辣根过氧化物酶在纤维材料生物整理中的应用研究进展
2019-04-25周步光范雪荣袁久刚
周步光, 王 平, 王 强, 范雪荣, 袁久刚
(生态纺织教育部重点实验室(江南大学), 江苏 无锡 214122)
由于淀粉分子结构本身的特点,使得淀粉浆料对疏水性纤维的黏附力不足[1]、成膜性差,为改善淀粉浆料对经纱的上浆性能,需要对淀粉进行接枝改性。黄麻纤维存在刚度大、刺痒、易成褶、染色深度不高、染鲜艳色难等缺点,并且植物纤维与非极性、疏水性树脂间的界面黏结性差,使其应用受到很大限制[2],因此,需要对黄麻纤维进行亲疏水改性处理。蚕丝织物经脱胶或溶解提取的丝蛋白中含有一定量的酪氨酸[3],其中的酚羟基有较高的反应性,可发生自交联或与外源功能性化合物进行接枝,改善丝蛋白材料的应用特性。目前,淀粉接枝[4-5]、黄麻改性[6-7]、丝蛋白功能化[8-10]主要采用物理和化学方法。物理法对设备投资大、处理要求高,化学法存在环境污染、对纤维损伤大等问题,因此,选用绿色环保的酶法改性[11]成为近年来纺织领域探究的重要课题。
辣根过氧化物酶(HRP)是一种植物过氧化物酶,从常年生香草辣根中提取,含量丰富,是商品化较早、应用广泛的一种酶制剂[12-13]。辣根过氧化物酶性质较稳定,具有高活性、高稳定性、低分子质量和存储方便等优点,在生物传感器[14]、有机合成[15-16]、环境废水处理[17]、食品工业等领域有着广泛的应用。
近年来,研究人员以HRP、过氧化氢(H2O2)和β-二酮类引发剂构建三元催化体系,开展了酶促纤维整理、丝蛋白基生物材料制备等相关研究[18]。本文介绍了HRP酶三元催化体系的氧化机制,并综述了其在淀粉浆料改性、麻纤维功能化及复合材料制备、丝素及丝胶分子修饰及其生物材料制备等方面的应用研究进展。
1 HRP的催化氧化机制
HRP是由辣根根部中多个同工酶组成的糖蛋白复合酶,其中辣根过氧化物酶同工酶C(HRP-C)是HRP酶的主要活性成分,且含量最大。HRP分子结构中含有2种不同类型的金属中心(高铁原卟啉环和2个钙原子),高铁原卟啉环决定了HRP-C的催化效能,而钙原子则影响着酶结构的稳定性,钙原子的缺失会降低HRP-C的活性和热稳定性[19]。
HRP有很强的催化活性,在以H2O2为氧化剂,β-二酮为引发剂的体系中,可催化乙烯类、苯胺类、酚类单体及其衍生物聚合,生成具有特殊结构的聚合物[20-21]。不同β-二酮对HRP催化中乙烯类单体的聚合物产率和分子质量有影响,其中乙酰丙酮(ACAC)的引发效果最好,可形成有效酶氧化还原催化体系,产生活性自由基,引发上述单体及其衍生物之间发生聚合反应。
图1示出HRP/H2O2/ACAC催化机制。该三元体系的催化过程是一个循环的氧化还原反应过程,主要分为3个步骤:首先,HRP与H2O2发生双电子氧化反应,即Fe(Ⅲ)和卟啉环各失去1个电子,HRP失去2个电子生成高价态的中间体 HRP-I;其次,HRP-I与还原底物ACAC(如麻纤维、丝蛋白等)发生单电子氧化还原反应,生成ACAC活性自由基和部分氧化中间体HRP-Ⅱ;最后,HRP-II会被ACAC进一步还原到初始价态,并再次产生ACAC活性自由基。在上述整个催化体系循环过程中,生成的ACAC自由基可引发单体间的聚合反应,从而实现催化可产生自由基的底物材料与乙烯基单体接枝共聚,改善材料的应用性能[22-23]。

图1 HRP/ACAC/H2O2催化机制Fig.1 Catalytic mechanism of HRP/H2O2/ACAC
2 HRP催化淀粉改性及其应用性能
淀粉由直链淀粉和支链淀粉构成,分子含大量羟基,浆液黏度高,流动性差,对疏水性纤维黏附性能较差。为改善淀粉浆料的性能,需要对淀粉进行改性,常用方法有物理改性(如预糊化)、化学改性(如酯化)、酶法改性(如接枝共聚等)和复合改性(如交联-酯化)[24]。HRP三元体系也可催化淀粉进行生物法改性加工,通过引发淀粉分子链的葡萄糖单元产生活性自由基,与性能优良的单体接枝共聚,达到改善淀粉物理和化学性质的目的。图2 示出HRP催化淀粉接枝丙烯酸甲酯反应示意图。可以看出,HRP催化淀粉分子上的羟基生成活性自由基,与丙烯酸甲酯(MA)接枝共聚,通过引入柔性聚合物支链聚丙烯酸甲酯(PMA),可改善淀粉浆膜的脆性,提升淀粉浆膜的力学性能和对疏水性纤维的黏附性[25]。
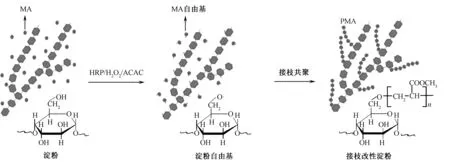
图2 HRP催化淀粉接枝丙烯酸甲酯反应示意图Fig.2 Schematic illustration of grafting reaction of MA onto starch catalyzed by HRP
Wang等[25]以HRP催化淀粉接枝MA反应后,以丙酮抽提去除其中的均聚物PMA,得到改性淀粉;改性淀粉的红外图谱上有羰基特征吸收峰出现,表明酶催化糊化淀粉与MA发生了接枝共聚反应,借助1H核磁共振谱和13C核磁共振谱测试分析,验证了MA成功接枝到淀粉分子上,且淀粉中葡萄糖单元的C6伯羟基为主要的接枝位点。采用粗纱浸浆法,测试HRP催化淀粉接枝改性前后浆液的黏附性和浆膜的力学性能,结果表明,HRP法催化接枝改性后的淀粉浆液对涤纶/棉粗纱的黏附性明显提高,且接枝率越高,黏附性能越好;随着接枝率增加,浆膜的断裂伸长率越大,表明浆膜的力学性能得到改善[26]。
3 HRP催化黄麻改性及复合材料制备
麻纤维中木质素含量较高,其基本结构单元为含酚羟基的愈创木基和紫丁香基[27]。HRP/H2O2/ACAC三元体系可催化氧化麻纤维,促进麻纤维中木质素的酚羟基形成酚氧自由基,也可催化乙烯基单体形成乙烯基自由基,这为麻纤维基材料改性提供了新方法,因此,麻纤维不仅可通过脱胶和练漂等工序加工成印染制品;未经脱胶处理的麻纤维也可通过纤维上木质素位点进行酶法接枝改性,加工成不同复合材料(包括黄麻纤维与热塑性树脂复合等),拓展了麻纤维的应用范围。
3.1 HRP催化黄麻纤维亲水化整理
利用HRP酶可催化黄麻纤维表面的木质素产生酚氧自由基的特性,实现黄麻与亲水性单体丙烯酰胺(AM)接枝共聚,从而改善黄麻纤维的亲水性能[28]。HRP催化黄麻接枝AM接枝改性反应示意图如图3所示。工艺为:采用HRP酶活力为 6 U/mL,AM用量为100%(o.w.f),ACAC体积分数为1%,通入氮气30 min后逐滴加入H2O2,在温度为 50 ℃、pH值为7的条件下反应5 h。经酶促反应后,通过水洗去除可能附着在麻纤维表面的自聚物聚丙烯酰胺(PAM),再测试织物的润湿时间、静态接触角和平衡回潮率。Liu等[28]研究表明,接枝样表面滴水润湿时间和静态接触角明显比对照样的滴水润湿时间短、接触角小。与对照样的平衡回潮率相比,接枝样的平衡回潮率较大,说明在HRP/H2O2/ACAC的催化作用下,黄麻织物表面接枝上了AM,引入了亲水基团—CONH2,使黄麻织物亲水性能有所提高。接枝改性黄麻织物红外谱图中出现酰胺基团特征吸收峰,扫描电镜照片中黄麻纤维表面更粗糙,有絮状物质的存在,也验证了HRP催化黄麻与AM发生了接枝共聚反应。
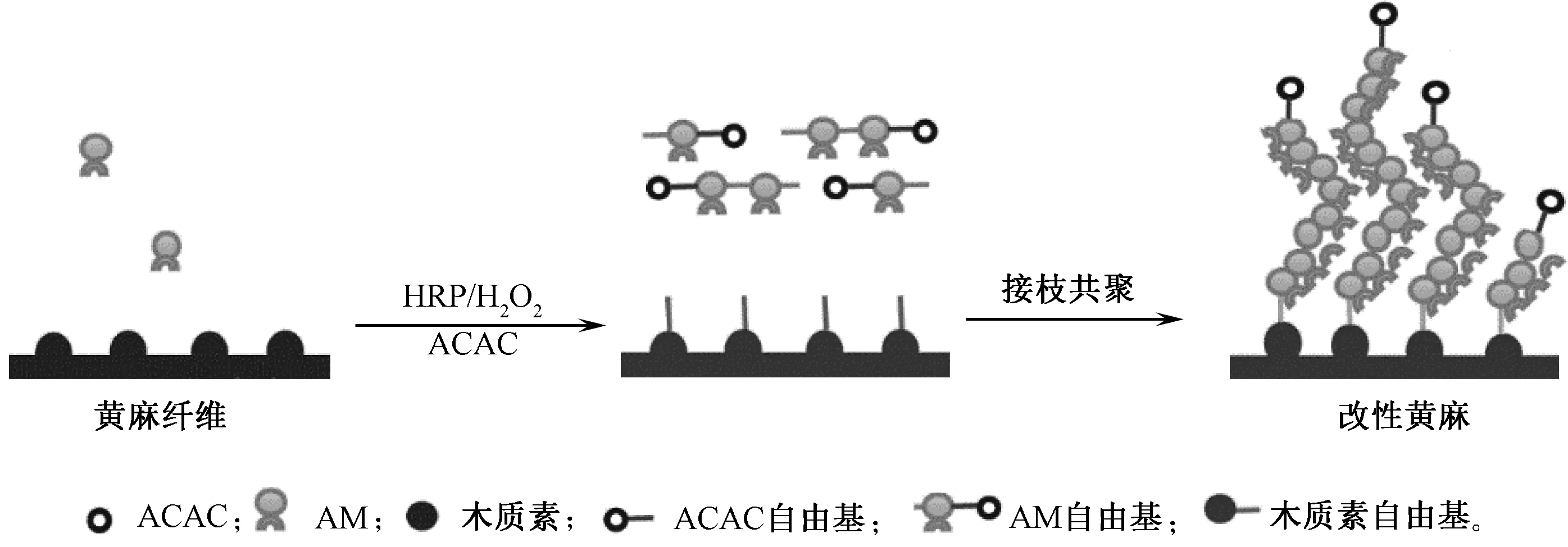
图3 HRP催化黄麻纤维中木质素与丙烯酰胺接枝共聚反应示意图Fig.3 Schematic illustration of HRP-catalyzed graft copolymerization of AM onto lignin of jute fabrics
3.2 HRP催化黄麻纤维疏水化整理
利用HRP酶催化黄麻表面的木质素产生酚氧自由基,将疏水性乙烯基单体(如丙烯酸丁酯、甲基丙烯酸六氟丁酯)接枝到黄麻纤维表面,可实现麻织物疏水化整理[29]。图4示出HRP催化疏水性乙烯基单体和黄麻接枝共聚反应式。ACAC在HRP/H2O2体系中形成自由基,电子转移至单体的乙烯基以及木质素的酚羟基后,通过自由基反应使木质素与疏水性单体发生接枝聚合。
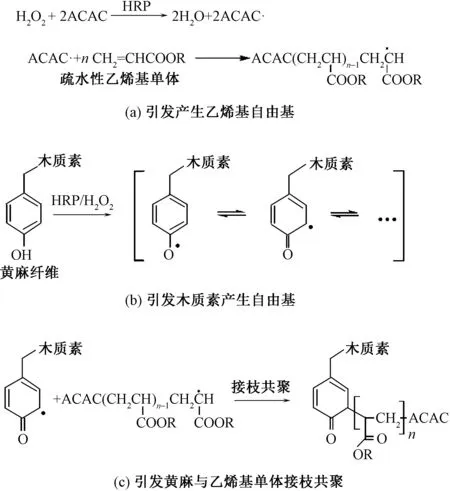
图4 HRP催化疏水性乙烯基单体和黄麻接枝共聚Fig.4 HRP-catalyzed graft copolymerization of hydrophobic vinyl monomers onto jute.(a) Generation of free radicals from vinyl monomers; (b) Generation of free radicals from lignin in jute; (c) Graft copolymerization of vinyl monomers onto jute
Wu等[29]选择甲基丙烯酸六氟丁酯(HFBMA)作为高疏水性单体,HFBMA经乳化后,以HRP/H2O2/ACAC催化其在黄麻纤维表面接枝共聚,反应后以丙酮抽提去除黄麻织物表面附着的HFBMA自聚物,得到疏水化改性黄麻。吴慧敏[30]采用X射线光电子能谱仪测定未处理样和疏水化改性黄麻纤维表面的C、O、N和F元素含量发现,C与O的量比从1.551增加到1.938,氟元素含量从0.12%增加到29.41%,验证了HFBMA及其聚合物在黄麻表面发生接枝共聚;测定HRP催化接枝后黄麻织物的静态接触角和润湿时间发现,织物表面接触角较大,润湿时间也较长,表明接枝后的黄麻织物具有较高的疏水性,验证了三元体系可用于黄麻纤维疏水化改性加工。
3.3 HRP催化黄麻纤维基复合材料制备
天然纤维基增强复合材料具有生态环保的特性,其中麻纤维因具有产量大、模量高的特点,使其在增强复合材料制备方面具有很大的应用前景。但由于麻纤维中含有大量羟基,使得其与非极性树脂之间的界面黏结性较差,影响纤维基复合材料的应用性能。刘锐锐等[31]选用HRP/H2O2/ACAC三元体系,通过催化黄麻纤维接枝3种没食子酸酯(没食子酸辛酯、没食子酸月桂酯和没食子酸十八酯),并与聚丙烯(PP)进行复合,制备了黄麻/聚丙烯基复合材料并测定其拉伸性能。结果表明,未改性黄麻/PP复合材料拉伸强力和拉伸模量最低,接枝没食子酸酯黄麻/PP复合材料的拉伸模量、拉伸强力明显提高,说明黄麻表面疏水性能有效提高了黄麻/PP复合材料的拉伸性能。这是因为黄麻通过接枝改性,在其结构中引入了没食子酸酯单体的疏水长碳链,其与疏水聚丙烯分子之间产生强作用力,提高了黄麻纤维与树脂间的界面相容性,从而使黄麻/PP复合材料力学性能提高[31]。
4 HRP催化蚕丝改性及再生材料制备
与多酚氧化酶类似,HRP三元体系也可催化氧化丝蛋白(桑蚕丝、丝胶或丝素)中酪氨酸剩基,使酪氨酸剩基中含酚羟基的苯环上产生自由基,引发丝蛋白分子自交联或与外源乙烯基功能性单体接枝聚合,实现丝蛋白材料的生物法功能化。
4.1 HRP催化丝素蛋白分子自交联
Zhou等[32]探究了HRP/H2O2体系催化氧化丝素蛋白的反应机制。首先,选择对羟基苯乙酰胺(PHAD)为酪氨酸的模型物,借助核磁、质谱等测试手段验证了PHAD可被催化氧化,引发聚合形成二聚体、三聚体和四聚体等。在此基础上,结合凝胶渗透色谱凝胶柱检测丝素蛋白分子质量的变化,HRP/H2O2处理后样品的出峰时间较未处理试样提前,验证了丝素蛋白分子间发生交联。HRP催化丝素蛋白分子交联的反应途径如图5所示。丝素中酪氨酸的酚羟基被氧化产生自由基,随后引发丝素蛋白中酪氨酸残基之间的酶促聚合。Zhou等[32]考察了HRP/H2O2处理后丝素冻干膜在热水中的溶失率。结果表明,丝素膜空白样于37 ℃振荡处理1 h后几乎全部溶解,HRP/H2O2处理丝素膜溶失率低于60%,验证了丝素分子质量增加,膜材料的溶失率随之降低。
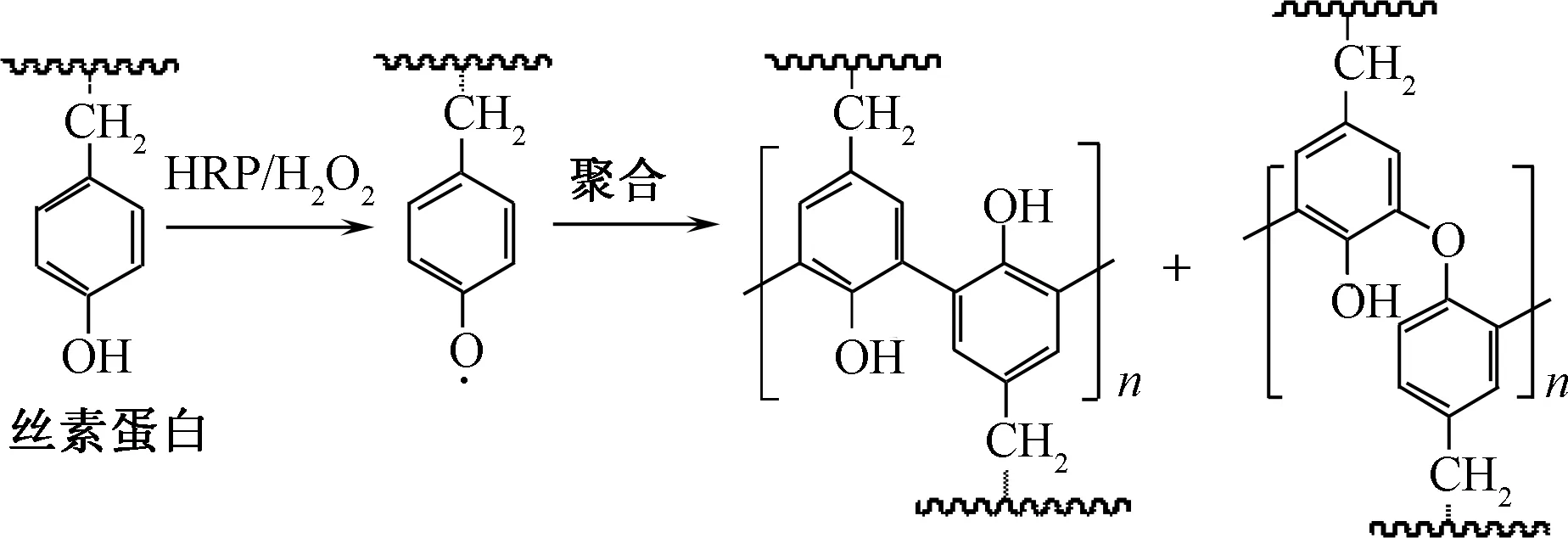
图5 HRP催化丝素蛋白分子间交联Fig.5 Intermolecular cross-linking of silk fibroins initiated by HRP
4.2 HRP催化丝素接枝丙烯酶及其仿生矿化
HRP三元体系可催化丝素蛋白中酚羟基与丙烯酸等乙烯基单体接枝共聚,制备丝素基生物材料。Zhou等[33]以丝素蛋白为对象,借助HRP催化丙烯酸(AA)在丝素表面接枝共聚,反应后将产物冻干并醇化处理,以蒸馏水洗涤去除丙烯酸自聚体(PAA)后,得到接枝有聚丙烯酸的丝素基复合膜材料(SF-g-PAA),其反应示意图如图6所示。SF-g-PAA 中负电性的羧基对钙离子有较强吸附和螯合作用,有利于仿生矿化中结合钙离子,促进磷酸钙盐的沉积和羟基磷灰石(Ca10(PO4)6(OH)2,简称HAp)的形成,为基于骨组织诱导的丝素基仿生矿化材料的构建提供了新方法。
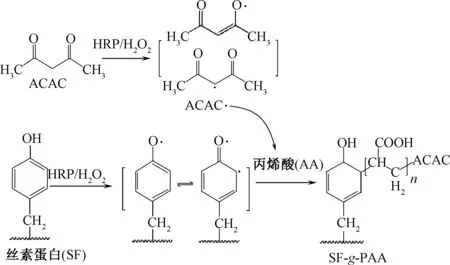
图6 HRP催化丝素与丙烯酸接枝共聚形成SF-g-PAA示意图Fig.6 Schematic illustration of SF-g-PAA produced by HRP-catalyzed graft copolymerization of AA onto silk fibroins
Zhou等[33]借助傅里叶变换衰减全反射光谱分析交替矿化后SF-g-PAA膜材料的结构,考察HAp晶体在材料表面的生长情况;SF-g-PAA在5个矿化周期后,就检测到PO43-的反对称伸缩振动特征谱带;交替浸渍矿化达到10个周期,HAp在SF-g-PAA表面的峰强度显著高于丝素膜矿化样,表明丝素与AA接枝共聚有利于HAp的矿化生成;借助X射线衍射仪对不同矿化周期下SF-g-PAA/HAp复合材料进行分析,经交替矿化20个周期后,试样的无定形结构非晶峰包几乎消失,成骨细胞MG-63在该复合材料表面具有良好的黏附和生长效果[33]。由此可见,采用HRP催化丝素蛋白接枝AA,有利于后续仿生矿化中HAp的成核和结晶,拓展了丝素蛋白作为医用生物材料的应用前景。
4.3 HRP催化丝胶接枝MMA及其成形性
丝胶多来自于桑蚕丝脱胶,传统意义上由于其水溶性较高,因此,多作为废弃物排放到印染废水中。随着人们对丝胶结构、性能、生物活性及功能(抗菌、抗氧化等)认识的逐步深入发现,丝胶在日化及生物材料领域有潜在用途。以丝胶制作生物材料,存在成形性较差的缺陷,采用HRP三元体对酶促丝胶蛋白交联、丝胶与疏水性单体甲基丙烯酸甲酯(MMA)接枝共聚进行相关研究。He等[34]研究表明,与未处理丝胶相比,HRP处理后丝胶分子质量增加,制备的丝胶/聚甲基丙烯酸甲酯复合材料(SS-g-PMMA)不仅水溶性较低,成形性改善,且NIH/3T3细胞在样品中的存活率较高,表明改性丝胶膜有良好生物相容性。
5 结束语
辣根过氧化物酶具有高效、专一和反应条件温和的催化特性,HRP/H2O2及HRP/H2O2/ACAC三元催化体系在纺织浆料、麻纤维改性、桑蚕丝功能化、丝素和丝胶蛋白分子改造与再生生物材料制备等领域均具有广泛的用途。HRP酶法不仅反应效率较高,且反应条件温和,对环境影响较小,为纤维整理和生物材料的制备提供了的新途径。
