黄曲霉产β-1,4-木聚糖酶 发酵条件的优化及其酶学特性
2019-04-12杨柳
, , ,杨柳,,
(北京工商大学食品学院,北京 100048)
木聚糖是半纤维素中主要的碳水化合物,也是植物细胞壁最主要的多糖成分,它的结构复杂多变,主链由多个木糖经β-1,4-糖苷键连接而成,同时还含有多种不同取代基形成的支链[1]。因此,木聚糖的降解需要多种酶的协同作用,其中最核心的就是β-1,4-木聚糖酶,用于随机性切断木聚糖的主链。目前,β-1,4-木聚糖酶在食品、饲料、造纸及生物转化等领域都有着广泛的应用。随着β-1,4-木聚糖酶研究的不断深入,人们对一些更具独特性能的β-1,4-木聚糖酶的需求也日益增多。
β-1,4-木聚糖酶广泛分布于自然界,而其主要来源是细菌、放线菌及真菌等[2-3]微生物。其中,丝状真菌凭借胞外酶分泌比例高、耐酸性能好等优势逐渐成为该酶工业化生产的首选菌种。曲霉一直是酶制剂工业化生产的主要菌种,也是β-1,4-木聚糖酶的重要生产菌种之一。至今,包括黑曲霉、烟曲霉、土曲霉、米曲霉和硫色曲霉等[1-2,4-5]多种曲霉均被发现具有产β-1,4-木聚糖酶的能力,但有关黄曲霉产β-1,4-木聚糖酶的报道还很少。黄曲霉作为酶制剂生产的重要菌种之一,具有独特产酶特性,它能高效分泌纤维素酶、内切葡聚糖酶、α-淀粉酶和β-葡萄糖苷酶等[6-8]。目前,关于黄曲霉β-1,4-木聚糖酶的报道主要集中于产酶能力的优化及酶的分离纯化[9-12]。在黄曲霉发酵产酶工艺研究方面,更多的是采用固态发酵的方式来优化菌株的产酶水平,而有关液态深层次发酵产酶的报道较少。国内,仅发现李科等[13]曾报道过黄曲霉液态发酵产β-1,4-木聚糖酶的研究;国际上,Bharat等[14]及Pal等[15]都曾研究过黄曲霉液态发酵生产β-1,4-木聚糖酶,但其产酶水平均低于40.0 U/mL,产量偏低。可见,目前国内外关于黄曲霉液态发酵产β-1,4-木聚糖酶的能力提升等相关研究仍存在诸多不足,非常值得深入探讨,这对于挖掘β-1,4-木聚糖酶的新资源具有重要意义。
本研究即以黄曲霉为出发菌株,旨在研究其液态发酵产β-1,4-木聚糖酶的能力以及该酶的酶学特性,以期为黄曲霉发酵生产β-1,4-木聚糖酶及其应用提供理论依据。
1 材料与方法
1.1 材料与仪器
榉木木聚糖(纯度为95%) 英国Apollo Scientific公司;大麦葡聚糖、脱枝阿拉伯聚糖 爱尔兰Megzyme公司;羧甲基微晶纤维素钠(CMC) 中国Macklin公司;对硝基苯-β-D-吡喃木糖苷、邻硝基苯-β-D-吡喃半乳糖苷 美国Sigma公司;酵母提取物、胰蛋白胨 英国Oxoid公司;燕麦粉、大麦粉 北京永辉超市;玉米芯、小麦麸皮、玉米苞皮、稻壳、白酒酒糟、花生壳等 北京官园花鸟市场;吐温、曲拉通、SDS和其余试剂 中国国药集团化学试剂有限公司;黄曲霉(CICC. 2476,不产黄曲霉毒素) 中国工业微生物菌种保藏中心。
MJPS-150型霉菌培养箱 上海精宏有限公司;ZWF-100型摇床 上海智诚分析有限公司;HH.S11-4型电热恒温水浴锅 上海博讯医疗有限公司;TGL-20BR型高速冷冻台式离心机 上海安亭科学仪器厂;TU-1810PC型紫外可见分光光度计 北京普析通用仪器设备有限责任公司;SDS-PAGE电泳仪 美国BIO-RAD公司。
1.2 实验方法
1.2.1 发酵产β-1,4-木聚糖酶 将黄曲霉划线接种于PDA平板上,置于30 ℃下培养3~5 d,再将大小为1 cm2黄曲霉菌块接种到含50 mL初始发酵培养基的250 mL锥形瓶中,37 ℃、200 r/min条件下培养4 d,然后在4 ℃条件下离心10 min(转速为10000 r/min)取上清,测定上清液的β-1,4-木聚糖酶酶活力。
初始发酵培养基为:2%玉米芯,1%胰蛋白胨,1%酵母提取物,0.5% NaCl,0.05% MgSO4·7H2O和0.075% KH2PO4,自然pH。
1.2.2 发酵产β-1,4-木聚糖酶条件的优化 以β-1,4-木聚糖酶的酶活力为考察指标,采用单因素实验优化黄曲霉的产酶条件,其他固定因素水平均与1.2.1相同。首先考察碳源对黄曲霉产β-1,4-木聚糖酶的影响,包括碳源种类(玉米芯、麸皮、燕麦粉、大麦粉、稻草、玉米苞皮、白酒酒糟、花生壳、葡萄糖、木糖、蔗糖、乳糖、麦芽糖、可溶性淀粉和葡聚糖,添加量均为2.0%)和麸皮添加量(0、0.5%、1.0%、1.5%、2.0%、2.5%、3.0%、3.5%、4.0%、4.5%和5.0%);接着考察氮源种类(胰蛋白胨、酵母提取物、蛋白胨、牛肉膏、大豆蛋白胨、豆粕粉、硫酸铵和磷酸氢二铵,添加量均为1.0%)及磷酸氢二铵添加量(0、0.5%、1.0%、1.5%、2.0%、2.5%、3.0%、3.5%、4.0%、4.5%和5.0%)对产β-1,4-木聚糖酶的影响;之后考察表面活性剂种类(吐温-20、吐温-40、吐温-60、吐温-80、曲拉通-110、曲拉通-114和SDS,添加量均为0.5%)及吐温-60添加量(0、0.5%、1.0%、1.5%、2.0%、2.5%和3.0%)对产β-1,4-木聚糖酶的影响。在此基础上,进一步优化发酵温度(25~45 ℃,每隔5 ℃)和发酵时间(0、1、2、3、4、5、6和7 d)对产β-1,4-木聚糖酶水平的影响。
1.2.3β-1,4-木聚糖酶酶活力及蛋白含量的测定β-1,4-木聚糖酶活力测定参照Liu等[16]的方法:180 μL榉木木聚糖(浓度为1.0%)底物在45 ℃下预热3 min,加入20 μL适当稀释的β-1,4-木聚糖酶液并于45 ℃下反应10 min,再加入200 μL的3,5-二硝基水杨酸(DNS)溶液后煮沸15 min显色,最后加入200 μL饱和酒石酸钾钠溶液(浓度为40%),待冷却后测定A540 nm值,以木糖为标准。β-1,4-木聚糖酶酶活力单位(U)定义为:在上述条件下每分钟水解榉木木聚糖生成1 μmol木糖所需的β-1,4-木聚糖酶量。
1.2.4 黄曲霉胞外β-1,4-木聚糖酶的盐析分级 收集黄曲霉分泌的胞外β-1,4-木聚糖酶发酵液,在4 ℃条件下以10000 r/min的转速离心10 min,收集发酵上清液。使用研钵将硫酸铵研碎成粉末,放置于105 ℃烘箱中烘干4 h以上待用。将装有发酵上清液的烧杯放置于冰水浴中并保持低速旋转(250 r/min),持续、缓慢地向其中添加预冷的硫酸铵粉末(确保上清液无泡沫、烧杯底部无沉积出现)至硫酸铵的饱和度为20%,然后在4 ℃条件下,10000 r/min的转速离心20 min,分离上清液和沉淀,收集沉淀并保存于4 ℃待用。分离后的上清液继续重复上述盐析分级步骤并分别收集饱和度达到0~20%、20%~40%、40%~60%和60%~80%时所沉淀出的蛋白。最后用磷酸盐缓冲液(20 mmol/L,pH8.0)重悬保存的沉淀并转移至透析袋中透析过夜(透析液与重悬液相同),在4 ℃条件下以10000 r/min的转速离心10 min,收集上清液待用。
1.2.5 SDS-PAGE及β-1,4-木聚糖酶酶谱分析 SDS-PAGE参照Laemmli等[17]的方法:分离胶为12.5%,浓缩胶为4.5%,考马斯亮蓝R-250显色。
酶谱分析方法:SDS-PAGE后(电泳胶中含有0.2%的榉木木聚糖),将电泳胶取出用异丙醇(浓度为25%)溶液浸泡三次(10 min/次)复性蛋白,然后用缓冲液浸泡三次(10 min/次)洗去异丙醇溶液,最后将电泳胶置于37 ℃条件下保温20 min,加入刚果红(浓度为0.15%)溶液染色30 min,再用NaCl(浓度为1 mol/L)溶液脱色直至显现透明带,最终以乙酸(浓度为0.5%)溶液浸泡1 min显色,出现的透明条带即是β-1,4-木聚糖酶。
1.2.6 黄曲霉β-1,4-木聚糖酶的酶学特性研究
1.2.6.1 最适pH测定 在50 ℃条件下,分别测定不同pH及不同缓冲体系下(MES,pH5.0~6.5;MOPS,pH6.5~8.0;Tris-HCl,pH7.5~9.0;Glycine-NaOH,pH9.0~10.5)β-1,4-木聚糖酶的酶活力,以酶活力最高点作为100%。
1.2.6.2 pH稳定性的测定 用1.2.6.1中不同pH的缓冲液分别稀释酶液(蛋白终浓度为0.5 mg/mL),将稀释好的酶置于45 ℃水浴锅中分别处理30 min,再迅速将样品放置于冰水中冷却30 min,测定残余酶活力。以未经处理的β-1,4-木聚糖酶为空白对照,计算残余酶活力占对照酶活力的百分比。
1.2.6.3 最适温度测定 用MOPS缓冲溶液(50 mmol/L,pH7.5)配制底物,分别在不同温度(30~70 ℃)下按照1.2.3方法测定β-1,4-木聚糖酶的酶活力,以酶活力最高点为100%。
1.2.6.4 温度稳定性的测定 用MOPS缓冲溶液(50 mmol/L,pH7.5)适当稀释酶液(蛋白终浓度为0.5 mg/mL)后放置于不同温度(30~65 ℃,每隔5 ℃)下处理30 min,再迅速将样品置于冰水中冷却30 min,测定残余酶活力。以未经处理的β-1,4-木聚糖酶为空白对照,计算残余酶活力占对照酶活力的百分比。
1.2.6.5 底物特异性测定 分别以浓度为1%的榉木木聚糖和羧甲基微晶纤维素钠,浓度为0.5%的脱枝阿拉伯聚糖和大麦葡聚糖以及浓度为5 mmol/L对硝基苯-β-D-吡喃木糖苷和邻硝基苯-β-D-吡喃半乳糖苷为水解底物,按照1.2.3方法测定酶活力,以榉木木聚糖为底物时的酶活力为100%,分别计算各底物对应相对酶活。
1.2.6.6 水解榉木木聚糖的特异性研究 向榉木木聚糖(浓度为1%)中加入10 U/mL的β-1,4-木聚糖酶进行水解反应,分别在不同时间点取样并经沸水蒸煮5 min以灭酶活,10000 r/min离心10 min后取上清待测。处理好的样品再经TLC分析,条件如下:采用德国Merck公司60F 254型号硅胶板,展层剂为正丁醇/乙酸/水=2∶1∶1 (v/v/v),显色剂为5%硫酸甲醇溶液,展层2次。
1.3 数据处理
本研究中涉及的实验内容均重复3次;黄曲霉发酵条件优化的数据处理均采用Microsoft Office Excel软件(2007版);图片的编辑和处理均采用Microsoft Office Picture Manager软件(2007版)。
2 结果与分析
2.1 碳源对黄曲霉发酵产胞外β-1,4-木聚糖酶的影响
碳源(2.0%)对黄曲霉发酵产胞外β-1,4-木聚糖酶的能力有重要影响(表1)。结果表明,以麸皮为单一碳源时黄曲霉胞外产β-1,4-木聚糖酶能力最强,当麸皮添加量为3.5%时得到最高酶活力,达31.83 U/mL(图1)。
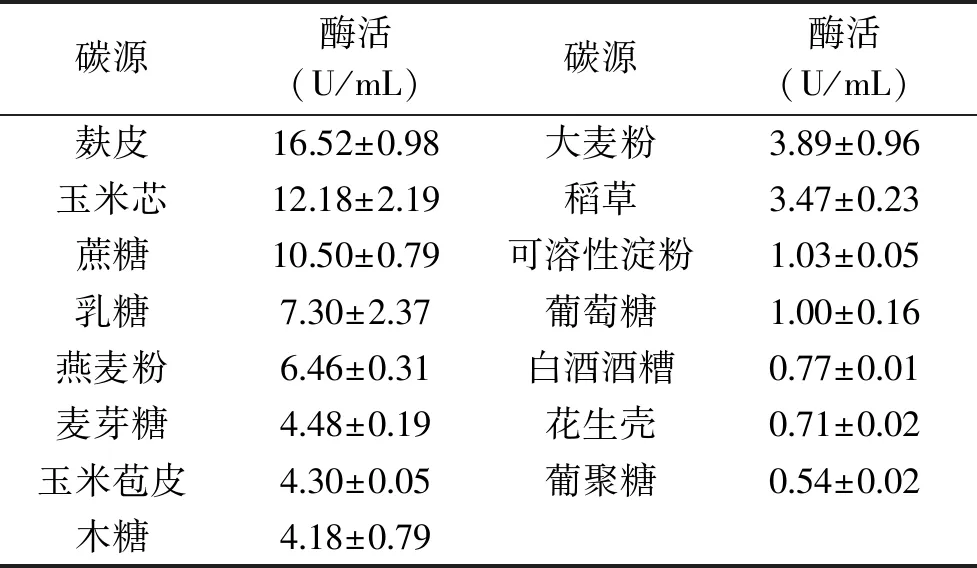
表1 碳源对黄曲霉发酵产β-1,4-木聚糖酶能力的影响Table 1 Effects of carbon sources on β-1,4-xylanase production by A. flavus
黄曲霉能利用天然纤维质原料发酵生产β-1,4-木聚糖酶,其产酶量明显高于单一糖类,说明黄曲霉更适合利用此类营养全面的天然底物。这可能是因为麸皮不仅富含蛋白质、糖类、维生素和矿物质等常见营养元素,还含有一些能促进微生物生长和产β-1,4-木聚糖酶的生长因子,它既可以作为碳源,又可以作为氮源。此外,麸皮还是农业生产过程当中的废弃物,价格低廉,它的利用可以一定程度上节约酶的生产成本。多种曲霉菌株也被发现具有代谢利用纤维质原料并诱导分泌β-1,4-木聚糖酶的能力,但它们的生产方式主要集中于固态发酵[9]。由图1可知,麸皮的添加量会对菌株的产酶能力造成影响,过低的添加浓度会造成培养基的养分不足,难以满足黄曲霉发酵产酶的需求;但过量添加麸皮又很容易引起培养基黏度的增加,导致溶氧降低,反而抑制了黄曲霉产β-1,4-木聚糖酶的能力。

图1 麸皮的添加量对黄曲霉产胞外 β-1,4-木聚糖酶的影响Fig.1 Effect of the additive amout of wheat bran on β-1,4-xylanase production
2.2 氮源对黄曲霉发酵产胞外β-1,4-木聚糖酶的影响
氮源(1.0%)能显著影响黄曲霉产β-1,4-木聚糖酶的能力(表2)。磷酸氢二铵是黄曲霉最适合的氮源(酶活力为58.66 U/mL)。研究还发现,当磷酸氢二铵添加量为3.0%时β-1,4-木聚糖酶的酶活力最高,达64.43 U/mL(图2)。

表2 氮源对黄曲霉发酵产β-1,4-木聚糖酶的能力影响Table 2 Effects of nitrogen sources on β-1,4-xylanase production by A. flavus
由表1可知,黄曲霉以磷酸氢二铵为氮源时的产酶能力明显高于有机氮源。而且,无机氮源的成本通常要低于有机氮源,这将有效降低该酶的生产成本。由图2可知,磷酸氢二铵的添加量也会严重影响黄曲霉的产酶能力,添加量过高,会使菌体生长过于旺盛,不利于代谢产物的积累,而添加量不足,菌体繁殖速率降低,影响了黄曲霉分泌β-1,4-木聚糖酶。
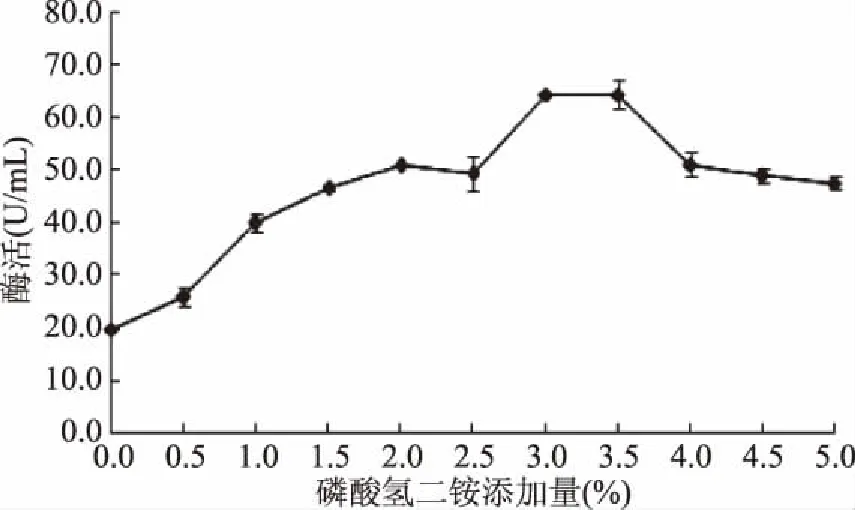
图2 磷酸氢二铵的添加量对黄曲霉产胞外 β-1,4-木聚糖酶的影响Fig.2 Effect of the additive amout of(NH4)2HPO4 on β-1,4-xylanase production
2.3 表面活性剂对黄曲霉发酵产胞外β-1,4-木聚糖酶的影响
表面活性剂对黄曲霉产胞外β-1,4-木聚糖酶的影响结果如表3所示。非离子型表面活性剂促进黄曲霉产β-1,4-木聚糖酶的作用明显高于离子型表面活性剂,以吐温-60作用效果最好。由图3知,添加1.0%的吐温-60时黄曲霉的产酶水平最高,酶活为86.42 U/mL。

图3 吐温-60添加量对黄曲霉产胞外 β-1,4-木聚糖酶的影响Fig.3 Effect of the additive amout of Tween-60 on β-1,4-xylanase production
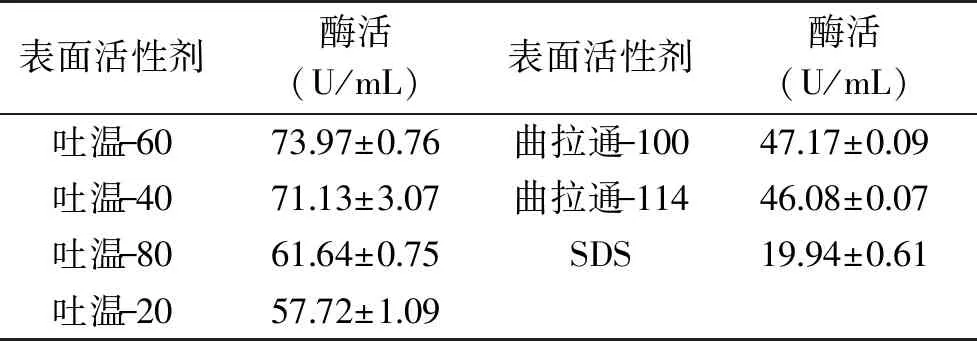
表3 表面活性剂对黄曲霉发酵产 β-1,4-木聚糖酶的能力影响Table 3 Effects of surfactants on β-1,4-xylanase production by A. flavus
一定浓度的表面活性剂通常具有增强微生物细胞膜通透性的功能,促进胞内酶向外渗透,进而提高胞外酶产量[18]。本研究发现吐温-60是最优的表面活性剂,在其作用下黄曲霉胞外β-1,4-木聚糖酶产量增长了47.0%,具有较强的促进效果。但过量添加吐温-60又很容易导致产酶量下降,可能是高浓度的表面活性剂造成了细胞膜的裂解,引起细胞内的电解质大量泄漏,从而导致代谢失调产生抑制作用。
2.4 发酵温度对黄曲霉发酵产胞外β-1,4-木聚糖酶的影响
发酵温度对黄曲霉发酵产β-1,4-木聚糖酶具有重要影响(图4)。结果表明,黄曲霉在35 ℃条件下培养时能够高产β-1,4-木聚糖酶,最高酶活为89.31 U/mL。
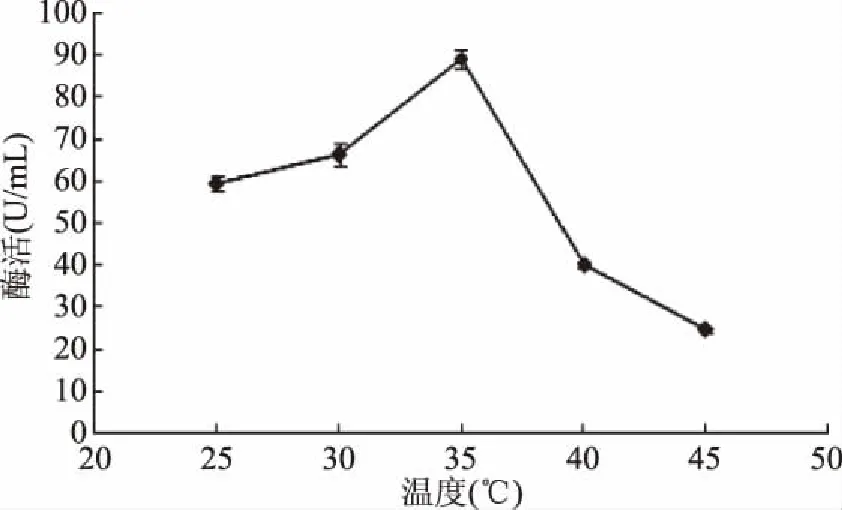
图4 发酵温度对黄曲霉发酵产胞外 β-1,4-木聚糖酶的能力影响Fig.4 Effect of fermentation temperature on β-1,4-xylanase production
温度是限制微生物生长代谢的关键因子之一,温度升高时菌体生长加快,且促进了合成酶等活性物质的作用效率。黄曲霉在35 ℃下产酶能力最强,但进一步升高温度时黄曲霉的生长明显受到抑制,导致其酶活降低。
2.5 发酵时间对黄曲霉发酵产胞外β-1,4-木聚糖酶的影响
基于以上优化的结果,最终研究发酵时间对黄曲霉产β-1,4-木聚糖酶的影响(图5)。β-1,4-木聚糖酶的酶活力随着时间的延长而逐渐增高,发酵至第5 d时酶活力最高,达115.08 U/mL。

图5 发酵时间对黄曲霉发酵产胞外 β-1,4-木聚糖酶的能力影响Fig.5 Effect of fermentation time on β-1,4-xylanase production
国内外主要采用固态发酵的方式优化黄曲霉产β-1,4-木聚糖酶的能力[9],而有关液态深层次培养发酵产酶的报道仍然较少。Bhushan等[11]的研究发现,黄曲霉能以小麦秸秆和玉米粉为原料发酵产β-1,4-木聚糖酶,发酵6 d后酶活力达27.30 U/mL;Pal等[15]研究发现,黄曲霉DFR-6同样能够以麸皮为碳源并发酵生产β-1,4-木聚糖酶,发酵至第6 d达到最高酶活31.40 U/mL;而国内仅发现李科等[13]系统研究过黄曲霉发酵产β-1,4-木聚糖酶的能力,发现该菌能以玉米芯为碳源发酵产酶,至第6 d时达到最高酶活(122.23 U/mL)。与上述报道相比,本研究中的黄曲霉具有相对较强的发酵生产β-1,4-木聚糖酶的能力,在发酵至第5 d时即可达到最高酶活(115.08 U/mL),表现出更高效的产酶能力,是未优化时的9.4倍。
2.6 黄曲霉胞外分泌β-1,4-木聚糖酶的特性分析
为解析黄曲霉分泌β-1,4-木聚糖酶的基本特性,首先采用盐析法对黄曲霉分泌的β-1,4-木聚糖酶进行分级沉淀,实验结果经SDS-PAGE和β-1,4-木聚糖酶酶谱分析(图6)。结果表明,黄曲霉能够分泌两种β-1,4-木聚糖酶,它们分别在硫酸铵饱和度达40%~60%和60%~80%时沉淀析出,其分子量大小分别为20.1和31.0 kDa左右(分别命名为β-1,4-木聚糖酶A和β-1,4-木聚糖酶B)。目前,国际上虽已有部分关于黄曲霉分泌β-1,4-木聚糖酶的报道,但其研究的β-1,4-木聚糖酶分子量仅有14.0、28.5和35.0 kDa三种,均与本研究中的β-1,4-木聚糖酶分子量不同[10-12]。因此,有必要对该酶的酶学特性进行深入研究。

图6 不同饱和度硫酸铵分级沉淀β-1,4-木聚糖酶的 SDS-PAGE及β-1,4-木聚糖酶酶谱分析Fig.6 SDS-PAGE and zymogram analysis of β-1,4-xylanase production by A. flavus
2.7 黄曲霉β-1,4-木聚糖酶的酶学特性研究
为了分析黄曲霉β-1,4-木聚糖酶粗酶的酶学特性,本研究分别考察了该酶的最适pH和pH稳定性(图7a、7b)、最适温度和温度稳定性(图7c、7d)以及底物特异性(表4)。结果显示,该酶的最适pH和最适反应温度分别为MOPS 7.5和55 ℃;该酶可在pH5.5~9.0范围内及30~50 ℃保持酶活力的稳定,相对酶活力均在80%以上;黄曲霉β-1,4-木聚糖酶不仅能水解榉木木聚糖,还能微弱降解大麦葡聚糖,表现出一定的β-1,3-1,4-葡聚糖酶的活性。进一步研究发现,该酶能够快速水解榉木木聚糖(图8),并最终生成木二糖、木五糖和木六糖等一系列木寡糖。

表4 β-1,4-木聚糖酶的底物特异性测定结果Table 4 Substrate specifitiy of the β-1,4-xylanase from A. flavus
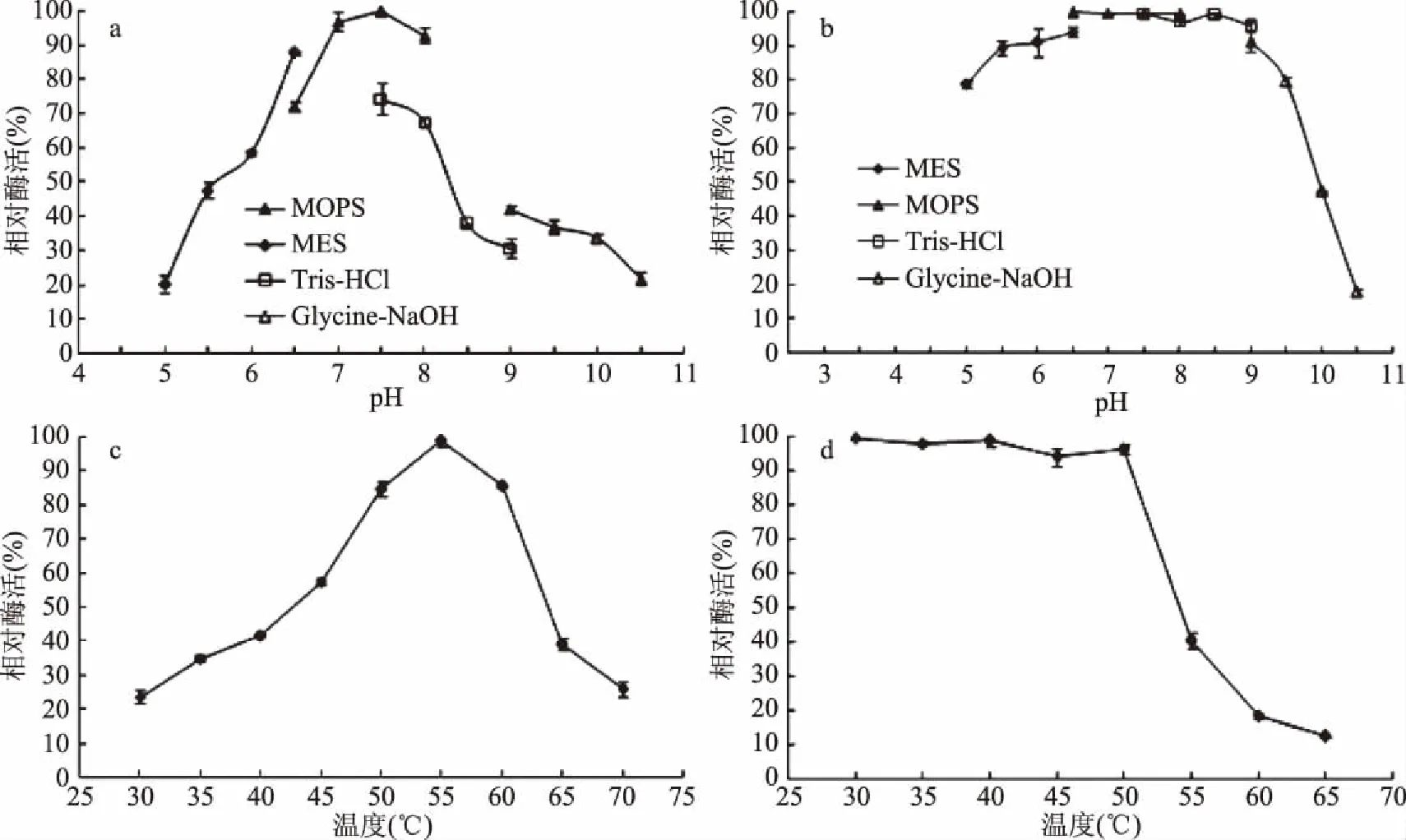
图7 黄曲霉发酵产胞外β-1,4-木聚糖酶的温度及pH特性Fig.7 Characterization of the temperature and pH of β-1,4-xylanase
目前,β-1,4-木聚糖酶被广泛应用在不同的工业领域,如造纸工业中,它主要用于漂白纸浆。然而,纸浆制造生产通常需要在偏碱性的环境下进行,这对于那些在碱性环境中酶活力低、稳定性差的β-1,4-木聚糖酶而言非常不利[19]。本研究的β-1,4-木聚糖酶最适pH为7.5,且在pH5.5~9.0范围内具有优越的稳定性能,表明它更加适用于这种偏碱性的环境。相比而言,多数曲霉来源的β-1,4-木聚糖酶仅在弱酸性(pH在5.0~7.0范围内)条件下表现有较高的酶活力[9,11,20-21],这也严重限制了它们的应用范围。此外,众多曲霉来源的β-1,4-木聚糖酶也只能在较窄的pH范围内保持酶活力的稳定,例如其他黄曲霉来源的β-1,4-木聚糖酶仅在pH范围为4.5~6.5时保持酶活力稳定[11],还有黑曲霉和烟曲霉来源的β-1,4-木聚糖酶分别在pH范围为4.0~6.5[15]和7.0~9.0[21]时保持酶活力的稳定。据报道,曲霉菌所分泌的β-1,4-木聚糖酶通常在45~60 ℃内表现出最高酶活力,本研究的β-1,4-木聚糖酶最适温度为55 ℃,说明该酶属于典型的曲霉β-1,4-木聚糖酶。此外,该酶与其它丝状真菌来源的β-1,4-木聚糖酶类似,都能保证在50 ℃及以下温度保持稳定的酶活力[20,22]。
众多β-1,4-木聚糖酶不仅对木聚糖底物具有很强的水解作用,还能够降解CMC、地衣多糖、淀粉和果胶等聚糖类底物以及对硝基苯-β-D-吡喃木糖苷等糖苷类底物[23]。而本研究中的β-1,4-木聚糖酶在对木聚糖底物具有很强水解作用的同时,还能微弱水解大麦葡聚糖(表4),这对于强化该酶降解木质纤维素材料的应用具有重要意义。另一方面,该酶不具备降解CMC的能力,有效避免了纸浆中的纤维素被降解,将有利于它在纸浆漂白工业中的应用,这一特性也与来源于聚多曲霉和烟曲霉的β-1,4-木聚糖酶类似[24-25]。此外,该酶也不能水解对硝基苯-β-D-吡喃木糖苷底物,说明它没有β-木糖苷酶的活性,这也为其制备和生产功能性低聚木糖奠定了良好的理论基础。
如图8所示,黄曲霉分泌的β-1,4-木聚糖酶可在0.5 h内快速将榉木木聚糖降解并生成不同聚合度的低聚木糖;随着时间的延长,降解产物又进一步被水解,最终生成木二糖、木五糖及少量的木六糖。一般来说,曲霉β-1,4-木聚糖酶降解木聚糖底物主要生成木二糖和木三糖[26],可见本研究中的β-1,4-木聚糖酶的水解特异性与常见木聚糖酶的特异性并不相同。据了解,木二糖至木七糖是目前公认的功能性低聚糖,其中木二糖具有最好的益生活性、木五糖还具有良好的抗肿瘤功效[27-28]。

图8 黄曲霉胞外β-1,4-木聚糖酶 水解木聚糖的TLC分析图Fig.8 TLC analysis of beechwood xylan hydrolytic by β-1,4-xylanase from A. flavus
3 结论
本研究发现黄曲霉能够利用廉价的麸皮发酵生产β-1,4-木聚糖酶,经培养基组成及培养条件优化后该菌株最高可产115.08 U/mL的β-1,4-木聚糖酶,高于其他报道;该菌能够分泌两种β-1,4-木聚糖酶,且分子量大小均不同于已有报道。酶学性质结果表明,该酶的最适pH和最适温度分别为MOPS 7.5和55 ℃,并在pH5.5~9.0范围内以及30~50 ℃保持酶活力的稳定;该酶不仅可以降解榉木木聚糖,还能够微弱水解大麦葡聚糖,有助于其在木质纤维素材料降解当中的应用。研究还发现,黄曲霉β-1,4-木聚糖酶可以降解榉木木聚糖,并主要生成木二糖和木五糖等功能性低聚糖,具有潜在的应用前景。
