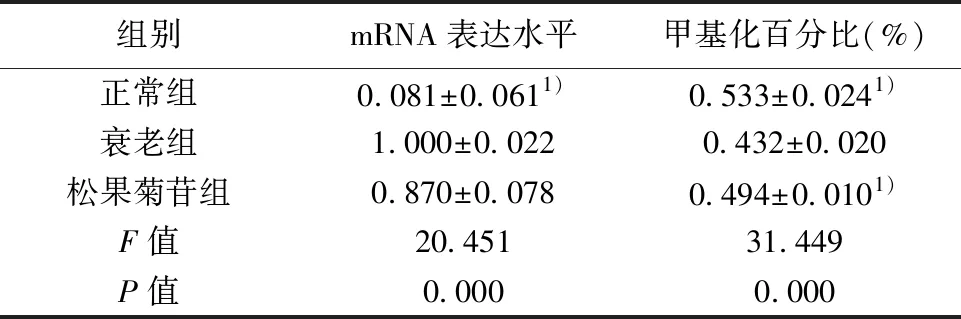
组别mRNA表达水平甲基化百分比(%)正常组0.081±0.0611)0.533±0.0241)衰老组1.000±0.0220.432±0.020松果菊苷组0.870±0.0780.494±0.0101)F值20.45131.449P值0.0000.000
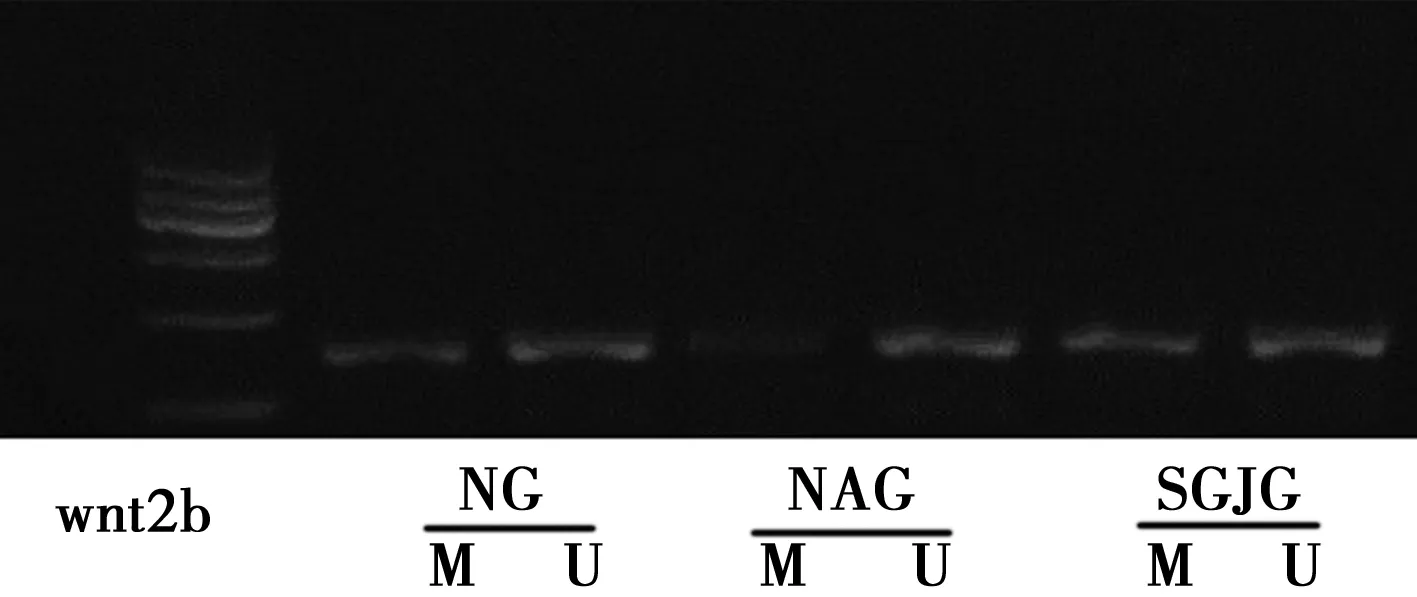
图2 不同组wnt2b基因DNA甲基化水平变化
3 讨 论
H2O2是一种性质稳定易于获得的强活性氧,常用于短期内氧化损伤诱导的细胞衰老〔2,3〕。de Silva等〔4〕用HaCaT 细胞系研究发现,HaCaT锚定阻滞会引起氧化应激反应,导致DNA损伤,从而引起了全基因组低甲基化趋势,表明氧化应激改变DNA甲基化从而导致基因组不稳定。Wang等〔5〕研究发现,SD大鼠通过饮用水口服Cr(vi)不仅会引起血浆氧化应激,而且会导致雄性大鼠血细胞的总体DNA低甲基化,并且在丙二醛(MDA)水平升高和总体DNA甲基化降低之间存在良好的相关性。目前,细胞衰老后的一些生物学特征,如形态结构的改变,衰老有关的β-半乳糖苷酶表达,细胞周期停滞在G1期等已经得到较好的阐述〔6〕。然而氧化损伤诱导细胞进入永久停滞状态的内在甲基化机制尚不清楚。因此,更好地理解老化过程中氧化应激和表观遗传相互作用,是确定老化过程关键基因改变的重要研究机制之一,对确定有效治疗衰老的药物,在衰老期间改善人类健康及衰老和老年病的研究意义重大。
表观遗传学逐渐吸引了越来越多的兴趣,因为它已经证明了在不修饰基因序列的情况下调节基因表达的能力,可以积极地影响炎症和癌症疾病以及神经退行性疾病的预防和发展过程〔7〕。老化时总的基因组DNA随机发生低甲基化。转座因子的DNA序列正常状态时沉默,DNA甲基化丧失时则可被激活。在不同的年龄段,一些甲基化的改变可以定向发生于基因组的特定区域〔8〕。老化过程中DNA甲基化的部分改变并非是随机发生的,大多与重复的DNA序列的CpG低甲基化有关〔8,10〕。Leydig细胞可能随着细胞的衰老,逐渐丧失睾酮合成能力〔11〕。进而推测Leydig 细胞的衰老状态,可能对男性的生殖功能有影响。
AXL 是受体酪氨酸激酶家族 TAM 受体中(Tyro3,AXL 和 MER)成员之一〔3〕。睾酮替代疗法(TRT)部分地通过增强Gas6表达来减轻细胞凋亡。此外,AXL的缺失使睾酮失效,这表明AXL是睾酮的重要下游调节剂。TRT将通过Gas6/ AXL信号通路改善与衰老相关的组织重塑,暗示其治疗老化相关疾病的治疗潜力〔12〕。本研究说明松果菊苷可以通过抑制衰老的 Leydig 细胞中 AXL mRNA 表达水平,缓解 Leydig 细胞的衰老状态。
wnt蛋白是一个分泌蛋白家族,调节基因表达、细胞增殖和细胞分化的许多方面〔13〕。wnt/β-catenin信号通路控制着发育和成人生活中的各种生物学现象〔14〕。睾丸中wnt/β-连环蛋白信号特异性促进SSC和祖细胞的增殖〔15〕。也有研究表示wnt2b在附睾头部高度表达,但在尾部几乎不存在〔16〕。wnt信号传导对于小鼠胚胎干细胞等多能性基因的转录调节至关重要。尽管如此,wnt信号传导与表观遗传调控机制之间的联系至今尚未阐明。有研究表明wnt信号作为参与表观遗传变化的关键参与者的新作用,该信号传导途径可以保护沉默基因组区域,并能维持基因组的稳定性〔17〕。研究发现,wnt信号传导参与睾丸减数分裂前未分化精原细胞的扩增〔18〕。wnt2b的过表达即可导致组织畸形生长,这主要是通过抑制细胞的周期和分化实现的〔19〕。本研究发现了松果菊苷可以通过提高wnt2b甲基化水平,改变Leydig细胞的衰老状态。
