IL-1β介导脓毒症大鼠血管α1肾上腺素能受体失敏机制的研究*
2019-04-25梁家林刘良明
梁家林,刘良明,董 惠
(1.空军杭州特勤疗养中心,杭州 310013;2.陆军军医大学大坪医院野战外科研究所二室/创伤、烧伤与复合伤国家重点实验室,重庆 400042;3.陆军军医大学免疫研究所,重庆 400038)
尽管研究不断深入,但脓毒症发病率和病死率始终居高不下[1],血管低反应性(即血管对血管活性物质的反应性降低)是致使其高病死率的重要原因[2]。
研究证实,白细胞介素(IL)-1β介导脓毒症血管低反应性发生,含NO依赖和非依赖性机制[3],但针对上述机制进行纠正却只能部分恢复血管反应性[4-5],提示可能存在其他机制。基于α1肾上腺素能受体(α1ARs)在心血管系统中的重要作用,目前认为α1ARs失敏(主要是表达下降)是导致脓毒症后难治性低血压发生的重要原因[6]。笔者前期研究发现IL-1β能通过JAK2-STAT3途径介导脓毒症休克家兔血管α1ARs失敏[7]。另有研究显示,IL-1β通过核因子-κB(NF-κB)途径在转录而不是转录后水平降低小鼠血管α1ARs的表达[8]。由于α1ARs的表达调控具有种属和组织的差异性[9],多种属作用机制的研究有利于阐述其详细作用机制,本文重点研究IL-1β对脓毒症大鼠α1ARs的表达调控机制。
目前已经克隆出大鼠血管平滑肌细胞(VSMCs)的α1ARs各亚型(α1AAR、α1BAR、α1DAR)的启动子区域,其中相同的转录因子结合区域仅有Sp1和AP2[10-12]。笔者前期研究显示:IL-1β对VSMCs α1ARs各亚型表达均有类似幅度的下调作用[7],推测IL-1β可能通过Sp1和AP2在转录水平调控VSMCs α1ARs表达,另外也研究IL-1β是否通过改变α1ARs mRNA半衰期在转录后水平调控VSMCs α1ARs表达。
1 材料与方法
1.1实验动物 体质量(200±10)g的SPF级健康成年雌性SD大鼠由陆军军医大学实验动物中心提供,通过陆军军医大学伦理委员会审批。
1.2主要试剂与仪器 重组大鼠IL-1β(美国Pepro Tech公司),兔抗大鼠α1AAR抗体(美国Epitomics公司),山羊抗大鼠α1BAR、α1DAR抗体(美国Santa Cruz公司),兔抗大鼠Sp1、AP2抗体(美国Pierce公司),放线菌素D(美国Invitrogen公司),X-tremeGENE HP DNA 转染试剂(美国Roche公司),双萤光素酶定量试剂盒(美国Gene Copoeia公司),Power Lab八道生理记录仪(澳大利亚AD Instrument公司),垂直板状电泳仪(美国Bio-Rad公司),CO2细胞培养箱(美国SHEL/JB公司),ABI Prism 7700(美国Perkin Elmer公司)。
1.3方法
1.3.1脓毒症模型的复制及指标检测 将32只大鼠分为假手术组、经盲肠结扎穿孔(CLP)3 h组、CLP 6 h组及CLP 12 h组,每组8只。采用CLP复制脓毒症模型,手术操作、分组处理和血管环制作同文献[13],测定各组血管环的α1ARs敏感性[对去氧肾上腺素(PE)的反应性][7];另游离出各组肠系膜上动脉(SMAs)分支,用RIPA裂解液提取其总蛋白,用蛋白质印迹法(Western blot)检测α1ARs、β-actin表达。分析上述指标与IL-1β浓度的关系。
1.3.2IL-1β对大鼠VSMCs α1ARs、Sp1、AP2表达水平的影响 原代培养SMAs来源的VSMCs[13],实验分为对照组、1 ng/mL IL-1β组、10 ng/mL IL-1β组和100 ng/mL IL-1β组,每组4瓶细胞。IL-1β孵育VSMCs方法同文献[13],用RIPA裂解液提取各组细胞的总蛋白,用Western blot检测α1ARs、Sp1、AP2、β-actin表达。
1.3.3IL-1β对大鼠VSMCs α1ARs mRNA半衰期的影响 IL-1β孵育VSMCs方法同前,在对照组或100 ng/mL IL-1β孵育12 h后加入放线菌素D(终浓度为4 μg/mL)[14],依次加入放线菌素D后0、2、4、6 h后Realtime-PCR检测α1ARs mRNA表达水平。所得Cq值结合标准曲线得到各组的量。引物序列见表1。
1.3.4Sp1和AP2 siRNA对大鼠VSMCs α1ARs表达的影响 实验分为空白对照组、FAM siRNA(阴性对照)组、AP2γ siRNA组、AP2α siRNA组、Sp1 siRNA组,每组4瓶细胞。细胞准备:取融合度40%左右的原代培养的大鼠VSMCs,PBS清洗2次,加入1 800 μL含20%胎牛血清的DMEM F/12培养基。配制转染复合物:193 μL DMEM F/12培养基,2 μL X-tremeGENE HP DNA,5 μL 20 mol/L的siRNA溶液(空白对照组加入5 μL无酶水),室温混匀放置15 min。孵育细胞:将转染复合物加入各组细胞后细胞培养箱中培养72 h,RIPA裂解液提取各组总蛋白,Western blot检测Sp1、AP2、β-actin及α1ARs表达水平。Sp1:正义siRNA 5′-GGA GCG AUC AUC UGU CAA ATT-3′,反义siRNA 5′-UUU GAC AGA UGA UCG CUC CTT-3′;AP2γ:正义siRNA 5′-UGU CAC CAC CGG AAU GCU UTT-3′,反义siRNA 5′-AAG CAU UCC GGU GGU GAC ATT-3′;AP2α:正义siRNA 5′-GGA GAG CGA AGU CUA AGA ATT-3′,反义siRNA 5′-UUC UUA GAC UUC GCU CUC CTT-3′;FAM:正义siRNA 5′-UUC UCC GAA CGU GUC ACG UTT-3′,反义siRNA 5′-ACG UGA CAC GUU CGG AGA ATT-3′。
1.3.5IL-1β对大鼠VSMCs α1DAR启动子活性的影响 实验分为对照组和100 ng/mL IL-1β 组,每组6瓶细胞。克隆α1DAR启动子并构建入质粒(GeneCopoeia公司代做),扩增质粒,用X-tremeGENE HP DNA转染试剂将质粒转染入VSMCs(转染方法基本同1.3.4),加入100 ng/mL(终浓度)IL-1β孵育,对照组仅加入转染试剂,24 h后吸取两组培养基,dual luciferase assay试剂盒按说明书操作检测两组培养基

表1 引物序列
与Gaussia萤光素酶(gaussia luciferase,GLUC)和分泌性碱性磷酸酶(alkaline phosphatase,SEAP)反应产生的光强度,GLUC/SEAP光强度即为启动子活性。

2 结 果
2.1脓毒症大鼠α1ARs敏感性和各亚型蛋白表达及与血清IL-1β的关系 与假手术组相比,在CLP后3、6和12 h,大鼠SMAs的α1ARs敏感性和各亚型蛋白表达均明显降低(P<0.05),见图1。笔者前期研究结果显示血浆IL-1β水平在CLP后6 h明显升高,相关性分析显示其与同时期SMAs的α1ARs敏感性及蛋白表达水平呈明显负相关(r=-0.823、-0.864,P<0.05)。

a:P<0.05,与假手术组比较
图1 脓毒症大鼠α1ARs敏感性和蛋白表达变化
2.2IL-1β对大鼠VSMCs α1ARs、AP2和Sp1表达的影响 与对照组相比,各浓度IL-1β组大鼠VSMCs α1ARs各亚型的蛋白表达均明显降低(P<0.05),而且呈浓度依赖性降低。各浓度IL-1β均能明显降低AP2和Sp1表达(P<0.05),见图2。
2.3IL-1β对大鼠VSMCs α1ARs mRNA稳定性及启动子活性的影响 与对照组比较,100 ng/mL IL-1β组大鼠VSMCs α1ARs各亚型的mRNA稳定性无明显改变(P>0.05)。100 ng/mL IL-1β能明显降低大鼠VSMCs α1DAR启动子活性(P<0.05),见图3。

a:P<0.05,与对照组比较
图2 IL-1β对大鼠VSMCs α1ARs、AP2和Sp1表达的影响
2.4Sp1和AP2对大鼠VSMCs α1ARs表达的影响 空白对照组和阴性对照组间α1ARs表达无明显差异(P>0.05)。与空白对照组或阴性对照组相比,Sp1 siRNA、AP2α siRNA和AP2γ siRNA组均能明显降低VSMCs α1ARs各亚型的蛋白表达(P<0.05),见图4。
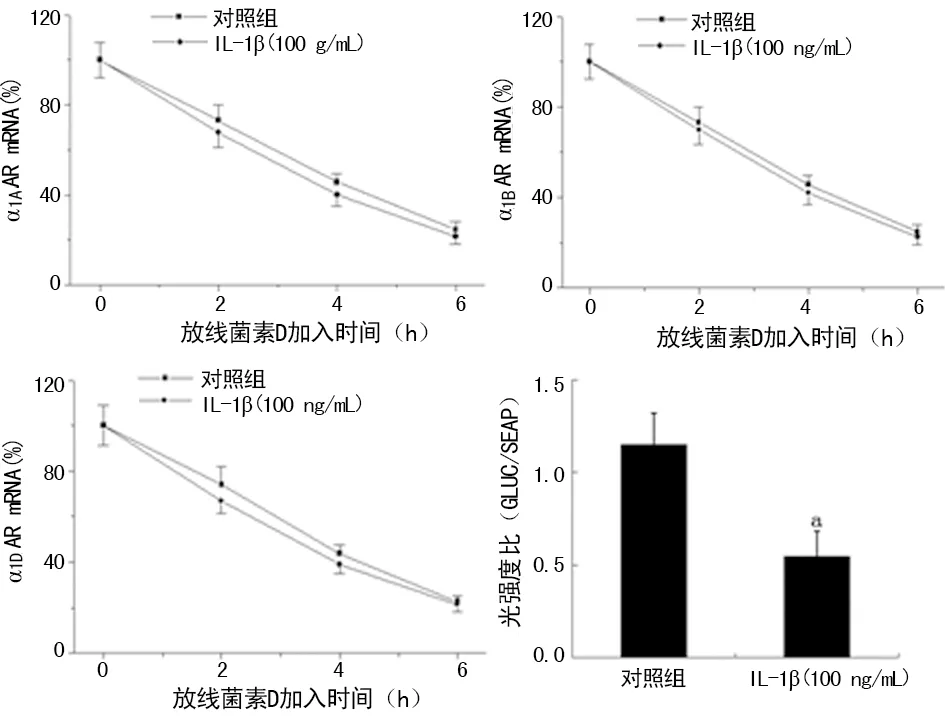
a:P<0.05,与对照组比较
图3 IL-1 对大鼠VSMCs α1ARs mRNA稳定性及α1DAR启动子活性的影响

a:P<0.05,与空白对照组比较;b:P<0.05,与阴性对照组比较
图4 Sp1、AP2 siRNA对大鼠VSMCs α1ARs表达的影响
3 讨 论
尽管对脓毒症的认知水平有明显提高,但在全球范围内,其发病率、病死率均较高。鉴于α1ARs在心血管系统中的重要作用,它在脓毒症的发生、发展和转归中具有重要地位。
研究显示脓毒症时存在血管α1ARs失敏,主要是受体减少,亲和力无明显改变[8]。体内外研究均显示IL-1β能下调脓毒症时血管α1ARs表达量[6-7],但是对其调控机制目前不甚清楚。
本研究通过CLP复制脓毒症大鼠模型,整体水平验证脓毒症时存在α1ARs失敏且与IL-1β相关。结果显示CLP 3 h后即存在α1ARs失敏和表达量进行性下降,但α1ARs亲和力无明显改变,而血浆IL-1β却在CLP 6 h后开始升高[13]。IL-1β浓度升高晚于α1ARs失敏,可能因为脓毒症早期其他细胞因子如肿瘤坏死因子α(TNF-α)对α1ARs失敏也有作用。而在IL-1β升高后α1ARs进一步失敏,相关性分析显示二者存在明显负相关。为排除脓毒症时其他细胞因子的干扰,笔者在离体时采用重组大鼠IL-1β孵育大鼠VSMCs,观察其对α1ARs表达的影响。结果显示IL-1β能明显降低α1ARs各亚型的表达。整体和离体水平验证了IL-1β通过下调α1ARs各亚型的表达介导脓毒症α1ARs失敏的发生。
尽管已有研究显示IL-1β通过下调小鼠血管α1ARs启动子活性在转录而不是转录后水平下调α1ARs的表达。但现有研究提示α1ARs各亚型的表达、分布和调控机制具有种属和组织的差异性,IL-1β对小鼠的作用机制是否适用于大鼠甚至人类,目前不得而知。通过运用转录抑制剂(放线菌素D),发现IL-1β并不能改变大鼠VSMCs α1ARs各亚型 mRNA的半衰期,提示IL-1β也可能是通过转录而不是转录后介导大鼠血管α1ARs表达下降。为验证这一结论,笔者随机选择检测α1DAR启动子活性,结果显示IL-1β能明显下调大鼠VSMCs α1DAR启动子活性,尽管没有检测α1AAR和α1BAR启动子活性,但仍然可以得到IL-1β通过转录水平介导大鼠VSMCs α1ARs表达下降这一结论。
目前α1ARs各亚型在大鼠VSMCs的启动子区域已经克隆出来, α1AAR主要有Sp1、NF-κB、NF/IL-6、AP1、CTF/NF1、AP2等转录因子的结合区域[10],α1BAR主要有Sp1、HNF-1/5、AP2、NF1、CREB、CP1等的结合区域[11],而α1DAR则主要有Sp1和AP2的结合区域[12]。它们都有Sp1和AP2的结合区域,上述结果提示IL-1β对大鼠VSMCs α1ARs各亚型的表达存在相同的下调作用,且下调趋势基本一致,为此笔者推测IL-1β可能通过相同的转录因子(Sp1和AP2)发挥调节作用。笔者检测了IL-1β对大鼠VSMCs Sp1和AP2表达的影响,发现IL-1β能明显降低Sp1和AP2的蛋白表达。Sp1和AP2的蛋白表达下调是否引起α1ARs的表达下降呢?通过运用Sp1和AP2(含AP2α和AP2γ)siRNA,发现Sp1和AP2表达下降使得α1ARs各亚型表达下降。由于α1AAR和α1BAR还有其他转录因子结合区域,也可能在IL-1β介导的脓毒症大鼠血管α1ARs失敏中发挥作用。鉴于本研究得出Sp1和AP2在脓毒症大鼠血管α1ARs失敏中的重要作用,值得进一步的研究,未来可以考虑作为脓毒症的治疗靶点。
综上所述,IL-1β可能通过下调大鼠VSMCs 的Sp1和AP2表达,在转录而不是转录后水平下调α1ARs表达,介导脓毒症大鼠血管α1ARs失敏的发生。
