SM22α对结直肠癌细胞侵袭和迁移能力的影响*
2019-04-22刘亚彬王光林
赵 斌, 刘亚彬, 王光林
(河北医科大学附属哈励逊国际和平医院普外二科, 河北 衡水 053000)
结直肠癌(colorectal cancer,CRC)是消化系统中常见的恶性肿瘤之一[1],近年来随着人们物质生活水平的提高,饮食习惯的改变,结直肠癌的发病率逐渐升高,结直肠癌发病率位于欧美发达国家恶性肿瘤第3位,死亡率则高居第2位[2]。在我国,结直肠癌发病率和死亡率居于恶性肿瘤中第5位[3],严重影响着人们的健康水平。目前对于结直肠癌患者多采取手术治疗的方式,可以获得一定的效果,但是对于转移性结直肠癌患者来说目前的治疗方法仍是难点,因此,深入探讨结直肠癌细胞侵袭和迁移的机制以及针对其生物学特点开发出有效的靶向干预手段十分重要。
细胞外信号调节激酶(extracellular signal-regu-lated kinase,ERK)是丝裂原激活蛋白激酶(mitogen-activated protein kinase,MAPK)家族重要成员,p-ERK是其主要发挥功能的形式,p-ERK通过增强转录因子磷酸化而增强其转录活性,调控细胞增殖、凋亡、分化和恶性肿瘤侵袭转移等。基质金属蛋白酶9(matrix metalloproteinase-9,MMP-9)是ERK信号通路的下游因子,通过降解细胞外基质中的蛋白成分,调节恶性肿瘤的侵袭和转移行为[4]。
平滑肌22α蛋白(smooth muscle 22α protein,SM22α)是细胞骨架蛋白中的一种,其序列上的高度保守性提示这种蛋白具有相当重要的作用[5]。既往研究发现,SM22α能够与肌动蛋白结合,调节血管平滑肌的收缩,此外还具有稳定和调节细胞骨架的作用。近期的研究显示,SM22α可能作为抑癌基因参与多种肿瘤的发生发展过程,SM22α通过抑制ERK信号通路调节MMP-9的表达活性[6],在肿瘤侵袭和转移过程中发挥一定的作用。然而SM22α与结直肠癌侵袭和迁移之间的关系及机制研究仍未报道。本研究旨在体外验证SM22α对结直肠癌侵袭和迁移的影响,探究ERK/MMP-9信号通路在其中发挥的功能,为结直肠癌患者的诊断和治疗提供新思路。
材 料 和 方 法
1 材料
人结直肠癌细胞HCT116购自ATCC;RPMI-1640培养基购自Gibco;胎牛血清购自杭州四季青公司;PD98059购自Sigma-Aldrich;SM22α过表达慢病毒由上海吉凯基因化学技术有限公司构建、筛选和包装;RNA提取试剂盒购自上海飞捷生物技术有限公司;兔抗人ERK、p-ERK和MMP-9抗体购自CST;兔抗人SM22α抗体购自Santa Cruz;鼠抗人β-actin抗体购自沈阳万类生物科技有限公司。
2 方法
2.1细胞培养和慢病毒转染 HCT116细胞培养于含有10%胎牛血清的RPMI-1640培养基中,置于37 ℃、体积分数为5%的CO2细胞培养箱中培养。培养2~3 d后用胰酶消化传代,按1 ∶2或1 ∶3比例进行传代,取生长状态良好处于对数生长期的细胞用于实验。人结直肠癌细胞HCT116正常培养传代,SM22α过表达慢病毒预实验感染人结直肠癌细胞HCT116,确定慢病毒的MOI值。重悬调整细胞浓度后接种于6孔板中,转染前用PBS清洗细胞,在人结直肠癌细胞中加入5 mg/L的polybrene和相应体积的慢病毒转染细胞48 h,使用2.5 mg/L嘌呤霉素进行筛选2~3周,荧光显微镜下检测荧光蛋白的表达,RT-qPCR和Western blot检测SM22α的mRNA和蛋白表达。
2.2Transwell迁移和侵袭实验 取生长状态良好处于对数生长期的对照组及SM22α过表达组HCT116细胞用胰酶消化离心,无血清RPMI-1640培养基重悬细胞并计数,吸取200 μL细胞悬液(每室2.0×104和4.0×104)加入上室(侵袭实验4 h前在小室上室铺50 μL稀释Matrigel),下室加入800 μL含10%胎牛血清的RPMI-1640培养基(每组设置3个副室);置于温度为37 ℃、体积分数为5%的CO2培养箱内培养,36 h取出Transwell小室。用4%的多聚甲醛固定15 min,0.1%的结晶紫染色15 min,使用棉签擦拭干净上室未穿膜细胞,在倒置显微镜下随机观察并计数5个视野下的细胞数目。
2.3细胞划痕实验 细胞接种于6孔板中,待细胞密度生长至90%~100%左右使用200 μL无菌枪头沿直尺垂直进行细胞划痕,PBS清洗悬浮脱落细胞并取划痕处进行拍照(0 h),更换无血清培养基继续培养,分别于6 h、12 h和24 h在同一划痕位置进行拍照。精确测量划痕平均间距,计算细胞迁移率和相对愈合率并分析实验结果。
2.4MTT实验 取对数生长期的对照组及SM22α过表达组HCT116细胞,按每孔5.0×103接种于6孔板,对照组和SM22α过表达组细胞各设5个复孔,细胞置于37 ℃、5% CO2培养箱继续培养。加入含10% MTT的培养基培养4 h,吸去上清液并加入150 μL DMSO,振荡器振荡混匀10 min,酶联免疫检测仪在490 nm波长处检测不同处理组吸光度(A值),绘制细胞生长曲线。
2.5Western blot实验 将处于对数生长期的细胞用胰酶消化离心后,重悬成细胞悬液并进行计数,以每孔2×105接种于6孔板中,置于37 ℃、体积分数为5%的CO2培养箱中培养,待细胞生长贴壁后取出并用4 ℃的PBS冲洗3次,每孔加入150 μL细胞裂解液(含PMSF)裂解细胞,将裂解下来的细胞吸入EP管中,放在涡旋振荡器上振荡30 s,冰上静置10 min,然后于4 ℃、15 000 r/min高速离心15 min,收集上清液中的总蛋白。吸取5 μL上清液做蛋白定量,余下上清液内加入6×loading buffer,在沸水中煮沸5 min,使蛋白质变性。每组吸取30 μg蛋白进行SDS-PAGE,待蛋白分离后,转移到PVDF膜上,用含5%脱脂牛奶的TBST室温封闭1 h,加入 I 抗抗体4 ℃孵育过夜,次日经TBST洗膜3次,每次10 min,加辣根过氧化物酶标记的 II 抗室温孵育1 h,再用TBST洗膜液3次,每次10 min,使用ECL化学发光法显影,目的蛋白通过与内参照蛋白β-actin标准化后得到相对比值。
2.6RT-qPCR实验 将处于对数生长期的细胞进行消化离心后,细胞传代于6孔板中,待细胞生长至80%~90%时按照飞捷RNAfast200试剂法提取RNA。从提取的RNA中吸取2 μL做RNA定量,并通过反转录得到稳定的cDNA模板,反转录体系参照TaKaRa PrimeScriptTMRT Master Mix试剂说明书。SM22α 的上游引物序列为5’-AGTGCAGTCCAAAATCGAGAAG-3’,下游引物序列为5’-CTTGCTCAGAATCACGCCAT-3’。使用β-actin作为内参照,其上游引物序列为5’-CATGTACGTTGCTATCCAGGC-3’,下游引物序列为5’-CTCCTTAATGTCACGCACGAT-3’。采用2-ΔΔCt计算方法进行分析。
3 统计学分析
通过SPSS 13.0进行统计分析。计量资料以均数±标准差(mean±SD)表示。组间均数比较采用单因素方差分析(one-way ANOVA)及LSD-t检验。以P<0.05表示差异有统计学意义。
结 果
1 构建HCT116过表达SM22α细胞系
应用SM22α过表达慢病毒感染HCT116细胞,构建SM22α稳定转染过表达细胞,使用Western blot和RT-qPCR检验SM22α蛋白和mRNA的表达,证实成功构建SM22α过表达细胞,见图1。

Figure 1.Construction of the SM22α-over-expressing HCT116 cells. A: Western blot was used to detect the protein expression of SM22α; B: RT-qPCR was applied to test the mRNA level of SM22α. Mean±SD.n=3.**P<0.01vscontrol group.
图1构建过表达SM22α的HCT116细胞
2 MTT实验检测SM22α对细胞活力的影响
与对照组相比,24 h和48 h细胞活力的差异无统计学显著性,72 h细胞活力的差异有统计学意义(P<0.05)。 SM22α过表达抑制细胞活力(P<0.05),见图2。
3 SM22α抑制HCT116细胞的迁移能力
将对照组和SM22α过表达组HCT116细胞接种于6孔板,应用细胞划痕实验检测细胞迁移能力的变化,结果发现SM22α过表达后HCT116细胞的迁移能力减弱,24 h划痕愈合率差异存在统计学意义(P<0.01),说明SM22α过表达抑制HCT116细胞迁移,见图3。
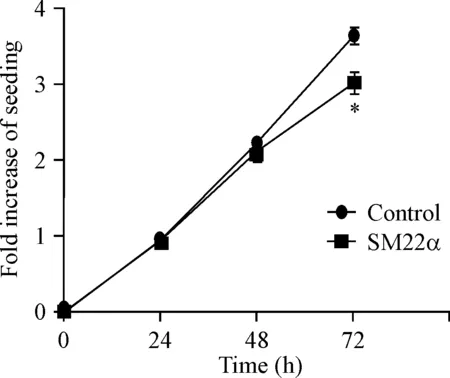
Figure 2.The effect of SM22α overexpression on cell viability. Mean±SD.n=3.*P<0.05vscontrol group.
图2SM22α过表达对细胞活力的影响

Figure 3.The cell migration ability was detected by cell scratch test. Mean±SD.n=3.**P<0.01vscontrol group.
图3细胞划痕实验检测细胞迁移能力
4 SM22α过表达抑制细胞迁移和侵袭
HCT116细胞对照组和SM22α过表达组分别行Transwell 迁移和侵袭实验。结果显示,SM22α过表达组与对照组相比HCT116细胞的迁移能力降低(P<0.01),在侵袭实验中SM22α过表达组侵袭能力减弱(P<0.05),说明SM22α抑制HCT116细胞迁移和侵袭,见图4。

Figure 4.The cell migration and invasion abilities were measured by Transwell expriments. Mean±SD.n=3.*P<0.05,**P<0.01vscontrol group.
图4Transwell实验检测细胞迁移和侵袭能力
5 SM22α过表达抑制ERK和MMP-9蛋白水平
Western blot 检测对照组和SM22α过表达组HCT116细胞MMP-9、p-ERK和ERK的蛋白水平。结果发现过表达SM22α抑制MMP-9和p-ERK的蛋白水平(P<0.01),见图5A。同时使用PD98059阻断ERK信号通路,检测HCT116细胞MMP-9蛋白水平的变化,发现阻断ERK信号通路后,过表达SM22α不能抑制MMP-9蛋白表达,见图5B。

Figure 5.The protein levels of ERK/MMP-9 were determined in the SM22α-over-expressing cells. A: the protein levels of MMP-9,ERK and p-ERK were detected by Western blot; B: ERK signaling pathway inhibitor PD98059 was used. Mean±SD.n=3.**P<0.01vscontrol group.
图5SM22α过表达细胞中ERK/MMP-9蛋白水平的变化
讨 论
近年来随着国人饮食结构的改变,结直肠癌的发病率和死亡率逐渐升高,且呈现年轻化的趋势,在临床上针对结直肠癌的治疗开发出了很多手段,如手术治疗、化疗和放疗等,靶向治疗主要用于治疗转移性结直肠癌[7-8]。结直肠癌具有较强的侵袭迁移能力,肝、肺、肾等远处转移是结直肠癌治疗的难点和重点[9-10]。因此,深入研究和探索影响结直肠癌侵袭迁移的分子机制,发现其潜在的治疗靶点具有重要的现实意义和临床价值。
研究表明,SM22α除了可以调节和稳定细胞骨架之外,还作为抑癌基因参与到肿瘤的发生发展及侵袭迁移等生物学过程当中,研究表明SM22α在前列腺癌[11]、结直肠癌[12]和肺癌[13]等肿瘤组织中表达降低。然而,在结直肠癌中关于SM22α对结直肠癌侵袭迁移能力的影响却未有报道,本研究主要探讨SM22α在结直肠癌中发挥的作用。通过慢病毒转染成功构建HCT116过表达SM22α细胞系,通过细胞划痕实验和Transwell小室实验发现SM22α过表达可抑制结直肠癌侵袭和迁移能力。
有研究发现SM22α可抑制MMP-9的表达,MMP-9在肿瘤组织中的表达明显高于正常组织[14],此外MMP-9是重要的细胞外基质蛋白水解酶,能降解多糖以外的所有细胞外基质成分,增强癌细胞的侵袭能力,促进肿瘤细胞侵入相邻组织并向远处转移。ERK是MAPK家族的重要组成,参与调节肿瘤发生、增殖凋亡及侵袭转移等[4],既往研究发现MMP-9是ERK信号通路下游分子[15-16]。为了进一步探究分子机制,本研究通过Western blot检测ERK、p-ERK和MMP-9蛋白水平的变化,结果发现SM22α过表达抑制p-ERK和MMP-9的蛋白水平。为了进一步验证SM22α通过调节ERK/MMP-9信号通路发挥功能,应用ERK抑制剂PD98059阻断ERK信号通路,检测SM22α过表达细胞MMP-9的表达水平,发现阻断ERK信号通路后,SM22α过表达不能引起MMP-9表达下调,说明SM22α可能通过ERK/MMP-9信号通路调节结直肠癌侵袭和迁移能力。
综上所述,SM22α抑制结直肠癌细胞侵袭和迁移能力,抑制ERK/MMP-9信号通路是其可能的分子机制。进一步深入探究SM22α抑制结直肠癌侵袭迁移的分子机制,将可能发现转移性结直肠癌治疗的新靶点,为提高结直肠癌的总体治疗水平和预后起到重要作用。
